Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Organ-on-a-chip wikipedia , lookup
Cellular differentiation wikipedia , lookup
Cell nucleus wikipedia , lookup
Histone acetylation and deacetylation wikipedia , lookup
List of types of proteins wikipedia , lookup
Transcription factor wikipedia , lookup
Eukaryotic transcription wikipedia , lookup
RNA polymerase II holoenzyme wikipedia , lookup
Promoter (genetics) wikipedia , lookup
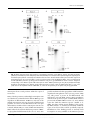
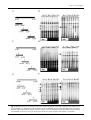
Journal of General Virology (2003), 84, 2909–2920 DOI 10.1099/vir.0.19332-0 Identification and characterization of a cluster of transcription start sites located in the E6 ORF of human papillomavirus type 16 Maiken W. Rosenstierne, Jeppe Vinther, Christina N. Hansen, Martin Prydsoe and Bodil Norrild Correspondence Bodil Norrild [email protected] Received 6 May 2003 Accepted 8 August 2003 Institute of Molecular Pathology, The Protein Laboratory, University of Copenhagen, Panum Institute, Bldg 6.2, Blegdamsvej 3C, DK.2200 Copenhagen N., Denmark Human papillomavirus type 16 (HPV-16) is the prototype strain among the malignant types of HPV in the western world. The main promoter, P97, located in front of the E6 ORF, has been shown to control expression of the oncogenes E6 and E7. These oncogenes are expressed continuously in HPV-16-transformed cells. In contrast to malignant HPV types, non-malignant HPV types have separate promoters driving the expression of E6 and E7. Experiments have shown that the translation of E7 is more efficient from monocistronic than bicistronic transcripts encoding both E6 and E7. Here, identification of a cluster of transcription start sites located in the E6 ORF of HPV-16 is presented. Transcripts from this region contain the E7 ORF as the first reading frame. The cluster consists of multiple transcription start sites located around nt 441. Additional transcription start sites were identified in a cluster around nt 480. A transcription start site has been identified previously at nt 480 but has never been characterized further. The region responsible for transcription activity was mapped to nt 272–448. Mutational analysis showed that initiation of transcription is independent of a TATA-box element, which is consistent with the finding of multiple transcription start sites. Furthermore, it is shown that proteins from HeLa and SiHa nuclear cell extracts bind to the two regions at nt 291–314 and 388–411, and that these two regions influence transcription activity in a cell type-dependent manner. INTRODUCTION Human papillomaviruses (HPVs) are epitheliotropic DNA viruses with a double-stranded circular genome of approximately 8 kbp. During the last 20 years, more than 85 different types of HPV have been identified and characterized, and novel types are still being isolated (zur Hausen, 1999). HPV types are divided into malignant and nonmalignant types according to their ability to transform the host cell. The HPV genome consists of an early region encoding six to seven early proteins [E6, E7, E1, E2, E4 (E8) and E5], a late region encoding two late proteins (L1 and L2) and two regulatory regions, designated the long control region (LCR) and the small non-coding region (Seedorf et al., 1985). In the western world, HPV-16 is considered to be the prototype strain of the malignant types of HPV. The oncoproteins E6, E7 and E5 from malignant types of HPV are responsible for the transformation of the epithelial host cell. In the normal life cycle of the virus, oncoproteins induce unscheduled replication in differentiating keratinocytes to facilitate virus replication (for reviews, see zur Published ahead of print on 27 August 2003 as DOI 10.1099/ vir.0.19332-0. 0001-9332 G 2003 SGM Hausen, 1996; and Zwerschke & Jansen-Dürr, 2000). The E1 and E2 proteins are responsible for DNA replication (reviewed by Phelps et al., 1998) and E2 also functions as a transcriptional regulator (Nishimura et al., 2000; Tan et al., 1994; Ushikai et al., 1994). The E1^E4 protein binds to intermediate filaments and is believed to facilitate the release of virus particles. In addition, the E1^E4 protein is believed to be involved in post-transcription regulation (Doorbar et al., 2000). L1 and L2 are the capsid proteins (reviewed by zur Hausen, 1996). Transcription regulation of the HPV genome is very complex and is controlled largely by the LCR, which binds cellular transcription factors and the HPV E2 protein (reviewed by O’Connor et al., 1995; for recent references, see O’Connor et al., 1998, 2000; Stünkel & Bernard, 1999; and Stünkel et al., 2000). Malignant HPV types have a main promoter located in front of the E6 ORF. This promoter is named P97 in HPV-16 (transcription initiation at nt 97) (Smotkin et al., 1989), P105 in HPV-18 (Thierry et al., 1987) and P99 in HPV-31b (Ozbun & Meyers, 1998). These promoters all produce differentially spliced polycistronic transcripts encoding the early genes. In contrast, non-malignant HPV types have the main promoter located in the E6 ORF in front of Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Thu, 03 Aug 2017 19:56:25 Printed in Great Britain 2909 M. W. Rosenstierne and others the E7 ORF, namely P270 in HPV-6 (Smotkin et al., 1989) and P264 in HPV-11 (Chow et al., 1987). Non-malignant HPV types also have promoters in front of the E6 ORF and these are responsible for the expression of E6, namely P80/ P90 in HPV-6 and P90 in HPV-11 (Karlen et al., 1996; Smotkin et al., 1989; Zhao et al., 1997). A difference between the malignant and non-malignant HPV types is therefore the production of transcripts encoding the E7 protein as the first ORF. In eukaryotes, the translation of the second ORF in a polycistronic transcript is generally very inefficient and it has been shown that the translation efficiency of the HPV-16 E7 oncoprotein is only 10 % compared to translation of a monocistronic E7 transcript (Stacey et al., 1995). It is possible therefore that a minor promoter that produces E7 transcripts as the first reading frame could contribute considerably to the overall expression of E7. In some malignant types of HPV, multiple transcription start sites have been identified in the E6 ORF but they have never been characterized. In HPV-18, transcription start sites were mapped to nt 200, 215, 310 and 455 (Schneider-Gädicke & Schwarz, 1986). In HPV-16, a transcription start site was identified in the E6 region at nt 480 but this was not characterized further (Grassmann et al., 1996). We have previously identified and characterized a new promoter, P542, with a transcription start site at nt 542 in front of the E7 ORF (Braunstein et al., 1999; Glahder et al., 2003). Our previous work indicated the existence of promoter activity located upstream of P542. This led us to search for additional transcription start sites in the HPV-16 E6 ORF that could contribute to the expression of E7. In this study, we identify a novel cluster of transcription start sites mapped to nt 430, 441 and 446. In addition, we find multiple transcription start sites clustered around the transcription start site identified previously at nt 480 (Grassmann et al., 1996). We have mapped the region responsible for transcription activity to nt 272–448 and we show that several nuclear proteins from the SiHa and HeLa cell lines bind to two terminal sites at nt 291–314 and 388–411 of this region. Furthermore, we find that the fragments spanning nt 272– 332 and 388–448 are important for the transcription activity of the region at nt 272–448. We also show that transcription activity is independent of the presence of a TATA-box-like element. METHODS Cell culture. The HPV-negative cervical carcinoma cell line C33A, the HPV-18-positive cervical cell line HeLa and the HPV-16-positive cervical cell lines SiHa and CaSki were cultured in DMEM supplemented with 10 % FCS, 1 % L-glutamine and 1 % penicillin and streptomycin at 37 uC and 5 % CO2. The day before transfection, 26105 cells were seeded in 3?5 cm 6-well Petri dishes (Nunc). Cells were then transfected using LipofectAMINE Plus (Invitrogen) and 0?5 pmol DNA per dish, following the manufacturer’s instructions. At 48 h post-transfection, cells were lysed with 300 ml Cell Culture Lysis reagent (Promega) with shaking for 90 min. Lysates were then used in a luciferase assay. Luciferin substrate (50 ml) was mixed 2910 manually with cell lysate (20 ml) and counted in a Turner 20/20 luminometer with a 3 s delay and a 12 s integration period. Each construct was tested in at least four independent experiments in triplicates using two different preparations of DNA (a minimum of 12 measurements). 5§RACE PCR and cloning. The 59RACE protocol used in these experiments is based on the CapFinder PCR cDNA principle (Chenchik et al., 1998). Total RNA was purified using TRIzol reagent (Invitrogen) and DNase treatment was performed with amplification grade DNase I (Invitrogen), according to the manufacturers’ instructions. mRNA purification was performed using the Oligotex mRNA kit (Qiagen), according to the standard procedures of the manufacturer. First-strand synthesis was performed using SuperScript II (Invitrogen). MnCl2 (2 or 3 mM) and BSA (1 mg ml21) were added to 56 first-strand buffer (Schmidt & Mueller, 1999). mRNA (100 ng) from the HPV-16-positive CaSki cell line was used as template with a poly(dT) primer and a special cap primer (rXhoICap, 59-aagcagtggtatcaacctcgagtacgc(ggg)r-39). First-strand reactions were then treated with RNase H (Boehringer Mannheim), according to the manufacturer’s instructions. PCR amplification of first-strand cDNA was performed with Pfu polymerase (Promega) using the programme: one cycle of 94 uC for 4 min, followed by five cycles of 94 uC for 30 s, 66 uC for 1 min and 72 uC for 1 min, five cycles of 94 uC for 30 s, 62 uC for 1 min and 72 uC for 1 min, and 20 cycles of 94 uC for 30 s, 58 uC for 1 min and 72 uC for 1 min. The primers used were an HPV-16-specific primer (770bHindIII, 59-catggacaagctttgtacgcacaaccgaagcgtagagtcacac-39), containing an HindIII restriction site, and the cap primer (XhoICap, 59-aagcagtggtatcaacctcgagtacgcggg-39), containing an XhoI site. cDNA was then visualized on a 1?2 % agarose gel. A 2 ml sample of the PCR mixture (100 ml) was used in a new PCR protocol using the programme: one cycle of 94 uC for 4 min, followed by 20 cycles of 94 uC for 30 s, 58 uC for 1 min and 72 uC for 1 min. The cDNA was then inserted into the pGL3-Basic vector (Promega) and sequenced. Cloning of reporter constructs. The template used was the pBR322 plasmid containing the HPV-16 genome (a gift from H. zur Hausen, DKFZ, Heidelberg). Different fragments of the HPV-16 genome were amplified by PCR using Pfu polymerase and specific primers containing restriction sites for KpnI (sense) and NheI (antisense). The coordinates of the different primers are nt 272–291, 332–352, 388–409, 448–463, 332–314, 388–370, 448–427 and 496–478. The different fragments were inserted into the promoter- and enhancerless pGL3-Basic or pGL3-Enhancer vectors (Promega), the latter contains the simian virus 40 (SV40) enhancer. pGL3-Basic constructs were all modified to reduce background from the vector, as described by Braunstein et al. (1999). Site-directed mutagenesis of the nt 272–448 fragment was performed using primers containing two point mutations (59-gcatggacgctagccttttcttcaggacacagtggcttttgacagttaatacacctacgtaacaaatca-39 and 59-gcatggacgctagccttttcttcaggacacagtggcttttgacagtgaagacacctaatt-39; mutations are shown in bold). The PCR product was amplified using Taq polymerase (Amersham Pharmacia Biotech) to avoid 39R59 exonuclease proofreading activity. All constructs were sequenced using T7 Sequenase DNA polymerase, version 2.0 (Amersham), according to the manufacturer’s instructions. Products were separated on a 6 % urea/polyacrylamide gel and exposed to a BioMax MS X-ray film (Kodak). RNase protection assay. Different probes were designed to span the HPV-16 genome from nt 53 to 632 (probe F), 277 to 526 (probe PI), 409 to 600 (probe PII) and 277 to 431 (probe PIII). The nt 53–632 fragment was amplified by PCR (59-atggacggatccgaaaccggttagtataaaagcagac-39 and 59-gtggacgaattccagtagagatcagttgtctc-39) and inserted into the pAlter-1 vector using the BamHI/EcoRI restrictions sites. The construct was then linearized with HindIII and used as template in a T7 in vitro transcription reaction. The fragments spanning the regions nt 277–526, 409–600 and 277–431 were amplified Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Thu, 03 Aug 2017 19:56:25 Journal of General Virology 84 HPV-16 and transcription by PCR using the following primers: PI, 59-gcatggacctcgagtccatatgctgtatgtgataaatg-39 and 59-gcatggacggtacctctgcaacaagacatacatcgaccg-39; PII, 59-gcatggacctcgaggtgtattaactgtcaaaagccactg-39 and 59-gcatggacggtacctaacatatattcatgcaatgtaggtg-39; PIII, 59-gcatggacctcgagtccatatgctgtatgtgataaatg-39 and 59-cgttaggtaccgtggcttttgacagttaatacacc-39. PCR products were then inserted into the Bluescript IISK+ vector using the KpnI/XhoI restriction sites. These constructs were all linearized with BamHI and also used as templates in a T7 in vitro transcription reaction. In vitro transcription was performed with MAXIscript (Ambion) in the presence of [a-32P]UTP to create labelled antisense probes. The probes were gel-purified and used in RNase protection assays (RPA III, Ambion). Probe F (50 000 c.p.m.) was mixed with 5 and 10 mg CaSki RNA. Probes PII (87 000 c.p.m.) and PI and PIII (80 000 c.p.m.) were mixed with 10 mg CaSki RNA. Prior to the assay, CaSki RNA was DNase-treated, according to the manufacturer’s instructions (Invitrogen). Unspecific RNA was degraded by digestion with RNase A/T1 (diluted 1 : 50 or 1 : 25) according to the protocol. Protected fragments were separated on a denaturing 5 % (7?5 % for PI and PIII) polyacylamide gel run at 260 V and visualized by exposing the gel to film (BioMax MS) for a minimum of 72 h. The RNA Century Marker Template set (Ambion) was used according to the manufacturer’s instructions to generate the RNA marker. Electrophoretic mobility shift assay (EMSA). The regions span- ning nt 272–448, 272–332, 332–388 and 388–448 of the HPV-16 genome were amplified by PCR and inserted into the pGL3-Basic vector using the NheI restriction site. Constructs were cut with NheI to make probes with a 59 overhang. Probes were purified using 1 % agarose gels, according to the manufacturer’s instructions (Qiagen). Probes were labelled with [a-32P]dATP using Klenow (New England Biolabs) and purified using Sephadex columns (Boehringer Mannheim). Small competitors were obtained by annealing complementary oligonucleotides (DNA technology, Århus). In each reaction, 10 mg HeLaScribe nuclear cell extract (Promega) or 15 mg SiHa nuclear cell extract, purified according to the procedure described by Dignam et al. (1983), were used together with 0?1 pmol probe ml21. Competitor DNA was added in 10-, 25- and 50-fold concentrations of the probe. Reactions were separated on a 6 % polyacrylamide gel and visualized by exposing the gel to BioMax MS film over night. RESULTS Novel transcription start sites in the HPV-16 genome To identify novel potential transcription start sites in the E6 ORF of HPV-16, 59RACE was used with mRNA isolated from the CaSki cell line. The method is based on Clontech CapFinder PCR cDNA synthesis (Chenchik et al., 1998). MnCl2 was included in first-strand cDNA synthesis in order to eliminate amplification of pre-terminated transcripts or partially degraded transcripts (Schmidt & Mueller, 1999). cDNA was amplified by PCR with an HPV-16-specific primer (770bHindIII) and the cap primer (XhoICap) (Fig. 1A). PCR products were analysed by agarose gel electrophoresis; several products corresponding to different 59 ends were observed (Fig. 1B). cDNAs were inserted into the pGL3-Basic vector using the HindIII/XhoI restriction sites. Colonies were then sequenced. The 59 ends for the main promoter P97 were mapped to nt 95 (data not shown) and 97 (Fig. 1C). Two 59 ends were mapped to nt 483 and 489. Promoter P542, identified previously, was confirmed http://vir.sgmjournals.org also. In addition, three 59 ends were mapped to nt 430, 441 and 446. To verify the novel 59 ends, RNase protection studies were performed with total RNA extracted from CaSki cells. A long antisense probe spanning the region of nt 53–632 (probe F) of the HPV-16 genome was hybridized with different amounts of CaSki RNA. After hybridization, unprotected RNA was degraded by digestion with RNase A/T1 and protected fragments were separated in a polyacrylamide gel. Protected fragments corresponding to transcripts originating from the main promoter (P97) with the known splice sites at nt 226^409 and 226^526 (Doorbar et al., 1990) were detected (Fig. 2A, B). In addition, two protected fragments of approximately 150 and 185 bp were observed. The experiment was performed several times with different RNA batches of CaSki and W12 cells (data not shown); these fragments were observed repeatedly (Fig. 2A, B). These two fragments could correspond to transcripts initiating around nt 441 and 480, respectively. In addition, a fragment of approximately 260 bp was observed repeatedly; the identity of this fragment is still unknown. To verify that the fragments observed were specific and originated from the 39 end of probe F, we repeated the RNase protection assay, this time using three small overlapping probes spanning the regions of nt 277–526 (PI), 409–600 (PII) and 277–431 (PIII) (Fig. 2C–E). Three protected fragments, which could correspond to transcripts originating from P97, nt 441 and 430, were observed using either PI or PII, but not PIII (Fig. 2C–E). The experiment performed with PIII verifies that the three fragments observed with probes F, PI and PII originate from the 39 end of the probes. Also, a protected fragment corresponding to a transcript beginning at nt 480 was observed using PII (Fig. 2D). In the experiment performed with PI, a protected fragment with the length of approximately 174 bp was observed. This fragment was not identified in the experiments using PII, but could correspond to the unidentified fragment observed using probe F (Fig. 2B). The experiments were repeated several times and the same fragments appear using the two different probes (F and PI); therefore, it must be a specific, protected fragment. The origin of this fragment is still unknown. In conclusion, the four different probes reproducibly protect fragments corresponding to the P97 and P480 mRNAs described previously, but also fragments corresponding to 59 ends around nt 441 and 430. Therefore, these experiments support the presence of transcripts initiating between nt 430 and 441. Promoter activity of luciferase reporter constructs To examine the promoter activity associated with the novel transcription start sites, different HPV-16 fragments were amplified by PCR and inserted into the pGL3-Enhancer vector, which contains the SV40 enhancer and the luciferase reporter gene. The SV40 enhancer was used as a substitute Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Thu, 03 Aug 2017 19:56:25 2911 M. W. Rosenstierne and others Fig. 1. (A) PCR amplification of cDNA using the special cap primer (rXhoICap) and the HPV-16-specific primer (770bHindIII). The transcripts expected are summarized. (B) PCR products were visualized on a 1?2 % agarose gel. The outer lanes show the pBR322-HinfI marker. Two different PCR reactions are shown in duplicate. The different products corresponding to different transcripts are shown. (C) PCR fragments were purified and inserted into the pGL3-Basic vector. Different clones were sequenced using a vector-specific primer. The first sequence shows a transcript beginning at nt 97 (the main promoter P97). The arrow points to the first nucleotide in the HPV-16 sequence. Sequences of the different transcription start sites identified in the E6 ORF of HPV-16 are shown, and the dotted lines separate the DNA sequences originating from the pGL3-Basic vector, the XhoI restriction site, the CapFinder sequence and the HPV-16 sequence. for the HPV enhancer (LCR), which, in the natural context of the full-length HPV genome, will influence the activity of different HPV promoters. Constructs were designed to include the novel transcription start sites at nt 430, 441 and 446 either alone (nt 272–448) or together with the transcription start site identified previously at nt 480 (nt 272–496) (Fig. 3A). The different pGL3-Enhancer constructs were transiently transfected into HPV-16-positive SiHa cells, HPV-18-positive HeLa cells and HPV-negative C33A cells. Luciferase activity was measured 48 h after 2912 transfection (Fig. 3B). Each construct was tested twice in triplicates with two different batches of DNA, resulting in at least 12 measurements for each construct. The 272–448E construct had a significantly higher luciferase activity than the empty pGL3-Enhancer vector in all three cell lines tested, with the highest activity in the SiHa cell line (8?5-fold increase). The 272–496E construct had lower activity than the 272–448E construct in SiHa and HeLa cells but in the C33A cell line both fragments showed similar activity (2fold increase). A short construct (448–496E) that contained Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Thu, 03 Aug 2017 19:56:25 Journal of General Virology 84 HPV-16 and transcription only the transcription start site at nt 480 had no activity. These results demonstrate that the region at nt 272– 448 is needed to activate transcription from nt 480. To identify the essential regions responsible for promoter activity, upstream and downstream deletions were made (Fig. 3A). Deletion of the region containing the transcription start sites reduced luciferase activity to that of the empty pGL3-Enhancer vector. Deletions of the upstream region also showed a luciferase activity corresponding to that of the empty vector, except for the 332–496E construct in the SiHa cell line, which showed a 1?5-fold increase in activity; this, however, is a 5-fold reduction compared to that seen with construct 272–496E (Fig. 3B). These results indicate that the region from nt 272 to 332 is essential for transcription initiation from the two clusters of transcription start sites around nt 441 and 480. proteins bind specifically to this region. No specific binding of proteins from the SiHa nuclear cell extract to the intermediate region from nt 332 to 388 was observed. In contrast, the HeLa nuclear cell extract (Fig. 4D, lane 4) showed binding to the probe specific for nt 332–388, which could be competed by the nt 332–354 competitor fragment. However, a 50-fold excess of competitor could not remove the shifted band completely and the specificity of this binding therefore remains unclear. The region from nt 388 to 448 showed four shifted bands, with both the SiHa and the HeLa nuclear cell extracts (Fig. 4F, lane 7). The binding region was within nt 388–411. These results demonstrate that proteins from SiHa and HeLa nuclear cell extracts bind specifically to the region upstream of the nt 441 transcription start sites. To verify that the promoter activity observed was not due to unspecific initiation of transcription caused by the strong SV40 enhancer, different fragments were also inserted into the pGL3-Basic vector, which lacks the SV40 enhancer. The pGL3-Basic vector was modified further to reduce background activity, as described by Braunstein et al. (1999). In these experiments, the HPV-16 fragments contained only the transcription start sites around nt 441 (Fig. 3C). The same upstream and downstream deletions as mentioned above were constructed. Transient transfections were performed using the SiHa cell line because this cell line showed the highest luciferase activity (Fig. 3B). Construct m272– 448B showed considerably higher activity (103-fold) than the empty pGL3-Basic vector. Deletions of the region containing the transcription start sites all reduced luciferase activity to that of the empty vector. Again, an approximately 5-fold reduction was seen for construct m332–448B. In conclusion, these results confirm that promoter activity is present within the region of nt 272–448 and that this activity is not an artefact introduced by the strong SV40 enhancer. Furthermore, the experiments confirmed that the region at nt 272–332 is essential for activation of transcription from the novel start sites around nt 441. Transcription is independent of a TATA-box-like sequence Binding of proteins to the region at nt 272–448 To investigate binding of transcription factors to the region of nt 272–448, EMSA analysis was performed with SiHa and HeLa nuclear cell extracts. DNA probes spanning the regions at nt 272–332, 332–388 and 388–448 were 59 end-labelled with [a-32P]ATP. All three probes were shifted by both nuclear extracts (Fig. 4B, D and F, lanes N). Competition experiments were performed with full-length, non-labelled probes (data not shown) or smaller (approximately 23 bp) overlapping DNA probes, as illustrated in Fig. 4(A, C and E). The binding of HeLa nuclear cell extract to the probe specific for nt 272–332 was competed by the nt 291–314 competitor fragment (Fig. 4B, lane 2). This indicates that a protein binds specifically in this region. The experiments performed with SiHa nuclear cell extract showed competition of two of the shifted bands within the same region (Fig. 4B, lane 2), again indicating that http://vir.sgmjournals.org Two TATA-box-like elements are present upstream of the transcription start sites around nt 441. One element is positioned at nt 401–407 with the sequence 59-TTAATTA-39 (Fig. 5A, mut I), which is in the protein-binding region of nt 388–411 identified by EMSA (Fig. 4F, lane 7). The second element is situated immediately adjacent to the protein-binding region at nt 412–417 with the sequence 59-TATTAA-39 (Fig. 5A, mut II). Point mutations were introduced into both elements to investigate influences on the activity of the start sites around nt 441. Fragments amplified by PCR were cloned into the pGL3-Enhancer vector, sequenced and transfected into the C33A, HeLa and SiHa cell lines. Luciferase activity was measured subsequently. No significant difference in the promoter activity between the wild-type and mutant constructs was observed (Fig. 5B). These results show that transcription from the cluster of transcription start sites around nt 441 functions independently of the TATA-box-like element. DISCUSSION The data presented here identify a novel cluster of transcription start sites around nt 441 in the E6 ORF of the HPV16 genome. Initiation sites were mapped with a 59RACE PCR method. In addition to the new initiation sites, we mapped transcripts from P97 to initiate at nt 97 and 95. We also found a cluster of transcription start sites present at around nt 480, which have been described previously as the promoter P480 (Grassmann et al., 1996; Braunstein et al., 1999). To verify the presence of transcription start sites around nt 441, we performed RNase protection assays with four different probes. The experiment performed with PIII was designed to verify that all protected fragments observed using the probes F, PI and PII originated from the 39 end of the probes. Probe F, PI and PII repeatedly identified protected fragments corresponding to transcripts from P97, including the different splice patterns. In addition, fragments that correspond to transcripts initiated from P480, nt 441 and 430 were identified with the same three probes and Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Thu, 03 Aug 2017 19:56:25 2913 M. W. Rosenstierne and others thus show that HPV-16 transcripts initiating around nt 441 are present in the CaSki cell line. Polymerase II promoters with several start sites are usually not dependent on a TATA-box element (Ince & Scotto, 1995). Accordingly, we find that point mutations made in the two TATA-box-like elements present upstream of the multiple transcription start sites did not influence promoter activity. The data, therefore, support the conclusion that transcription initiation from the several novel transcription start sites is independent of a TATA-box element. Comparison of the sequence surrounding the transcription start sites did not reveal any core elements or initiator sequences (for reviews, see Butler & Kadonaga, 2002; and Ince & Scotto, 1995). Promoters with multiple transcription start sites and no recognizable core elements have also been identified in other regions of the genomes from a number of different HPV types. They include the late differentiationdependent promoters (DiLorenzo & Steinberg, 1995; Grassmann et al., 1996; Karlen et al., 1996; Klumpp & 2914 Laimins, 1999; Nasseri et al., 1987; Ozbun & Meyers, 1997, 1998; del Mar Peña & Laimins, 2001; Tomita et al., 1996). Some of these transcription start sites are scattered over relatively large regions of the genome; for example, P670 (HPV-16) has transcription start sites spanning approximately 100 nt from nt 667 to 766 (Grassmann et al., 1996). In HPV-31b, 30 transcription start sites have been mapped to a region spanning approximately 200 nt in the E7 ORF (del Mar Peña & Laimins, 2001). Considering these findings, it seems likely that the transcription start sites clustered around nt 441 and P480 are part of a large cluster of transcription initiation sites and are controlled by the same sequence elements. Our finding supports this view, since the promoter activity of the fragment at nt 448–496 does not show any promoter activity unless the upstream region (nt 272–448) is included. Furthermore, our finding suggests that the upstream region at nt 272–332 is required for transcription activity. Possibly, this region recruits the general transcription machinery or parts thereof and, in the absence of consensus core promoter signals, initiates Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Thu, 03 Aug 2017 19:56:25 Journal of General Virology 84 HPV-16 and transcription Fig. 2. RNase protection assay. (A) Locations of the different promoters of the HPV-16 genome. The HPV-16-specific probes F (nt 53–632), PI (nt 277–526), PII (nt 409–600) and PIII (nt 277–431) and their expected fragments originating from the different splice patterns of transcripts and initiation sites are shown. Crossed boxes indicate part of the linker region from the linearized vector, 30 nt from the pAlter-1 vector on probe F and 60 nt from the Bluescript IISK+ vector on probes PI, PII and PIII. (B) RNase protection gel using probe F. Reactions using 5–10 mg CaSki RNA are shown in lanes F. Fragments corresponding to the different specific HPV transcripts are shown. RNase protection gel using PI (C), PII (D) and PIII (E). Fragments corresponding to the different specific HPV transcripts are shown. Lanes: M, RNA marker (bp); U, 10 % of the probe used in the experiment without CaSki RNA; C, control for the specificity of the probe using 20 mg yeast RNA. transcription from several positions within the region of nt 388–496. Many of the late promoters with multiple transcription start sites that have been identified in the different HPV types are dependent on differentiation. In HPV-31, only some of the multiple transcription start sites clustered around the P742 promoter are active in non-differentiated cells, but upon differentiation a number of these transcription start sites are activated (Ozbun & Meyers, 1998; del Mar Peña & Laimins, 2001). It has also been shown that the E7 promoter of HPV6 is upregulated in differentiated cells (Ai et al., 1999). One http://vir.sgmjournals.org possible mechanism for this upregulation is differentiationdependent downregulation of the repressor protein (CDP/ cut). This protein is present in non-differentiated cells and binds to AT-rich MAR (nuclear matrix attachment) elements in the promoter region. A similar MAR element is present in the E6 ORF of HPV-16 (Tan et al., 1998) and this region also binds the CDP/cut repressor (Stünkel et al., 2000). Our data confirm protein binding to this region. By EMSA analysis, we found four band shifts within the region of nt 388–411 that could be specifically competed. Identification of the binding proteins is currently under investigation. A strong candidate is the CDP/cut repressor, Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Thu, 03 Aug 2017 19:56:25 2915 M. W. Rosenstierne and others Fig. 3. Luciferase reporter assay. (A) The two clusters of transcription start sites around nt 441 and 480 are shown together with the different DNA fragments inserted into the pGL3-Enhancer vector. The different coordinates are given. (B) All fragments inserted into the pGL3-Enhancer vector are indicated by the suffix E. Luciferase activity is given as a percentage of the empty pGL3-Enhancer vector. Two different batches of DNA were used for each construct and tested twice in triplicates with a total of 12 measurements. Bars indicate SD. (C) The different DNA fragments inserted into the pGL3-Basic vector are shown together with the coordinates. (D) The pGL3-Basic constructs were all modified to reduce background activity and are therefore given the prefix m in addition to the suffix B. Luciferase activity is given as a percentage of the empty modified pGL3Basic vector. Two different batches of DNA were used for each construct and were tested twice in triplicates with a total of 12 measurements. Bars indicate SD. since Stünkel et al. (2000) have identified a strong binding of CDP/cut to an oligonucleotide spanning the region of nt 374–403. It would be interesting to study the influence of 2916 cell differentiation on the transcription activity of the cluster of transcription start sites around nt 441 and we plan to address this in future studies. Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Thu, 03 Aug 2017 19:56:25 Journal of General Virology 84 HPV-16 and transcription Fig. 4. Binding of protein to the promoter region. (A, C and E) The different probes and competitors with coordinates are shown. Numbers 1–9 designate the small competitors. (B, D and F) EMSA gels using HeLa and SiHa nuclear cell extracts. Reactions with nuclear extract but without competitor are seen in lanes N. The different competitors added in three increasing concentrations are shown for each gel. Arrows point towards the specific fragments that respond to competition. http://vir.sgmjournals.org Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Thu, 03 Aug 2017 19:56:25 2917 M. W. Rosenstierne and others Fig. 5. Site-directed mutagenesis. (A) The sequence of nt 388–448 containing the cluster of transcription start sites around nt 441 is shown. The protein-binding region is marked with a grey background. TATA-box-like elements are shown in bold and mutated nucleotides are underlined. Adenine was changed to cytosine and thymidine was changed to guanine. (B) Luciferase activity in the three cell lines is given as a percentage of the empty pGL3-Enhancer vector. Two different batches of DNA were used for each construct and tested twice in triplicates with a total of 12 measurements. Bars indicated SD. Previously, Stünkel et al. (2000) have shown that the E6 MAR element of HPV-16 is a strong cis-responsive element when situated downstream of P97 in a reporter construct. In transient transfection studies, the E6 MAR element repressed the promoter activity of P97. However, in stable transfections with the constructs integrated into the host genome, the promoter activity of P97 was enhanced. Deletion constructs showed that the region at nt 246–356 had to be included in the constructs in order to activate P97 when integrated in the host genome. We found that the region at nt 272–332 is essential for the promoter activity of nt 441. Our results, together with the data presented by Stünkel et al. (2000), indicate therefore that the region at nt 272–332 is a strong cis-responsive element, which, in the natural context of the genome, might work on both the main promoter P97 and the cluster of transcription start sites around nt 441. In summary, our data show that there is a new cluster of transcription start sites around nt 441 in the E6 ORF of 2918 HPV-16. The transcription start site clusters around nt 441 and 480 and the P542 promoter identified previously all produce messengers with potential to express E7. Although the P441, P480 and P542 messengers are expressed at low levels compared with P97 messengers, they may contribute considerably to the overall expression of E7 through more efficient translation. Therefore, the P441, P480 and P542 promoters could be potentially important for malignant progression of HPV-16-infected cells. ACKNOWLEDGEMENTS We acknowledge Harald zur Hausen (DKFZ, Heidelberg) for the pBR322 plasmid containing the HPV-16 genome. We thank Med Jørgen Olsen (Biochemistry Department C, Panum Institute) for advice and technical guidance on the preparation of SiHa nuclear cell extracts and EMSA analysis. This work was supported by grants from the Danish Cancer Institute, the Danish Medical Research Council, the Biotechnology Foundation, the Novo Nordisk Foundation, the Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Thu, 03 Aug 2017 19:56:25 Journal of General Virology 84 HPV-16 and transcription Glud Foundation, the Arvid Nilsson Foundation, the Wedell Wedellsborg Foundation, the Klestrup Foundation, the Direktør E. Danielsen Foundation and the Dagmar Marshall Foundation. Nishimura, A., Ono, T., Ishimoto, A., Dowhanick, J. J., Frizzell, M. A., Howley, P. M. & Sakai, H. (2000). Mechanisms of human papil- lomavirus E2-mediated repression of viral oncogene expression and cervical cancer cell growth inhibition. J Virol 74, 3752–3760. O’Connor, M. J., Chan, S. Y. & Bernard, H.-U. (1995). Transcription REFERENCES factor binding sites in the long control regions of genital HPVs. In Human Papillomaviruses 1995 Compendium, vol. IIIA, pp. 21–40. Los Alamos, NM: Los Alamos National Laboratory. Ai, W., Toussaint, E. & Roman, A. (1999). CCAAT displacement protein binds to and negatively regulates human papillomavirus type 6 E6, E7, and E1 promoters. J Virol 73, 4220–4229. O’Connor, M. J., Stünkel, W., Zimmermann, H., Koh, C.-H. & Bernard, H.-U. (1998). A novel YY1-independent silencer represses Braunstein, T. H., Madsen, B. S., Gavnholt, B., Rosenstierne, M. W., Koefeld Johnsen, C. & Norrild, B. (1999). Identification of a new promoter in the early region of the human papillomavirus type 16 genome. J Gen Virol 80, 3241–3250. Butler, J. E. F. & Kadonaga, J. T. (2002). The RNA polymerase II core promoter: a key component in the regulation of gene expression. Genes Dev 16, 2583–2592. Chenchik, A., Zhu, Y. Y., Diatchenko, L., Li, R., Hill, J. & Siebert, P. D. (1998). Generation and use of high-quality cDNA from small amounts of total RNA by SMARTTM PCR. In Gene Cloning and Analysis by RT-PCR. Biotechniques Books: Eaton. Chow, L. T., Nasseri, M., Wolinsky, S. M. & Broker, T. R. (1987). Human papillomavirus type 6 and 11 mRNAs from genital condylomata acuminata. J Virol 61, 2581–2588. del Mar Peña, L. M. & Laimins, L. A. (2001). Differentiation- dependent chromatin rearrangement coincides with activation of human papillomavirus type 31 late gene expression. J Virol 75, 10005–10013. Dignam, J. D., Lebovitz, R. M. & Roeder, R. G. (1983). Accurate transcription initiation by RNA polymerase II in a soluble extract from isolated mammalian nuclei. Nucleic Acids Res 11, 1475–1489. DiLorenzo, T. P. & Steinberg, B. M. (1995). Differential regulation of human papillomavirus type 6 and 11 early promoters in cultured cells derived form laryngeal papillomas. J Virol 69, 6865–6872. Doorbar, J., Parton, A., Hartley, K., Banks, L., Crook, T., Stanley, M. & Crawford, L. (1990). Detection of novel splicing patterns in an HPV16-containing keratinocyte cell line. Virology 178, 254–262. Doorbar, J., Elston, R. C., Napthine, S. & 9 other authors (2000). The E1E4 protein of human papillomavirus type 16 associates with a putative RNA helicase through sequences in its C terminus. J Virol 74, 10081–10095. Glahder, J. A., Hansen, C. N., Vinther, J., Madsen, B. S. & Norrild, B. (2003). A promoter within the E6 ORF of human papillomavirus type 16 contributes to the expression of the E7 oncoprotein from a monocistronic mRNA. J Gen Virol (in press). Grassmann, K., Rapp, B., Maschek, H., Petry, K. U. & Iftner, T. (1996). Identification of a differentiation-inducible promoter in the E7 open reading frame of human papillomavirus type 16 (HPV-16) in raft cultures of a new cell line containing high copy numbers of episomal HPV-16 DNA. J Virol 70, 2339–2349. the activity of the human papillomavirus type 16 enhancer. J Virol 72, 10083–10092. O’Connor, M. J., Stünkel, W., Koh, C.-H., Zimmermann, H. & Bernard, H.-U. (2000). The differentiation-specific factor CDP/Cut represses transcription and replication of human papillomaviruses through a conserved silencing element. J Virol 74, 401–410. Ozbun, M. A. & Meyers, C. (1997). Characterization of late gene transcripts expressed during vegetative replication of human papillomavirus type 31b. J Virol 71, 5161–5172. Ozbun, M. A. & Meyers, C. (1998). Temporal usage of multiple promoters during the life cycle of human papillomavirus type 31b. J Virol 72, 2715–2722. Phelps, W. C., Barnes, J. A. & Lobe, D. C. (1998). Molecular targets for human papillomaviruses: prospects for antiviral therapy. Antivir Chem Chemother 9, 359–377. Schmidt, W. & Mueller, M. W. (1999). CapSelect: a highly sensitive method for 59 CAP-dependent enrichment of full-length cDNA in PCR-mediated analysis of mRNA. Nucleic Acids Res 27, e31. Schneider-Gädicke, A. & Schwarz, E. (1986). Different human cervical carcinoma cell lines show similar transcription patterns of human papillomavirus type 18 early genes. EMBO J 5, 2285–2292. Seedorf, K., Krämmer, G., Dürst, M., Suhai, S. & Röwekamp, W. G. (1985). Human papilloma virus type 16 DNA sequence. Virology 145, 181–185. Smotkin, D., Prokoph, H. & Wettstein, F. O. (1989). Oncogenic and nononcogenic human genital papillomaviruses generate the E7 mRNA by different mechanisms. J Virol 63, 1441–1447. Stacey, S. N., Jordan, D., Snijders, P. F. J., Mackett, M., Walboomers, J. M. M. & Arrand, J. R. (1995). Translation of the human papillomavirus type 16 E7 oncoprotein from bicistronic mRNA is independent of splicing events within the E6 open reading frame. J Virol 69, 7023–7031. Stünkel, W. & Bernard, H.-U. (1999). The chromatin structure of the long control region of human papillomavirus type 16 represses viral oncoprotein expression. J Virol 73, 1918–1930. Stünkel, W., Huang, Z., Tan, S.-H., O’Connor, M. J. & Bernard, H.-U. (2000). Nuclear matrix attachment regions of human papillomavirus type 16 repress or activate the E6 promoter, depending on the physical state of the viral DNA. J Virol 74, 2489–2501. Tan, S.-H., Leong, L. E. C., Walker, P. A. & Bernard, H.-U. (1994). defines a new class of RNA polymerase II promoters. J Biol Chem 270, 30249–30252. The human papillomavirus type 16 E2 transcription factor binds with low cooperativity to two flanking sites and represses the E6 promoter through displacement of Sp1 and TFIID. J Virol 68, 6411–6420. Karlen, S., Offord, E. A. & Beard, P. (1996). Functional promoters in Tan, S.-H., Bartsch, D., Schwarz, E. & Bernard, H.-U. (1998). Ince, T. A. & Scotto, K. W. (1995). A conserved downstream element the genome of human papillomavirus type 6b. J Gen Virol 77, 11–16. Klumpp, D. J. & Laimins, L. A. (1999). Differentiation-induced changes in promoter usage for transcripts encoding the human papillomavirus type 31 replication protein E1. Virology 257, 239–246. Nasseri, M., Hirochika, R., Broker, T. R. & Chow, L. T. (1987). A human papilloma virus type 11 transcript encoding an E1^E4 protein. Virology 159, 433–439. http://vir.sgmjournals.org Nuclear matrix attachment regions of human papillomavirus type 16 point toward conservation of these genomic elements in all genital papillomaviruses. J Virol 72, 3610–3622. Thierry, F., Heard, J. M., Dartmann, K. & Yaniv, M. (1987). Charac- terization of a transcriptional promoter of human papillomavirus 18 and modulation of its expression by simian virus 40 and adenovirus early antigenes. J Virol 61, 134–142. Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Thu, 03 Aug 2017 19:56:25 2919 M. W. Rosenstierne and others Tomita, Y., Shiga, T. & Simizu, B. (1996). Characterization of a promoter in the E7 open reading frame of human papillomavirus type 11. Virology 225, 267–273. Ushikai, M., Lace, M. J., Yamakawa, Y. & 9 other authors (1994). Trans activation by the full-length E2 proteins of human papillomavirus type 16 and bovine papillomavirus type 1 in vitro and in vivo: cooperation with activation domains of cellular transcription factors. J Virol 68, 6655–6666. Zhao, W., Chow, L. T. & Broker, T. R. (1997). Transcription activities of human papillomavirus type 11 E6 promoter-proximal elements in 2920 raft and submerged cultures of foreskin keratinocytes. J Virol 71, 8832–8840. zur Hausen, H. (1996). Papillomavirus infections: a major cause of human cancers. Biochim Biophys Acta 1288, F55–F78. zur Hausen, H. (1999). Papillomaviruses in Human Cancers, vol. 111, pp. 581–587. Proceedings of the Association of American Physicians. Zwerschke, W. & Jansen-Dürr, P. (2000). Cell transformation by the E7 oncoprotein of human papillomavirus type 16: interactions with nuclear and cytoplasmic target proteins. Adv Cancer Res 78, 1–29. Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Thu, 03 Aug 2017 19:56:25 Journal of General Virology 84