Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Paolo Macchiarini wikipedia , lookup
Sexual reproduction wikipedia , lookup
Cell culture wikipedia , lookup
Cell encapsulation wikipedia , lookup
Somatic cell nuclear transfer wikipedia , lookup
Development of the nervous system wikipedia , lookup
Regeneration in humans wikipedia , lookup
Cellular differentiation wikipedia , lookup
4/7/2015
Morphogenesis and Differentiation in Animal Development | Principles of Biology from Nature Education
Principles of Biology
164 Morphogenesis and Differentiation in Animal Development
contents
Every embryo undergoes a series of stages wherein cells divide, migrate,
and differentiate into the organism's adult form. Each cell performs a specific
role in the body, serving as a component of a particular organ system. An
immature cell can differentiate into part of the skeletal system or digestive
system, depending on the signals that direct its maturation process. How do
cells know what to do during development? Why do some cells move to a
certain area of the developing body and not to another area? The terms
morphogenesis and differentiation encompass these developmental
processes, including directing cells to move towards specific areas and
inducing these cells to mature into fully functional, specialized cells that the
body needs to survive.
Morphogenesis and Differentiation During Animal
Development
In very broad terms, two forces influence the migration and maturation of
cells during morphogenesis. One involves cytoplasmic factors found in the
egg that direct cells to differentiate into specific cell types. These cytoplasmic
proteins and messenger RNA molecules, which are derived solely from the
maternal genome, induce an immature cell to follow a particular
differentiation route. The other force originates from the external environment
of the cell, manipulating the differentiation process towards a specific
outcome based on the conditions of its surroundings. This is the result of
cellcell interactions and the release of chemical signals from cells that are
detected by neighboring cells, which in both cases trigger signaling pathways
within the recipient cells. These signaling pathways cause changes in gene
expression and protein activity that ultimately result in morphogenesis and
differentiation.
The process of morphogenesis requires the establishment of specific body
axes separating regions of the body, an organization related to the symmetry
of developing and fully formed organisms. This form of axial orientation
guides cells during migration and differentiation, assisting in the correct
physical development of body parts. An animal's body can be separated into
three major axes. The anteriorposterior axis separates the body into the
head region and the tail region. The mediallateral axis divides the body into
central and left/right regions, while the dorsoventral axis defines the dorsal,
or back, region and the ventral, or chest, region. Each of these axes triggers
the production of specific proteins needed for the proper development of
arms, feet, or the spine. For example, expression of the dorsal gene in the fruit
fly Drosophila generates a protein called Dorsal, which is a transcription factor
that guides gene expression in immature cells of the embryo (Figure 1). Note
that in Figure 1, Dorsal is expressed in a gradient along the dorsalventral
axis. Expression is highest on the ventral side of the embryo. The differing
concentrations of Dorsal impact the expression of specific sets of genes in
those groups of cells. These different patterns of gene expression help to
define the dorsalventral axis. Mutation of the dorsal gene results in only
dorsal tissues being formed; hence the name of the gene.
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145882/1
1/8
4/7/2015
Morphogenesis and Differentiation in Animal Development | Principles of Biology from Nature Education
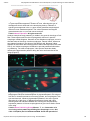
Figure 1: Morphogenesis in fruit
files.
The red fluorescence in this image of a
fruit fly embryo (Drosophila) shows the
presence of the morphogen Dorsal in
the immature cells. Dorsal is a
transcription factor that helps define the
dorsalventral orientation of the
embryo. The concentration of Dorsal in
each cell will determine which type of
cell it will mature into. scale bar = 100
µm
© 2011 Nature Publishing Group
Chung, K. et al. A microfluidic array
for largescale ordering and
orientation of embryos. Nature Methods
8, 171–176 (2011).
doi:10.1038/nmeth.1548. Used with
permission.
Cytoplasmic factors found in the egg direct cells to differentiate into
specific cell types.
As the cells in the embryo divide, maternal components of the egg's
cytoplasm play a major role in morphogenesis, directing the migration of
immature cells and their differentiation into specific cell types. Many of these
signaling molecules, which are sometimes called morphogens because of
their role in directing morphogenesis, form concentration gradients within an
embryo. The varying concentration of a morphogen leads to differential gene
expression in neighboring groups of cells. Thus, the morphogen gradient
functions as positional information that activates developmental processes in
cells based on their relative locations. Some morphogens are also secreted
from cells, where they serve as chemical signals among neighboring cells.
The morphogens initiate a series of cellular responses, such as the
induction of neighboring cells to produce a second wave of messengers.
These cellular responses again alter gene expression patterns in the
affected cells, creating the continuum of reactions necessary for
morphogenesis.
Test Yourself
If a concentration gradient of a morphogen was not established across an embryo, would
morphogenesis as directed by that morphogen still proceed in the same manner? Why or why
not?
Submit
Asymmetric cell division facilitates diversity among cells in a developing
embryo. The initial division of a zygote generates two identical daughter
cells. If maternal cytoplasmic factors are distributed evenly in the cell, then
the daughter cells share them equally and at that point share the same fate
or potential. But if the morphogens are located primarily in one of the cell
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145882/1
2/8
4/7/2015
Morphogenesis and Differentiation in Animal Development | Principles of Biology from Nature Education
poles, then the distribution of these morphogens in the daughter cells will be
unequal. This asymmetry sets the stage for two cell lineages in which the
progeny of these two daughter cells will be induced to differentiate along
separate pathways. Morphogens trigger specific signaling pathways that
influence cellular and biochemical reactions, resulting in the maturation of
cells and creating fully differentiated cells. This directional signaling can be
observed in embryos of the nematode Caenorhabditis elegans, in which a
fluorescent morphogen has been engineered. During interphase, the green
fluorescence covers the entire embryo but moves towards one pole at
anaphase. After the completion of the initial cell division, the green dye
moves exclusively to one daughter cell, establishing polarity across the entire
embryo (Figure 2).
Figure 2: Asymmetric cell division in the nematode Caenorhabditis
elegans.
In embryos of the nematode Caenorhabditis elegans, a morphogen, indicated
by the green fluorescence, appears across the entire embryo, eventually
moving toward one pole during anaphase. After one cell division cycle, the
morphogen has moved to one daughter cell, creating polarity across the
embryo.
© 2008 Nature Publishing Group Gönczy, P. Mechanisms of
asymmetric cell division: flies and worms pave the way. Nature Reviews
Molecular Cell Biology 9, 355366 (2008) doi:10.1038/nrm2388. Used with
permission.
Cellcell interactions and cell signaling are essential for proper
patterning of cells, tissues, and organs.
How do morphogens establish and maintain a concentration gradient across
a developing embryo? Morphogens generally last for a specific duration
within the extracellular matrix and are eventually removed from circulation.
Scientists have tried to induce a gradient in embryos by introducing a dye
that does not interact with morphogens. While the dye quickly spreads
around the embryos by diffusion, morphogens and other cytoplasmic factors
maintain their positions within the developing gradient, in opposition to the
embryo's forces of active transport and diffusion.
Researchers have proposed three models of morphogen gradient formation
(Figure 3). The models also describe how these signaling molecules retain
their positions in respective sites within the developing embryo. One model
suggests that morphogens move randomly from the extracellular space into
cells by Brownian motion. A second model implicates directed cycles of
release and uptake of morphogens for transporting and maintaining high
concentrations within and around specific cells. A third model suggests that
low amounts of cellsurface carbohydrates facilitate uptake of morphogens
into cells, while cells with greater amounts of extracellular carbohydrates
prevent morphogens from entering cells.
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145882/1
3/8
4/7/2015
Morphogenesis and Differentiation in Animal Development | Principles of Biology from Nature Education
Figure 3: Proposed models for establishment of morphogen
gradients.
Three transport theories describe how morphogens enter specific cells,
creating gradients that influence embryonic development: by Brownian
motion, through directed cycles of release and uptake, or by the quantity
of cellsurface carbohydrate molecules.
© 2011 Nature Education All rights reserved.
Certainly, morphogens induce cells to differentiate into specific cell types.
But do morphogens trigger the production of specific proteins during
morphogenesis? Research studies have shown that over the course of
morphogenesis, cells secrete such morphogens (Figure 4). For example, in
the development of the dorsal neural tube in chicks, the morphogen sonic
hedgehog homolog (Shh) influences the production of transcription factors
needed for differentiation. Sonic hedgehog also determines which cells
undergo cellular activities by activating or suppressing the production of
these transcription factors. At approximately 6 hours into morphogenesis,
Shh activates production of the transcription factor Olig2 in all cells of the
neural tube. Six hours later, Shh facilitates the production of another
transcription factor, Nkx2.2, at the periphery of the dorsal neural tube, while
the inner regions of the neural tube retain Olig2. At 18 hours, both
transcription factors occupy the inner region of the neural tube. By the 24th
hour, Shh induces the entire structure to produce Nkx2.2. Throughout this
process, Shh also suppresses the production of the Pax7 transcription factor.
This regulation of protein production by Shh is an example of how
morphogens function during morphogenesis.
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145882/1
4/8
4/7/2015
Morphogenesis and Differentiation in Animal Development | Principles of Biology from Nature Education
Figure 4: Different morphogens
produced during development.
During morphogenesis of the neural
tube in chicks, the morphogen sonic
hedgehog homolog (Shh) influences
differentiation by inducing the
production of specific transcription
factors. Olig2 is stained red and Nkx2.2
is stained green (Pax7 would be
stained blue, but is absent from these
photos). During the first 6 hours of Shh
exposure, the protein Olig2 dominates
the entire structure, followed by Nkx2.2
at the 12th hour. At 18 hours, the
periphery of the neural tube contains
higher concentrations of Nkx2.2,
reaching its maximum concentration at
24 hours. Shh prevents the production
of the protein Pax7 during the entire
process.
© 2007 Nature Publishing Group
Dessaud, E., et al. Interpretation of
the sonic hedgehog morphogen
gradient by a temporal adaptation
mechanism. Nature 450, 717720
(2007) doi:10.1038/nature06347.
Used with permission.
Why is our head on the top of our body and our feet on the floor? Why do a
fly's wings appear where they do and not down by its legs? These are
questions of patterning along the anteriorposterior access of an animal. A
set of genes known as homeotic or Hox genes regulate the anteriorposterior
axis of an embryo. Highly conserved groups of Hox genes are found in many
animal phyla from fruit flies to humans (Figure 5). Each region of the Drosophila
embryo, for example, is dictated by specific Hox genes. One Hox gene dictates
the formation of the mandibular region, while another induces the formation
of the labial region. How did scientists study the role of these Hox genes? In
mutational studies, researchers removed specific gene segments in the Hox
gene cluster to identify morphological defects that would develop in the
resulting organism. Comparative analysis also showed that other vertebrate
species carry Hox genes, with speciesspecific modifications to their gene
cluster composition and order. DNA analysis allowed for the identification of
sequence motifs within Hox genes for binding with other proteins. A
counterpart of Hox genes also exists in plants, including some of the MADS
box and floral patterning genes.
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145882/1
5/8
4/7/2015
Morphogenesis and Differentiation in Animal Development | Principles of Biology from Nature Education
Figure 5: Hox genes and body patterning.
Hox gene expression appears in clusters or rows, reflecting the type of
development that is observed in the developing embryo. Deletion of
specific gene segments in the Hox gene cluster generates morphological
defects in most vertebrate species. The colors represent homologous
genes between the Drosophila and mouse embryos.
© 2011 Nature Education All rights reserved.
Another example of body patterning is visible in the spots in the wings of fruit
flies. These spots result from the morphogenetic effects of a specific
morphogen called Wingless. Secretion of the Wingless morphogen occurs in
discrete areas of the wings. Another morphogen, named Yellow, influences
spot pattern formation in the wings of fruit flies as well. Because Yellow
appears at sites where the brown pigment melanin later deposits in the adult
fruit fly, the Yellow morphogen is believed to provide positional information
on patterning. Two other morphogens, Vein spot and Intervein shade,
influence the pigmentation patterns along the veins and interveins of the
wings (Figure 6).
Figure 6: Morphogens and wing patterns in Drosophila melanogaster.
Morphogens influence the development of pigmented spots in the wings of
fruit flies. In these fluorescence microscopy photos, two morphogens, Vein
spot and Intervein, influence pigmentation patterns in the veins (a) and
interveins (b) of the wing. (c) Merged fluorescence image with veins
stained green and interveins stained red, showing the complementary
expression patterns of the two morphogens that give rise to distinct zones
in the wing.
© 2010 Nature Publishing Group Werner, T., et al. Generation of a
novel wing colour pattern by the Wingless morphogen. Nature 464,
1143–1148 (2010) doi:10.1038/nature08896. Used with permission.
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145882/1
6/8
4/7/2015
Morphogenesis and Differentiation in Animal Development | Principles of Biology from Nature Education
Cell adhesion molecules are critical for keeping cells connected.
The directed movement of tissues in a developing embryo results in the
generation of three major germ layers: the ectoderm, endoderm, and
mesoderm. These movements range from subtle cell migrations to massive
invaginations, changing the shape of the entire embryo. How do cells of each
germ layer remain intact during these movements? Is there a force or
mechanism that keeps them together as they slide over other tissues?
Critical in this process are a group of cell adhesion molecules called
cadherins. These are transmembrane glycoproteins that bind cells to one
another and to the extracellular matrix. Cytoskeletal structures, such as actin
filaments, within the cell provide support to the cytoplasmic region of the
cadherin proteins. The sugarcoated extracellular parts of these
glycoproteins serve as the recognition sites, often binding to the same
molecule on a different cell. This enables cells that express specific cadherin
proteins on their surface to stick to each other, while preventing different cell
types from binding. At the tissue level, cadherins allow each germ layer to
remain connected during embryogenesis.
Spemann's organizer controls the formation of embryonic axes.
Even during the early 1900s, biologists speculated about the driving force of
morphogenesis. In 1910, Alexander Gurwitsch presented the theory of the
morphogenetic field, which posited that cells developed into specific body
regions in response to specific chemicals secreted by the embryo. By the
1920s, scientists Hans Spemann and Hilde Mangold conducted a simple
transplantation experiment that confirmed this theory. Their experiment
involved embryos of two closely related newt species that followed different
patterns of pigmentation. These slight differences allowed a viable embryo to
be constructed that also demonstrated which newt was the source of
developmental results. The host embryo, Triton cristatus, was entirely non
pigmented while the donor embryo, Triton taeniatus, showed general
pigmentation. Transplantation of a specific portion on the dorsal side of the
host gastrula to its donor, thereafter called Spemann's organizer, resulted
in the development of a second notochord and neural tube in the host
embryo. Interestingly, these secondary structures were nonpigmented,
suggesting that these cells originated from the host embryo. Decades later,
scientists identified morphogens in these transplanted cells.
IN THIS MODULE
Morphogenesis and Differentiation During
Animal Development
Summary
Test Your Knowledge
WHY DOES THIS TOPIC MATTER?
Cancer: What's Old Is New Again
Is cancer ancient, or is it largely a
product of modern times? Can
cuttingedge research lead to prevention
and treatment strategies that could make
cancer obsolete?
Stem Cells
Stem cells are powerful tools in
biology and medicine. What can
scientists do with these cells and their
incredible potential?
PRIMARY LITERATURE
Classic paper: Fruit fly research
reveals how complex organisms
form (1980)
Mutations affecting segment number and
polarity in Drosophila.
View | Download
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145882/1
7/8
4/7/2015
Morphogenesis and Differentiation in Animal Development | Principles of Biology from Nature Education
SCIENCE ON THE WEB
Manipulating Embryos
Watch a video explaining Spemann's
experiment
page 843 of 989
2 pages left in this module
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145882/1
8/8
4/7/2015
Morphogenesis and Differentiation in Animal Development | Principles of Biology from Nature Education
Principles of Biology
164 Morphogenesis and Differentiation in Animal Development
contents
Test Your Knowledge
1. What is the series of cellular responses of neighboring cells that creates the
continuum of reactions necessary for morphogenesis?
induction
morphogen
cytoplasmic protein
body axes
None of the answers are correct.
2. What causes morphogens to generate a gradient?
The morphogens vary in concentration within an embryo.
asymmetrical cell division
induction
elimination
All answers are correct.
3. Which of the following suggests morphogens trigger the production of specific
proteins?
The morphogen sonic hedgehog influences the production of transcription factors.
The morphogen sonic hedgehog determines which cells undergo cellular activity.
The morphogen sonic hedgehog influences the activation or repression of multiple
transcription factors.
The morphogen sonic hedgehog stimulates the production of transcription factors,
such as OLIG2, in multiple cells to coordinate protein production in these cells.
All answers are correct.
4. Which of the following holds the three germ layers together as they slide over other
tissues?
morphogens
pheromones
ectoderm
cadherins
None of the answers are correct.
5. In the SpemannMangold experiment, what extra structure(s) developed after the
dorsal side of the donor's gastrula was transferred to the host organism?
newt embryos
notochord
morphogens
neural tube
a notochord and a neural tube
Submit
IN THIS MODULE
Morphogenesis and Differentiation During
Animal Development
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145882/3
1/2
4/7/2015
Morphogenesis and Differentiation in Animal Development | Principles of Biology from Nature Education
Summary
Test Your Knowledge
WHY DOES THIS TOPIC MATTER?
Cancer: What's Old Is New Again
Is cancer ancient, or is it largely a
product of modern times? Can
cuttingedge research lead to prevention
and treatment strategies that could make
cancer obsolete?
Stem Cells
Stem cells are powerful tools in
biology and medicine. What can
scientists do with these cells and their
incredible potential?
PRIMARY LITERATURE
Classic paper: Fruit fly research
reveals how complex organisms
form (1980)
Mutations affecting segment number and
polarity in Drosophila.
View | Download
SCIENCE ON THE WEB
Manipulating Embryos
Watch a video explaining Spemann's
experiment
page 845 of 989
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145882/3
2/2
4/7/2015
Embryonic Development | Principles of Biology from Nature Education
Principles of Biology
162 Embryonic Development
contents
Stages of Embryonic Development
In animals, embryonic development describes the earliest stages of
development. There are stages of embryonic development that are common
to all animals, although the specific details do differ. In general, embryos
develop through a series of sequential stages involving specific changes in
cells and tissues that eventually produce a newborn animal. In the initial
stage of development, called fertilization, the gametes (egg and sperm)
fuse and create a single cell or zygote. The zygote then undergoes multiple,
rapid cell divisions in a process known as cleavage. The resulting structure,
called a blastula, goes through complex rearrangements, resulting in
specialized cell layers. The resulting structure is known as a gastrula.
Organogenesis is the process by which these specialized cell layers
develop into specific organs.
Using classical embryological studies and more recent molecular
approaches, scientists have generated a comprehensive picture of how a
fertilized egg is transformed into a fully formed organism.
Different species exhibit diverse body plans, yet most species share a similar
course of embryonic development. For example, a human gene that
influences specific cells to develop into a heart has a counterpart in the fruit
fly. In flies, this gene, called the tinman gene, is responsible for the
development of the dorsal vessel, the insect equivalent of the heart. Because
of similarities such as this in the process of animal development, scientists
can use a variety of animals as model organisms to investigate how
development unfolds. Model organisms are often chosen because they are
easily manipulated in the laboratory or possess special features that make
them advantageous for studying development. For example, members of the
fruit fly genus Drosophila serve as classic model organisms due in part to
Drosophila's short life cycle and rapid development, which allow scientists to
investigate changes in specific body parts over several generations. In
humans, such studies might require decades of observations to generate
conclusions. In addition, the genetics of Drosophila are mapped in great detail,
and many mutant forms of Drosophila exist that represent specific
malformations, conditions, and diseases in other species.
When a sperm fertilizes an egg, a zygote forms.
Animal sperm are remarkable haploid gametes. They are designed to swim
to an egg, bind the egg, and through a set of chemical interactions with the
egg either enter the egg or release their nucleus into the egg so that two
haploid gametes can fuse together into a diploid zygote and begin the
process of embryonic development.
A sperm cell is constructed of three parts: the head, the midpiece, and the
tail (Figure 1). The head contains the nucleus and the acrosome. The
midpiece is the site of ATP production. Hydrolysis of ATP provides the
energy that drives the tail, which is a eukaryotic flagellum.
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145869/1
1/9
4/7/2015
Embryonic Development | Principles of Biology from Nature Education
Figure 1: Human sperm cells.
Sperm are motile haploid gametes. Each sperm cell is made of three
parts: a head, which contains the nucleus and the acrosome, the midpiece
filled with mitochondria, and the tail, which is a flagellum.
© 2011 Nature Publishing Group (left) Lishki, P. V., et al. Progesterone
activates the principal Ca2+ channel of human sperm. Nature 471, 387–
391 (2011) doi:10.1038/nature09767. Used with permission.
During fertilization, two gametes, an egg and a sperm, fuse, resulting in a
zygote (Figure 2). Each of the gametes is haploid and thus contributes half of
the genes that the embryo will possess. Fusion of the two gametes creates a
diploid zygote with a complete genome. This is the main purpose of
fertilization: to form a diploid cell that can then become an entire organism.
But the process of animal development also requires that the egg become
activated. This process is also initiated by the fusion of sperm and egg. As
we'll see, activation of the egg involves fairly rapid changes within the egg
that trigger the beginning stages of embryonic development.
When a sperm comes into contact with the surface of an egg (Figure 2),
chemical signals cause the release of enzymes from the head of the sperm
that dissolve part of the egg's protective external layer, exposing the egg's
plasma membrane. This enables the sperm and egg plasma membranes to
come into close proximity, and, in particular, it allows surface proteins on the
sperm to bind to specific receptors on the egg. The receptorprotein binding
stimulates a set of changes that cause the sperm and egg plasma
membranes to fuse. In addition, the surface of the egg undergoes changes
that prevent other sperm from reaching the plasma membrane. Let's now
look at these steps in more detail.
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145869/1
2/9
4/7/2015
Embryonic Development | Principles of Biology from Nature Education
Figure 2: Fertilization of an egg.
The surface of the egg plays an important role in the process of
fertilization. When a sperm comes in contact with the egg's surface, part of
the egg's protective external layer dissolves, exposing its plasma
membrane and allowing the fusion of the two gametes.
Don W. Fawcett/Science Source
Much of our knowledge about the changes in the surface of animal eggs at
fertilization comes from investigations of sea urchins, marine animals from
the phylum Echinodermata. How can such a simple animal be useful for
developmental studies of far more complex animals? Sea urchins serve as
essential model organisms for this research mainly due to the large number
of easily collected gametes produced. The external fertilization of eggs,
occurring outside the sea urchin's body, allows researchers to readily
observe development in the laboratory. Sea urchins also follow a
developmental process similar to vertebrate species.
Upon their release into seawater, sea urchin eggs produce molecules on
their jelly coats that attract sperm. Once the head of a sperm comes in
contact with an egg's jelly coat, proteins in the jelly trigger an acrosomal
reaction. This response starts with the release of hydrolytic enzymes from
the apex of the sperm's head, or acrosome. The hydrolytic enzymes destroy
a portion of the egg's jelly coat, which allows a filamentous structure, called
the acrosomal process, to extend from the sperm's head, elongate, and
diffuse through the jelly coat. The acrosomal process contains the proteins
that attach to specific receptors on the plasma membrane of the egg. The
binding between acrosomal proteins and egg receptors is species specific (it
is often referred to as a "lockandkey" type of binding), which ensures that
only gametes of the same species can fuse.
A similar fertilization process occurs in mammals (Figure 3). When a sperm
approaches the surface of a mammalian egg, it first encounters the zona
pellucida, a layer of extracellular matrix that surrounds the egg. Proteins on
the head of the sperm bind to ZP3, which is both a component of the zona
pellucida and the egg's receptor for the sperm. This binding activates the
acrosomal reaction, releasing enzymes that break down the zona pellucida.
This enables the sperm to reach the egg's plasma membrane, where the two
membranes fuse together. Binding of the two plasma membranes first
initiates a depolarization of the membrane, which is a fast block to
polyspermy, as it is in sea urchin eggs. This is followed by a cortical reaction
that serves as a slow block to polyspermy.
Test Yourself
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145869/1
3/9
4/7/2015
Embryonic Development | Principles of Biology from Nature Education
Test Yourself
How would fertilization be affected if an acrosome did not contain hydrolytic proteins?
Submit
What mechanism ensures that only one sperm will fertilize an egg when
many sperm cells are swimming toward the egg? That is, what prevents
polyspermy — the fertilization of an egg by two or more sperm? Once the
acrosomal proteins and egg receptors join, the plasma membranes of the
sperm and the egg fuse. This allows the entry of the sperm nucleus into the
egg cytoplasm. Membrane fusion also opens ion channels present on the
plasma membrane of the egg, allowing sodium ions to move into the egg.
The presence of these ions decreases the membrane potential of the plasma
membrane; that is, it depolarizes the membrane. Depolarization happens
very quickly, taking only a few seconds. The change in the charge of the
plasma membrane prevents other sperm cells from binding to the egg, acting
as a fast block to polyspermy.
Depolarization provides a very brief block to polyspermy. Vesicles located
just underneath the egg's plasma membrane, in a region of the cytoplasm
called the cortex, further extend this blocking effect. Once gametes fuse,
these enzymecontaining vesicles quickly merge with the egg's plasma
membrane. Enzymes from cortical vesicles enter the perivitelline space,
which is the region between the egg's plasma membrane and its extracellular
sheath, called the vitelline layer. The vitelline layer then separates from the
egg and hardens into a fertilization envelope. The change in egg structure
due to the action of vesicle enzymes is called the cortical reaction or the
slow block to polyspermy. Other enzymes dismantle spermbinding receptors
on the egg's plasma membrane, especially those close to the area where the
sperm attached to the egg. All these events help block polyspermy (Figure
3).
Figure 3: Fusion of mammalian sperm and egg and the subsequent
cortical reaction.
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145869/1
4/9
4/7/2015
Embryonic Development | Principles of Biology from Nature Education
(a) A sperm approaches the surface of a mammalian egg. (b) The sperm
first encounters the zona pellucida, a layer of extracellular matrix that
surrounds the egg. (c) Proteins on the head of the sperm bind to ZP3, its
receptor on the egg. This activates the acrosomal reaction, releasing
enzymes that break down the zona pellucida and enable the sperm to
reach the egg's plasma membrane. (d) Binding of the two plasma
membranes first initiates a fast block to polyspermy followed by a cortical
reaction that serves as a slow block to polyspermy.
© 2014 Nature Education All rights reserved.
After fertilization, cleavage occurs.
After fertilization, the zygote divides repeatedly during a stage known as cleavage. This division consists largely of the
DNA synthesis and mitosis phases of the cell cycle, bypassing the protein synthesis and growth stages. Because there
is no cell growth, cleavage results in the formation of small cells called blastomeres. Cleavage begins with the
relatively large singlecelled zygote. Since there is little protein synthesis during cleavage, the cytoplasm of the zygote
becomes portioned into the blastomeres. This results in blastomeres that may contain distinct cytoplasmic proteins and
mRNA. Cleavage, thus, is the first step in creating asymmetry, which is essential for embryonic development.
Test Yourself
Why is the total number of blastomeres at any stage of cleavage theoretically always even
numbered?
Submit
The zygotes of most animal species, including those of frogs and humans,
possess a yolk. This organic material supplies nutrients to the embryo during
development. The yolk is concentrated at one region of the zygote, called the
vegetal pole. The region of the zygote opposite the vegetal pole is called the
animal pole.
During cell division, a groove known as a cleavage furrow develops as a
single cell separates into two. The first two cleavage furrows of frog embryos
develop parallel to the meridian, forming a line between the vegetal and the
animal poles. The third cell division occurs perpendicular to the first two
divisions. The yolk impedes the process of cell division, so in the eightcell
frog embryo the cells at the vegetal pole are larger than the four at the
animal pole because the yolk pushes the cleavage furrow closer to the
animal pole. This asymmetrical cleavage pattern continues during
subsequent cell divisions, so that the cells closer to the animal pole are
smaller than the cells near the vegetal pole (Figure 4).
After several more rounds of cell division, the cells produced by cleavage
form a sphere, called the blastula, that contains a fluidfilled cavity called the
blastocoel. Depending on the type of animal, the blastula can be made of
from 128 cells to thousands of cells. Formation of the blastula marks the end
of cleavage.
Experimental studies on blastomeres make use of dyes that label specific
cells of a blastula. As the blastula undergoes the next stages of
development, the labeled cells rearrange, migrating to their destination within
the differentiating embryo. This experimental technique, called fate mapping,
allows scientists to track various processes that each blastomere undergoes
during embryonic development.
Figure 4: Cleavage and the development of the blastocoel.
The early stages of embryonic development feature multiple divisions.
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145869/1
5/9
4/7/2015
Embryonic Development | Principles of Biology from Nature Education
© 2014 Nature Education All rights reserved.
Transcript
During gastrulation, three germ layers form in an embryo.
In the next stage of embryonic development, the blastula undergoes
substantial cellular rearrangements that result in the formation of the germ
layers that will eventually develop into tissues and organs. The organization
of these germ layers influences cell and tissue arrangements that give shape
to an animal's body. This process of cell movements and reorganization is
called gastrulation. The embryo composed of these layers of cells is called
a gastrula.
Each germ layer in the gastrula will give rise to specific types of cells.
Animals in which the gastrula is a twolayered structure are called
diploblasts, while animals that generate three germ layers are referred to as
triploblasts. Diploblasts consist of the outer ectoderm and the inner
endoderm. Triploblasts possess both ectoderm and endoderm layers, and a
third middle layer called mesoderm. Simple animals, including many radially
symmetrical species, are diploblasts.
What role do these germ layers perform in the developing embryo? One
might assume that the ectoderm forms the exterior of animal, the mesoderm
forms the internal organs, and the endoderm forms the digestive cavity.
However, the actual fate of each germ layer is more complex. Some organs
originate from specific germ layers, while others develop from combinations
of germ layers. For example, most endocrine glands of mammals arise from
the endodermal layer, while the adrenal glands develop from both the
ectodermal and mesodermal germ layers.
In sea urchins, gastrulation begins when some cells separate from the
vegetal pole of the blastula and disperse throughout the blastocoel as
mesenchyme cells. Other cells at the vegetal pole elongate in shape,
causing that end of the embryo to dip inward in a process called
invagination. The shape of the entire embryo changes as a result of the
movement and modification of these cells. With further modification of nearby
cells, the invagination develops into a deeper indentation called the
archenteron. The initial opening formed by the archenteron, known as the
blastopore, later develops into the anus of the animal. The archenteron
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145869/1
6/9
4/7/2015
Embryonic Development | Principles of Biology from Nature Education
reaches and fuses with the inner lining of the blastocoel. This region
eventually develops into the mouth. As gastrulation nears completion, the
surface cells of the embryo form the ectoderm. The cells on the surface of
the archenteron develop into the endoderm, while the cells between these
layers become the mesoderm (Figure 5).
The steps described in the previous paragraph are for animals called
deuterostomes, which means "second mouth." Gastrulation follows similar
steps in amphibians and mammals, with modifications allowing for the more
intricate arrangements of blastomeres in these embryos (Figure 5).
Amphibians and mammals are two more examples of deuterostomes. There
is an alternative gastrulation pattern in which the blastopore becomes the
mouth and the second opening of the archenteron becomes the anus.
Animals that develop in this fashion are called protostomes ("first mouth")
and include arthropods, nematodes, annelids, and molluscs.
Figure 5: Gastrulation.
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145869/1
7/9
4/7/2015
Embryonic Development | Principles of Biology from Nature Education
Gastrulation is a stage of embryonic development featuring marked
differentiation of cells. Gastrulas from a sea urchin, frog, and human are
shown. Note the similarities and differences between them.
© 2014 Nature Education All rights reserved.
Following gastrulation the central nervous system begins to form.
In organogenesis, cells and tissues of the developing embryo start to form
into organs. The central nervous system develops in vertebrate species
through the process of neurulation. Neurulation starts with mesodermal
cells forming a notochord, a column of cells positioned along the dorsal
region of the embryo. These cells produce signaling molecules that trigger
the ectodermal cells above to differentiate into elongated cells that are
collectively known as the neural plate. The neural plate folds inward until its
edges, called neural folds, pinch inward toward each other. The fusing of the
neural folds creates a hollow neural tube that extends from the anterior to
the posterior regions of the developing embryo (Figure 6). The neural tube
serves as the forerunner of the brain and the spinal cord.
In vertebrate species, two groups of cells involved in neurulation go on to
develop into other tissues and organs. The neural folds detach from the
overlying ectoderm as the neural tube is formed. One group of cells, the
neural crest cells, emigrates from the dorsal neural tube as mesenchymal
cells. These cells continue to migrate to various regions of the embryo and
ultimately form teeth, parts of the skull, and components of the peripheral
nervous system, among other parts of the animal. Sequential blocks of
mesodermal cells, called somites, form along the notochord. Somites
develop into a variety of tissues, including muscle and bone. Vertebrae, for
example, develop from somites.
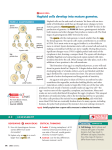
Figure 6: Neural tube formation.
Neurulation starts with infolding of the neural plate. The neural folds fuse,
forming a neural tube.
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145869/1
8/9
4/7/2015
Embryonic Development | Principles of Biology from Nature Education
© 2014 Nature Education All rights reserved.
Test Yourself
How does the sequential arrangement of blocks of somites in the developing embryo aid in
the development of adult vertebrates?
Submit
Organ formation occurs once the central nervous system is in place.
Organogenesis, or the formation of organs, follows a similar pattern in most
vertebrates. The first organs developed are those of the central nervous
system, including the brain, the spinal cord, and other related neural
structures such as the eyes, nerves, and ganglia. Soon after the central
nervous system emerges, other organs develop. The heart forms right after
the development of the brain. The embryonic heart mainly consists of muscle
tissues that pump blood for circulation across the animal's body. The organs
of the respiratory system, namely the lungs, bronchus, and bronchioles,
develop next. The initial lung structures are not capable of functioning
independently as respiratory organs, but through continued cell and tissue
differentiation, the lungs will be able to function once they are needed for the
animal's survival. The digestive system, consisting of the stomach,
intestines, and anus, also develops during organogenesis. These organs
also acquire the capacity to function during later stages of development.
What factors stimulate the germ layers to undergo organogenesis?
Morphogens are molecules that convey messages regarding a tissue's
position within the developing embryo and information about its neighboring
tissues. These morphogens are critical signaling molecules that assist in
organogenesis. Through this cellular mechanism of signaling, each cell is
apprised of which activities should or should not be performed. The end
result of organogenesis is an animal with a complete set of organs, each of
which consists of differentiated cells.
IN THIS MODULE
Stages of Embryonic Development
Summary
Test Your Knowledge
WHY DOES THIS TOPIC MATTER?
Stem Cells
Stem cells are powerful tools in
biology and medicine. What can
scientists do with these cells and their
incredible potential?
Cancer: What's Old Is New Again
Is cancer ancient, or is it largely a
product of modern times? Can
cuttingedge research lead to prevention
and treatment strategies that could make
cancer obsolete?
SCIENCE ON THE WEB
Fast Forward on Brain Development
Fast forward on brain development
page 831 of 989
2 pages left in this module
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145869/1
9/9
4/7/2015
Embryonic Development | Principles of Biology from Nature Education
Principles of Biology
162 Embryonic Development
contents
Test Your Knowledge
1. What organ system develops first during organogenesis?
cardiovascular system
respiratory system
nervous system
digestive system
All of the organ systems develop simultaneously.
2. A developing organism forms a neural plate. What is the next step in neural plate
development?
It forms the vertebrae.
It becomes a somite.
It will fold into a tube.
It becomes the brain.
It turns into the blastopore.
3. What does the very tip of the sperm head contain that helps it penetrate the egg?
gamete
lipids
jelly coat
hydrolytic enzymes
chromosomes
4. Which of the following events occurs in a stage prior to gastrulation?
germ cell layers form
the gastrula takes shape
the cortical reaction takes place
the ectoderm develops
the endoderm develops
5. Neural crest cells emigrate from the dorsal neural tube as ___ cells, which
eventually form the teeth and skull. The ___, or mesodermal cells, develop into
tissues such as muscle and bone tissues.
ectodermal, somites
endodermal, ectoderms
mesenchymal, ectoderms
mesenchymal, somites
mesenchymal, endoderms
6. Which of the following actions might indicate that a fertilized egg failed to develop
normally after several days?
The egg cell had increased in size.
The egg divided into several small cells.
A hole formed in the center of the new cells.
A deep indentation formed in the cell.
All answers are correct.
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145869/3
1/2
4/7/2015
Embryonic Development | Principles of Biology from Nature Education
Submit
IN THIS MODULE
Stages of Embryonic Development
Summary
Test Your Knowledge
WHY DOES THIS TOPIC MATTER?
Stem Cells
Stem cells are powerful tools in
biology and medicine. What can
scientists do with these cells and their
incredible potential?
Cancer: What's Old Is New Again
Is cancer ancient, or is it largely a
product of modern times? Can
cuttingedge research lead to prevention
and treatment strategies that could make
cancer obsolete?
SCIENCE ON THE WEB
Fast Forward on Brain Development
Fast forward on brain development
page 833 of 989
http://www.nature.com/principles/ebooks/principlesofbiology104015/29145869/3
2/2