Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Paolo Macchiarini wikipedia , lookup
Somatic cell nuclear transfer wikipedia , lookup
Cell culture wikipedia , lookup
Bone morphogenetic protein 4 wikipedia , lookup
Cell encapsulation wikipedia , lookup
Sonic hedgehog wikipedia , lookup
Regeneration in humans wikipedia , lookup
Human embryogenesis wikipedia , lookup
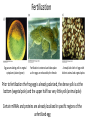
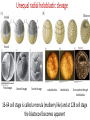
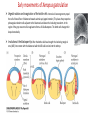
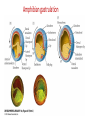
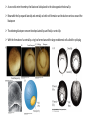
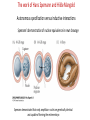
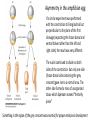
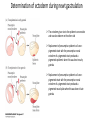
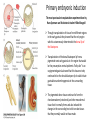
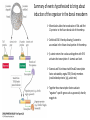
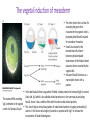
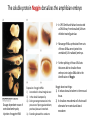
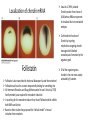
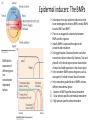
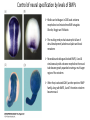
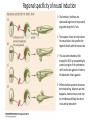
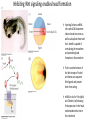
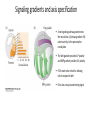
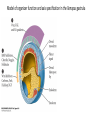
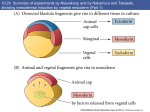
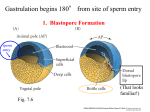
Axis Specification and Patterning III Axis specification in the frog embryo (Xenopus laevis) Amphibians were the first models for studying vertebrate development Studying these organisms has led to formulation of “key” questions and also provided some tentative answers to these questions allowing vertebrate developmental biology to flourish Developmental history of the leopard frog Fertilization Egg accumulating yolk in vegetal cytoplasm (stained green) Fertilization is external and takes place as the eggs are released by the female A newly laid clutch of eggs with distinct animal and vegetal poles Prior to fertilization the frog egg is already polarized, the dense yolk is at the bottom (vegetal pole) and the upper half has very little yolk (animal pole) Certain mRNAs and proteins are already localized in specific regions of the unfertilized egg Fertilization can occur anywhere in the animal hemisphere The point of sperm entry determines dorsalventral polarity The point of sperm entry marks the belly (ventral) side of the embryo The point 180° opposite to the point of sperm entry marks the dorsal (spinal) side The sperm centriole organizes the microtubules into parallel tracks in the vegetal cytoplasm The cortical cytoplasm now is separated from the yolky central cytoplasm and rotates30° with respect to it This exposes the grey cytoplasm (grey crescent) opposite the sperm entry point. Gastrulation begins here. Unequal radial holoblastic clevage First clevage Second clevage Fourth clevage early blastula late blastula Cross section through late blastula 16-64 cell stage is called a morula (mulberry like) and at 128 cell stage the blastocoel becomes apparent The blastocoel serves two functions: It can change its shape to facilitate cell migration during gastrulation. It prevents premature interaction between cell below with the cells above. Mid-blastula transition preparing for gastrulation • The major transcription of zygotic genes begins such that different genes are transcribed in different cells • The cell cycle acquires gap phases • Blastomeres acquire the capacity to become motile Goals of amphibian gastrulation To bring inside the embryo those areas destined to form the endodermal organs. To surround the embryo with cells capable of forming the ectoderm. To place the mesodermal cells in the proper positions between the ectoderm and endoderm. Fate map at mid-blastula stage Early movements of Xenopus gastrulation Vegetal rotation and invagination of the bottle cells: Internal cell rearrangements propel the cells of dorsal floor of blastocoel towards animal cap (vegetal rotation). This places the prospective pharangeal endoderm cells adjacent to the blastocoel and above the involuting mesoderm. In the region of the grey crescent cells invaginate to form a slit-like blastopore. The bottle cells change their shape dramatically. Involution at the blastopore lip: After the bottle cells have brought the involuting marginal zone (IMZ) into contact with the blastocoel wall the IMZ cells involute into the embryo. Amphibian gastrulation Amphibian gastrulation As new cells enter the embryo the blastocoel is displaced to the side opposite the dorsal lip Meanwhile the lip expands laterally and ventrally as bottle cell formation and involution continue around the blastopore The widening blastopore crescent develops lateral lips and finally a ventral lip With the formation of a ventral lip a ring has formed around the large endodermal cells called the yolk plug The work of Hans Spemann and Hilde Mangold Autonomous specification versus inductive interactions Spemann’s demonstration of nuclear equivalence in newt cleavage Spemann demonstrated that early amphibian nuclei are genetically identical and capable of forming the entire embryo Asymmetry in the amphibian egg If a similar experiment was performed with the constriction still longitudinal but perpendicular to the plane of the first cleavage (separating the future dorsal and ventral halves rather than the left and right sides) the result was very different. The nuclei continued to divide on both sides of the constriction but only one side (future dorsal side containing the grey crescent) gave rise to a normal larva. The other side formed a mass of unorganized tissue which Spemann named “the belly piece” Something in the region of the grey crescent was essential for proper embryonic development The grey crescent region was essential for embryonic development How? Fate map showed that grey crescent region gives rise to the dorsal blastopore lip where gastrulation is initiated. Spemann speculated: The importance of the grey crescent region lies in its ability to initiate gastrulation. Critical changes in cell potency occur during gastrulation. Determination of ectoderm during newt gastrulation The ectoderm gives rise to the epidermis on one side and neural ectoderm on the other side Replacement of presumptive epidermis of a nonpigmented strain with the presumptive neural ectoderm of a pigmented strain produced a pigmented epidermis when this was done in early gastrula Replacement of presumptive epidermis of a nonpigmented strain with the presumptive neural ectoderm of a pigmented strain produced a pigmented neural plate when this was done in late gastrula Conclusion: In the early gastrula the transplanted tissue undergoes conditional development because the ultimate fate depends on the location in the embryo. In the late gastrula the transplanted tissue exhibits autonomous development independent of their final location after transplantation, i.e. their fate was already determined. Primary embryonic induction The most spectacular transplantation experiment done by Hans Spemann and his doctoral student Hilde Mangold Through transplantation of tissues from different regions in the early gastrula they showed that the only region which is autonomously determined is the dorsal lip of the blastopore. Transplantation of the dorsal blastopore lip from a pigmented newt early gastrula to the region that would be the presumptive ventral epidermis “belly skin” in an unpigmented gastrula showed that this tissue not only continued to be the dorsal blastopore lip it could initiate gastrulation and embryogenesis in the surrounding tissue. The pigmented donor tissue continued to form the chordamesoderm (notochord) and other mesodermal tissue that it normally forms and also induced the changes in the surrounding host cells to make tissues that they normally would not have made. Spemann’s organizer: Dorsal blastopore lip cells and their derivatives the notochord and head endomesoderm Called an “organizer” because: They induce the host’s ventral tissues to change their fates to form a neural tube and dorsal mesoderm They organize the host and donor tissues into a secondary embryo with clear anterior-posterior and dorsal-ventral axes. However Spemann’s work together with the work of others has shown that this interaction of the chordamesoderm and the ectoderm is not sufficient to organize the entire embryo. Rather it initiates a series of sequential inductive events. This key induction in which the progeny of the dorsal lip induce the dorsal axis and the neural tube is known as the primary embryonic induction. Questions raised after Spemann’s organizer was identified ??? How did the organizer get its properties? What makes the dorsal blastopore lip different from the rest of the embryo? What factors secreted from the organizer cause the formation of the neural tube and specification of the anterior-posterior and dorsal-ventral axes? How did the different parts of the neural tube along the A-P axis get established? Molecular mechanism of amphibian axis formation How does the organizer form? Isolated animal cap cells become ectoderm, isolated marginal/equatorial cells become mesoderm and isolated vegetal cells become gut-like tissue If animal cap cells are put in direct contact with vegetal cells they become mesoderm. Thus vegetal cells secrete factors that induce the mesoderm. Experiments done by Pieter Niewkoop and by Osamu Nakamura and Takasaki Nieuwkoop center: The dorsal most vegetal cells of the blastula capable of inducing the organizer Gimlich and Gerhart in 1984, transplanted the dorsalmost vegetal blastomere from one blastula to into the ventral vegetal side of the other blastula two embryonic axes formed Dale and Slack in 1987 recombined single vegetal blastomeres with uppermost animal tier of a fluorescently labelled blastula of same stage, the dorsalmost vegetal cell induced animal pole to become dorsal mesoderm Transplantation and recombination experiments on Xenopus embryos demonstrate that the vegetal cells underlying the prospective dorsal blastopore lip region are responsible for initiating gastrulation What gives the dorsal most vegetal cells their special properties? The Wnt pathway is involved and a major candidate is β-catenin WHY? Two types of experiments support the presence of a Wnt activity in the Nieuwkoop center: 1. Vegetal explants need the maternal Wnt pathway to induce expression of organizer and dorsal mesoderm genes in recombined animal caps or equatorial explants. 2. Components of the Wnt pathway induce a secondary axis, or rescue the axis of UV-ventralized embryos, when injected in vegetal cells that do not themselves form dorsal mesoderm, and thus behave like a Nieuwkoop center β-catenin is found throughout the embryo as it is translated from the maternal mRNA but it preferentially accumulates in the presumptive dorsal side. In the vegetal balstomeres the presumptive dorsal side shows nuclear localization whereas the ventral side does not. Model of the mechanism by which the Disheveled protein stabilizes -catenin in the dorsal portion of the amphibian egg Three proteins, Wnt11, GSK-3 binding protein (GBP) and Dishevelled (Dsh) are translocated from the vegetal side of the egg to the future dorsal side of the embryo during fertilization. -catenin is targeted for destruction by GSK-3. In fact if GSK-3 activity in ventral cells is blocked a secondary axis forms. GSK-3 can be inactivated by GBP and Dsh. These two proteins release GSK-3 from the degradation complex and prevent it from binding -catenin and targeting it for destruction. GBP binds to Kinesin a motor that moves towards the growing end of microtubules i.e. away from the site of sperm entry. Dsh binds to GBP and is thus transported along with it. Thus in the future dorsal side of the embryo GBP and Dsh inactivate GSK-3 and stabilize -catenin However mere translocation of GBP and Dsh is not sufficient to stabilize -catenin. Wnt activation is required. Wnt activity is provided by Wnt 11. The mRNA for Wnt 11 is also localized to the vegetal cortex and is transported to the future dorsal side by cortical rotation Summary of events hypothesized to bring about induction of the organizer in the dorsal mesoderm Microtubules allow the translocation of Dsh and Wnt 11 proteins to the future dorsal side of the embryo. Dsh binds GSK-3 thereby allowing -catenin to accumulate in the future dorsal portion of the embryo. -catenin enters the nucleus and together with Tcf-3 activates the transcription of siamosis and twin. Siamosis and Twin interact with Smad2 transcription factor activated by vegetal TGF-β family members (nodal related proteins, Vg1, activin etc.) Together these transcription factors activate “organizer” specific genes such as goosecoid, chordin, noggin etc. Specification of the germ layers The vegetal cells have two major functions: 1. To differentiate into endoderm 2. To induce the cells immediately above them to become mesoderm The mechanism of this “bottom up” specification resides in a set of mRNAs tethered to the vegetal cortex. The maternal mRNA for the transcription factor VegT is critical for both endoderm and mesoderm specification. The VegT mRNA is translated shortly after fertilization and in the vegetal blastomeres turns on the Sox17 transcription factor required to specify endoderm fate. VegT also induces the expression of Nodal paracrine factors in the vegetal cells which act on the cells above them to activate Smad2 by phosphorylation in these cells which then induce the expression of eomesodermin and Brachyury which impart a mesodermal fate to the cells lying above the vegetal cells. The Eomesodermin and pSmad2 co-operate to transcribe zygotic VegT thus setting up a positive feedback loop needed to maintain mesodermal fate. Another maternal mRNA for Vg1 localized to the dorsal region of the vegetal pole is translated. Vg1 is a Nodal-like protein which induces dorsal mesodermal fate to the cells above it. The vegetal induction of mesoderm The other factor that is critical for activating the genes that characterize the organizer cells is phosphorylated Smad2 required for mesoderm formation. Smad2 is activated in the mesodermal cells when it becomes phosphorylated downstream of the Nodal related paracrine factors secreted by the vegetal cells. Activated Smad2 functions as a transcription factor with a partner. In the late blastula there is a gradient of Nodal- related proteins form dorsal (high) to ventral (low) side. Vg1 which is also a Nodal-related protein acts in the same way by activating The maternal RNA encoding Smad2, hence it has a additive effect with the other nodal-related proteins. Vg1 is tethered to the vegetal The dorsal high to ventral low gradient of nodal related proteins is largely produced by βcortex of a Xenopus Oocyte catenin. In the future dorsal region β-catenin co-operates with Veg-T to increase the transcription of Nodal-related genes. The dorsal high to ventral low gradient of Nodalrelated proteins is largely produced by β-catenin. In the future dorsal region β catenin co-operates with Veg-T to increase the transcription of Nodal-related genes (Xnr1, 5 and 6). The more ventral blastomeres in the endoderm lack the expression of these Nodal-related genes. In the region that will become the most anterior portion of the organizer-pharangeal endodermhigher levels of Nodal-related proteins produces highest activity of pSmad2. These induces the transcription of hhex gene. Hhex together with Twin and Siamosis (induced by β-catenin) specify pharangeal endoderm to be foregut and to induce anterior brain development in the ectoderm above. Slightly lower levels of pSmad2 activate goosecoid expression in cells that will become prechordal mesoderm and notochord. Even lower levels of pSmad2 result in the formation of ventral and lateral mesoderm. Mesodermal regions with little or no Xnr have high levels of BMP4 and XWnt8. In summary, the intersection between the vegetal pathway activating the Nodal-related genes and pSmad2 in the mesodermal cells above them with the β-catenin activated Twin and Siamosis on the future dorsal side leads to the “organizer” being specified. The Nieuwkoop center is in the endodermal cells. The “organizer” is in the mesoderm comprising of four different tissues: pharangeal endoderm, head mesoderm (prechordal plate), dorsal mesoderm (notochord) and the dorsal blastopore lip. The properties of the “organizer” The ability to self differentiate into dorsal mesoderm ( prechordal plate, chordamesoderm etc.) The ability to dorsalize the surrounding mesoderm into paraxial (somite forming) mesoderm when it would have differentiated into ventral mesoderm. The ability to dorsalize the ectoderm and induce formation of the neural tube. The ability to induce the movement of gastrulation. What are the factors produced by the organizer? Evidence showing that the organizer produces soluble factors which impart neural fate to the ectoderm If notochord fails to migrate beneath the ectoderm it will not become neural tissue rather it will become epidermis. Transfilter studies where neural structures were induced in presumptive newt ectoderm by newt dorsal lip tissue separated from ectoderm by nucelopore filter with a pore diameter of 0.05 mm. Induction of the neural ectoderm and dorsal mesoderm: BMP inhibitors Neural induction ? Scientists were looking for molecules secreted from the organizer that induce neural fate in the ectoderm. Wrong idea! As it turns out the default fate for the ectoderm is to become neural tissue. BMPs induces the ectoderm to become epidermis. The organizer secretes BMP antagonists which block this and allow a certain portion of the ectoderm to acquire its default fate. Three of the major BMP inhibitors secreted by the “organizer” are Noggin, Chordin and Follistatin The soluble protein Noggin dorsalizes the amphibian embryo In 1992 Smith and Harland constructed a cDNA library from dorsalized (Lithium chloride treated) gastrulae. Messenger RNAs synthesizied from sets of these cDNAa were injected into ventralized (UV-irradiated) embryos. Further splitting of those cDNA sets that were able to dorsalize these embryos into single cDNAs led to the identification of Noggin. Dosage-dependent rescue of ventralized embryos by injection of noggin mRNA Expression of noggin mRNA: i) Accumulted in dorsal marginal zone ii) In the dorsal blastopore lip iii) During convergent extension in the precursors of pharyngeal endoderm, prechoral plate and notochord. iv) Extending beneath the ectoderm. Noggin does two things: 1) It induces dorsal ectoderm to form neural tissue. 2) It dorsalizes mesodermal cells that would otherwise form ventral and lateral mesoderm. Localization of chordin mRNA Sasai et al. 1994, isolated Chordin protein from clones of cDNA whose RNA were present in dorsalized but not ventralized embryos. Confirmed the function of Chordin by injecting morpholinos targeting chordin message which blocked secondary axis formation by the organizer graft. Follistatin Follistatin is also transcribed in the dorsal blastopore lip and the notochord Follistatin was found in a screen unexpectedly looking for something else Ali Hemmanti-Brivanlou and Doug Melton wanted to see if Activin (a TGFβ family member) was required for mesoderm induction In searching for the mesoderm inducer they found Follistatin which inhibits both BMPs and Activin. Based on their studies they proposed the “default model” of neural induction from ectoderm. Of all the organizer genes chordin is the one most acutely activated by β-catenin. Epidermal inducers: The BMPs BMP4 elicilts expression of different genes in a concentration dependent fashion In Xenopus the major epidermal inducers are the bone morphogenetic proteins (BMPs) mostly BMP4 but also BMP2 and BMP7. There is an antagonistic relationship between BMPs and the organizer. Initially BMP4 is expressed throughout the ectoderm and mesoderm. During gastrulation Goosecoid protein and other transcription factors induced by Siamosis, Twin and pSmad2 in the dorsal region repress thanscription of bmp4 and wnt8 expression in the dorsal region. In the ectoderm BMPs repress the genes (such as neurogenin) involved in neural tissue formation. In the mesoderm graded levels of BMP4 activate different mesodermal genes: i) Absence of BMP4 specifies dorsal mesoderm ii) A low amount specifies intermediate mesoderm iii) High amount specifies lateral mesoderm Control of neural specification by levels of BMPs Khokha and colleagues in 2005 used antisense morpholinos to eliminate three BMP antagoists Chordin, Noggin and Follistatin. The resulting embryos had catastrophic failure of dorsal development,lacked neural plate and dorsal mesoderm. Reversdale and colleagues blocked BMP2, 4 and & simultaneously with antisense morpholinos the neural tube became greatly expanded covering a much larger region of the ectoderm. When they inactivated ADM ( another protein of BMP family) along with BMP2, 4 and 7 the entire ectoderm became neural Regional specificity of neural induction The forebrain, hindbrain and spinocaudal regions must be properly organized along the A-P axis The organizer tissue not only induces the neural tube it also specifies the regional identity with the neural tube This was demonstrated by Otto mangold in 1933, by transplanting fur successive region of the archenteron roof from the late gastrula of newt to the blastocoel of early gastrula. Different anterio-posterior structures were induced e.g. balancers and oral apparatus, head structures nose, eye etc, hindbrain and finally the dorsal trunk and tail mesoderm Temporal specificity of induction Mangold also demonstrated that transplantation of dorsal lip from early gastrula to blastocoel of another early gastrula led to formation of secondary heads Whereas transplantation of dorsal lip from a late gastrula to and early gastrula led to formation of secondary tails. What paracrine factors from the organizer provide this specificity? Paracrine factor antagonists from the organizer are able to block specific paracrine factors to distinguish head from tail Inhibiting Wnt signaling enables head formation Injecting Cerberus mRNA into ventral D4 blastomere induces head structures as well as a duplicate heart and liver. Xwnt8 is capable of ventralizing the mesoderm and preventing head formation in the ectoderm Frzb is secreted version of the Wnt receptor Frizzled and hence can sequester Wnt ligands and prevent them from acting mRNA in situ for Frzb (dark) and Chordin (red) showing Frzb expression in the head endomesoderm but not in the notochord Signaling gradients and axis specification A wnt signaling pathway posteriorizes the neural tube. (A) shows gradient of βcatenin activity in the presumptive neural plate. The Wnt gradient provides A-P polarity and BMP gradient provides D-V polarity. FGFs seem to be critical for allowing cells to respond to Wnt RA is also a major posteriorizing signal Model of organizer function and axis specification in the Xenopus gastrula