Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Gaseous signaling molecules wikipedia , lookup
Catalytic triad wikipedia , lookup
Mitogen-activated protein kinase wikipedia , lookup
Oligonucleotide synthesis wikipedia , lookup
Metalloprotein wikipedia , lookup
Photosynthesis wikipedia , lookup
Photosynthetic reaction centre wikipedia , lookup
Paracrine signalling wikipedia , lookup
Basal metabolic rate wikipedia , lookup
Microbial metabolism wikipedia , lookup
Metabolic network modelling wikipedia , lookup
Nicotinamide adenine dinucleotide wikipedia , lookup
Enzyme inhibitor wikipedia , lookup
Adenosine triphosphate wikipedia , lookup
Lactate dehydrogenase wikipedia , lookup
Fatty acid synthesis wikipedia , lookup
Blood sugar level wikipedia , lookup
Biosynthesis wikipedia , lookup
Oxidative phosphorylation wikipedia , lookup
Biochemical cascade wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
Phosphorylation wikipedia , lookup
Amino acid synthesis wikipedia , lookup
Fatty acid metabolism wikipedia , lookup
Citric acid cycle wikipedia , lookup
Glyceroneogenesis wikipedia , lookup
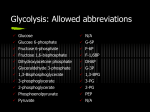
KAPITOLA 8 Energetický metabolismus I • glykolýza a kvašení • pentózový cyklus • glukoneogeneze • lokalizace reakcí v buňce • fyziologické aspekty bioenergetiky Glycogen, starch, sucrose Major pathways of glucose utilization in cells of higher plants and animals. Although not the only possible fates for glucose, these three pathways are the most significant in terms of the amount of glucose that flows through them in most cells. Storage Glucose oxidation via pentose phosphate pathway Ribose-5-phosphate oxiadation via glycolysis Pyruvate Three possible catbolic fates of the pyruvate formed in the payoff phase of glycolysis. Pyruvate also serves as a precursor in many anabolic reactions, not shown here. •Primary metabolism of glucose Glycolysis The two phases of glycolysis. For each molecule of glucose that passes through the preparatory phase (a), two molecules of glyceraldehyde3-phosphate are formed; both pass through the payoff phase (b). Pyruvate is the end product of the second phase under aerobic conditions, but under anaerobic conditions pyruvate is reduced to lactate to regenerate NAD+. For each glucose molecule, two ATP are consumed in the preparatory phase and four ATP are produced in the payoff phase, giving a net yield of two molecules of ATP per one of glucose converted to pyruvate. Keep in mind that each phosphate group, represented here as P, has two negative charges (- PO32-). Entry of glycogen, starch, disaccharides, and hexoses into the preparatory stage of glycolysis. Covalent regulation Allosteric regulation Covalent and allosteric regulation of glycogen phosphorylase in muscle. (a) The enzyme has two identical subunits, each of which can be phosphorylated by phosphorylase b kinase at Ser14 to give phosphorylase a, a reaction promoted by Ca2+. Phosphorylase a phosphatase, also called phosphoprotein phosphatase-1, removes these phosphate groups, inactivating the enzyme. Phosphorylase b can also be activated by noncovalent binding of AMP at its alosteric sites. Conformational changes in the enzyme are indicated schematically. Liver glycogen phosphorylase undergoes similar a and b interconversions, but has different regulatory mechanisms. (b) The three-dimensional structure of the enzyme from muscle. The two subunits (gray and blue) of the glycogen phosphorylase a dimer, showing the location of the phosphates (orange) attached to the Ser14 residues (red) in each.. In phosphorylase b, the amino-terminal peptide containing Ser14 is disordered. However, with the attachment of negatively charged phosphate group at Ser14 this peptide folds toward several nearby (positively charged) Arg residues (pink), forcing compensatory changes in regions distant from Ser14 and activating the enzyme. AMP, the allosteric activator of phosphorylase b, binds very near Ser14. On the back side of the enzyme is a deep channel that admits the substrate glycogen to the active site, which is 3,3 nm away from the allosteric site. (c) A close-up view of the region around the phospho-Ser residue; note its proximity to the interface between dimers. Hormonal regulation of glycogen phosphorylase in muscle and liver. A cascade of enzymatic activations leads to activation of glycogen phosphorylase by epinephrine in muscle and by glucagon in liver. When catalysts activate catalysts large amplifications of the initial signal results. • Secondary metabolism of glucose: Pentose phosphate pathway The oxidative reactions of the pentose phosphate pathway, leading to D-ribose5-phosphate and producing NADPH. The nonoxidative reactions of the pentose phosphate pathway convert pentose phosphates back into hexose phosphates, allowing the oxidative reactions to continue. The enzymes transaldolase and transketolase are specific to this pathway; the oher enzymes also serve in the glycolytic or gluconeogenetic pathways. A simplified schematic diagram showing the pathway leading from six pentoses (5C) to five hexoses (6C). Secondary pathways for glucose metabolism through UDP-glucuronate. Energy metabolism (anabolism) Gluconeogenesis The pathway from phosphoenolpyruvate to glucose-6-phosphate is common to the biosynthetic conversion of many different precursors into carbohydrates in animals and plants. bypass #1 Gluconeogenesis bypass #2;3 The opposing pathways of glycolysis and gluconeogenesis in rat liver. The three bypass reactions of gluconeogenesis are shown in orange. Two major sites of regulation of gluconeogenesis are also shown. Alternative paths from pyruvate to phosphoenolpyruvate. The paths differ depending upon the gluconeogenetic precursor (lactate or pyruvate) and are determined by cytosolic requirements for NADH in gluconeogenesis. Gluconeogenesis bypass #1 Liver cells Muscle cells FA/ β – oxidation Triacylglycerols stored in seeds are oxidized to acetyl-CoA and dihydroxyacetone phosphate during germination; both are substrates for gluconeogenesis in plants. Recall that acetyl-CoA is not a substrate for gluconeogenesis in animals. Glyoxylate cycle The conversion of stored fatty acid to sucrose in germinating seeds begins in glyoxysomes, which produce succinate and export it to mitochondria. There it is converted to oxaloacetate by enzymes of the citric acid cycle. Oxaloacetate enters the cytosol and serves as the starting material for gluconeogenesis and the synthesis of succrose, the transported sugar in plants. gluconeogenesis FA/ β – oxidation Triacylglycerols stored in seeds are oxidized to acetyl-CoA and dihydroxyacetone phosphate during germination; both are substrates for gluconeogenesis in plants. Recall that acetyl-CoA is not a substrate for gluconeogenesis in animals. Energy metabolism (Anabolism) Glycogenogenesis The glycogen-branching enzyme glycosyl-(46)-transferase (or amylo (14) to (1 6) transglycosylase) forms a new branch point during glycogen synthesis. The glycogen-branching enzyme glycosyl-(46)-transferase (or amylo (14) to (1 6) transglycosylase) forms a new branch point during glycogen synthesis. !!! Initiating the synthesis of a glycogen particle with a protein primer, glycogenin. Steps 1 through 5 are described in the text. Glycogenin is found within glycogen particles, still covalently attached to the reducing end of the molecule. xxxxx xxxxxx x Energy metabolism Gluconeogenesis