Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
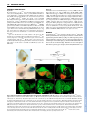
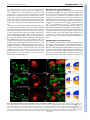
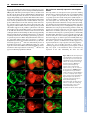
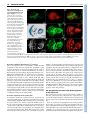
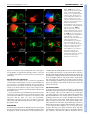
RESEARCH ARTICLE 659 Development 134, 659-667 (2007) doi:10.1242/dev.02786 Dorsal-ventral midline signaling in the developing Drosophila eye Atsushi Sato and Andrew Tomlinson* Boundaries between different cell types play key roles in many developmental patterning processes. They can be established by various mechanisms, and signaling between the different cell types can occur in a number of ways. One mechanism of crossboundary signaling is controlled by the Notch (N)-modifying protein Fringe (Fng). At the Drosophila wing dorsal-ventral (D-V) border, the mechanism by which an Fng+-Fng– interface controls local N activation has been well characterized. A similar Nactivating Fng+-Fng– interface has also been described at the D-V border of the fly eye, but the mechanisms that establish and regulate it are different from those in the wing. Here we describe the ventral role of the Sloppy-paired (Slp) transcription factor, and its interactions with dorsally expressed Iroquois (Iro) transcription factors in the regulation of signaling about the Fng+-Fng– interface in the developing eye. The two transcription factors are mutually repressive and initially abut at the D-V midline. However, N signaling at the interface downregulates Slp expression, and a gap opens between the two expression domains in which Serrate (Ser, an N ligand) is upregulated. INTRODUCTION Development is a stepwise process in which new information is generated through the interactions of previously specified cell types. The place where different cell types abut often defines the position from which secreted molecules are released, and these signals then direct further elaboration of patterning (Lawrence and Struhl, 1996). The cell types at the interface not only define the position from which the signal will be released, but they also carry the molecular information that directs which type of signal will be expressed, and the modes of response of the cells. Interfaces between different cell types can be established by a number of mechanisms. In the fly wing, the dorsal-ventral (D-V) boundary is established by a compartment mechanism, whereby cells on either side of the border are clonally isolated from one another: dorsal cells express the Apterous (Ap) transcription factor and share a lineage ancestry isolated from the ventral cells that do not express Ap (Blair et al., 1994; Diaz-Benjumea and Cohen, 1993). Differential adhesion between the two cell types leads to the formation of a long interface (the D-V border). Borders between distinct cell types can be generated by non-compartment mechanisms. For example, in the vertebrate neural tube, Sonic Hedgehog released from the ventral floorplate diffuses dorsally to elicit the expression of an array of different transcription factors, each expression domain depending on its distance from the source (Jessell, 2000). This first step establishes a relatively crude array of expression domains with many cells expressing more than one transcription factor. The transcription factors, however, are autoregulatory and mutually repressive, and if a cell initially expresses two, the action of one will force the extinction of the other. As with the compartment mechanism, the transcription factors also provide a miscibility code, such that likecells mix and unlike-cells repel, and sharply defined interfaces are formed. Department of Genetics and Development, College of Physicians and Surgeons, Columbia University, 701 West 168th St, HHSC 1120, New York, NY 10032, USA. *Author for correspondence (e-mail: [email protected]) Accepted 14 December 2006 There is great variability in the types of signaling that can occur at interfaces. A particularly important form is that regulated by Fringe (Fng), a glycocyltransferase that modifies the receptor Notch (N) and thereby modulates its ligand sensitivities (Bruckner et al., 2000; Moloney et al., 2000; Munro and Freeman, 2000). The N/Fng-mediated signaling was first described at the D-V border of the fly wing (Klein and Arias, 1998; Panin et al., 1997) and has since been described elsewhere in fly and vertebrate patterning (Haines and Irvine, 2003; Irvine, 1999). Once a Fng+Fng– border is established, a dynamic interaction between N and its ligands leads to high-level ligand expression and consequential upregulation of N activity. A Fng+-Fng– boundary also forms at the D-V midline of the fly eye and the resulting N signaling activates a number of genes needed for growth and patterning of the eye (Cho and Choi, 1998; Dominguez and de Celis, 1998; Papayannopoulos et al., 1998). The D-V midlines of the eye and wing display a number of major differences. First, the D-V domains of the eye do not appear to be generated by a compartment mechanism as clones can violate the DV midline (Becker, 1957). Second, in the wing, Fng is expressed dorsally (under Ap transcriptional control), but in the eye it is expressed ventrally [repressed dorsally by Iroquois (Iro) transcription factors] (Cho and Choi, 1998; Dominguez and de Celis, 1998; Irvine and Wieschaus, 1994; Papayannopoulos et al., 1998). Thus, the eye and wing both use the Fng/N signaling mechanism at the D-V midlines, but the mechanisms by which the interfaces are established are different. Sloppy-paired 1 and 2 (Slp1 and Slp2) are homologous Forkhead transcription factors that have extensive and frequently redundant roles in embryonic patterning (Grossniklaus et al., 1992). We describe here the ventral-specific expression of Slp proteins in the developing eye. We show that Slp and Iro proteins are mutually repressive transcription factors expressed in the ventral and dorsal domains of the developing eye, respectively. Initially, the two domains directly abut at the center of the eye disc, but as development proceeds a gap opens up between them. This gap is caused by the N-induced downregulation of slp, which facilitates the upregulation of Serrate, one of the N ligands. DEVELOPMENT KEY WORDS: Drosophila, Eye, Sloppy-paired, Dorsoventral signaling RESEARCH ARTICLE Development 134 (4) MATERIALS AND METHODS Histology Fly stocks Antibodies used were rat anti-Mirr (Yang et al., 1999) (1:2000), rat anti-Ser (Papayannopoulos et al., 1998) (1:1000), mouse anti-Dl (DSHB, 1:600), rabbit anti-LacZ (Cappel, 1:2000), mouse anti--galactosidase (Promega, 1:500), anti-rabbit IgG Alexa Fluor 488-conjugated (Molecular Probes, 1:1000), anti-mouse IgG Alexa Fluor 488-conjugated (Molecular Probes, 1:1000), anti-rabbit IgG Cy3-conjugated (Jackson, 1:1000), anti-mouse IgG Cy3-conjugated (Jackson, 1:1000), anti-rat IgG Cy3-conjugated (Jackson, 1:1000), anti-mouse IgG Cy5-conjugated (Jackson, 1:500) and antidigoxigenin alkaline phosphatase-conjugated (Roche, 1:2000). Standard methods were used for antibody staining. slp1, slp2 (Grossniklaus et al., 1992), mirr (McNeill et al., 1997) and fng (Irvine and Wieschaus, 1994) cDNAs were used for in situ hybridization following Curtiss and Mlodzik (Curtiss and Mlodzik, 2000) and/or Sato et al. (Sato et al., 1999). Fly strains used are: yw, slp105965 (slp1-lacZ; Bloomington Stock Center), mirrF7 (mirr-lacZ) (McNeill et al., 1997), eyg-Gal4 (Wehrli and Tomlinson, 1998), Tub>Gal80, y+>Gal4 (gift from Gary Struhl), UAS-slp1 (gift from Gary Struhl), UAS-slp2 (Riechmann et al., 1997), UAS-mirr (Cavodeassi et al., 2000), UAS-Nintra (Struhl and Adachi, 1998), UAS-GFP (Bloomington Stock Center), iroDFM3 (Gomez-Skarmeta et al., 1996), UASP35 (Hay et al., 1994), Nts (Shellenbarger and Mohler, 1978) and hs-N-GV (Chung and Struhl, 2001). Clones were made using slpS37A FRT40A, ubiGFP, FRT40A (Bloomington Stock Center), hs-GFP M(2) w+30C FRT40A (gift from Gary Struhl), iroDFM3 FRT2A (Tomlinson, 2003) and hs-GFP M(3)i55 FRT2A chromosomes (gift from Gary Struhl). We also made slpS37A mutant clones with UAS-P35 using MARCM methods (Lee and Luo, 1999). Clones were induced at 48 or 72 hours before dissection. Nts flies were grown at 18°C and shifted to 31°C and left for 24 hours before dissection. slpAVm is an insertion line of a UAS construct at the slp2 locus. The insertion site was determined by inverse PCR and sequencing. slpS37A was made by imprecise P-element excision from slpAVm, and screened by complementation with slp105965. Break points of the deletion were determined by PCR and sequencing. Part of the P-element was not deleted. This sequence of this lesion is available at GenBank (Accession No. EF141189). RESULTS An insertion (slpAvm) of a construct carrying a mini-w+ selectable marker (transformed for other purposes) showed a ventral-anterior pigmentation pattern in adult eyes (Fig. 1A). This expression pattern suggested that the ‘trapped’ gene was expressed exclusively in the ventral region of the developing eye. Inverse PCR identified the slp locus as the site of insertion (Fig. 1B). There are two transcription units in the slp locus (slp1 and slp2) and in situ hybridization to eye Fig. 1. Expression patterns, mutant phenotypes and genomic organization of slp. (A) Adult eye of the slp2 insertion line (slp2AVm) shows pigmentation in the anterior/ventral domain of the eye. (B) Genomic map of the slp locus showing the site of insertion of slp2AVm and the deletion induced (slpS37A, dotted lines). (C) slp-lacZ expression in the second-instar eye disc. (D) mirr-lacZ expression in the second instar. (E) Double staining of slp-lacZ (red) and Mirr (green) of second-instar eye disc. The two domains abut in the center of the disc. (F) Double staining of slp-lacZ (red) and Mirr (green) of third-instar head disc. The white arrowhead indicates the gap between the slp and Mirr expression domains. Arrows indicate slp expression in the dorsal regions of the extreme anterior of the eye disc, and in the posterior dorsal (relative to the eye) regions of the antennal disc. (G-I) Survival of slpS37A mutant clones (black) in third-instar eye discs, induced at different times of development. Yellow arrowhead indicates morphogenetic furrow (MF). (G) When clones are induced 0-24 hours AEL, twin spots (arrow) are readily evident in the ventral regions, but mutant clones are rarely observed. (H) Clones induced 24-48 hours AEL do not survive ventrally ahead of the MF (arrow points to a twin spot), but can be observed behind (white arrowheads), but are rounded with smooth borders. (I) Clones induced 48-72 hours AEL. slpS37A mutant clones (white arrowheads) survive ahead of the MF but show smooth borders. In all images, dorsal is up and posterior is left. DEVELOPMENT 660 discs showed that the two were co-expressed (not shown) in the same pattern as the slp1-lacZ line (slp105965). slp-lacZ thus appears to be a faithful reporter of transcription of the slp locus in the developing eye. slp-lacZ expression was observed in the ventral domain of the second-instar eye disc, extending into the posterior portion of the antennal disc (Fig. 1E), and in the anterior/ventral regions of late-third-instar discs (Fig. 1F), and also in dorsal regions of the extreme anterior of the eye disc and in the posterior dorsal (relative to the eye disc) regions of the antennal disc (arrows in Fig. 1F). Eye discs were double-stained for transcriptional activity of the iro complex (dorsally expressed) and the slp complex (ventrally expressed) to determine the spatial relationship of the two expression domains. In second-instar eye discs, mirror (mirr, one of the three iro genes) transcription was exclusively dorsal (Fig. 1D,E) whereas slp was exclusively ventral (Fig. 1C,E), and the two expression domains directly abutted (with no overlap) in the medial region of the eye disc (Fig. 1E). In the third larval instar the morphogenetic furrow (MF) begins to progress across the presumptive retina, and slp and mirr transcription was extinguished in post-furrow regions as evidenced by in situ hybridization (not shown). However, lacZ reporters from both genes showed persistent expression posterior to the MF, which is likely to result from perdurance of the long-lived lacZ gene product (Fig. 1F). Ahead of the furrow mirr was still exclusively dorsal, and slp ventral, but now the two expression domains did not abut; instead, a gap opened that appeared to be generated by loss of slp expression in the region close to the D-V midline (white arrowhead in Fig. 1F). RESEARCH ARTICLE 661 Generation of a slp1, slp2 mutant slp1 and slp2 are functionally redundantly elsewhere in Drosophila development (Grossniklaus et al., 1992), and as transcripts from both genes were expressed identically in the developing eye, both needed to be removed for an effective loss-of-function analysis to be performed. The only existing lesion that removes both transcripts (without affecting other loci) is on a CyO balancer chromosome (Grossniklaus et al., 1992; Grumbling and Strelets, 2006) that was not useful to us. We therefore generated a new deficiency mutation (slpS37A) by mobilization of the slpAvm insertion. Molecular characterization of slpS37A showed a deletion that removes the slp1 transcription unit and all intervening sequence to 16 bp downstream of the slp2 transcriptional start site (Fig. 1B). Thus, although the slp2 coding sequence is intact, the promoter sequence is removed. To test whether the lesion is effectively null for both transcripts it was crossed to a slp⌬34B deficiency and a ‘null’ embryonic phenotype of fused denticle belts was observed (not shown) (Grossniklaus et al., 1992). slp phenotypes in the ventral eye slpS37A mutants are embryonic lethal, and to assess Slp function in the developing eye, clones were induced at various developmental stages and examined in late-third-instar discs. Regardless of the time of induction, ventral clones showed significant differences in appearance and frequency to their dorsal counterparts (Fig. 1G-I). Clones induced at 0-24 hours after egg lying (AEL), although frequently present dorsally, were largely absent from the ventral domain (Fig. 1G). Clones induced at 24-48 hours AEL occurred in the ventral region of Fig. 2. The reciprocal effects of ectopic expression or loss of Slp and Iro. (A-A⬙) mirr-lacZ (red) is repressed by ectopic slp (green, arrow). Inset in A⬙ is higher magnification showing the autonomous repression of mirr-lacZ (arrows mark diminishing staining inside dorsal slp). (B-B⬙) slp-lacZ (red) is ectopically expressed in the inner regions of iro mutant clone (loss of green). Note that the ectopic slp-lacZ evident dorsal to the arrowed clone is an outgrowth of eye tissue into the head capsule. Why slp is non-autonomously induced in such outgrowths is unclear. (C-C⬙) slp-lacZ (red) is repressed by ectopic mirr (green, arrow). Inset in C⬙ shows the non-autonomous effects on slp-lacZ. Immediately surrounding the clone staining is absent, and surrounding this is a ring of cells showing decreased expression. (A, B, C) Schematic summaries of the effects observed. DEVELOPMENT Dorsoventral signaling in the fly eye RESEARCH ARTICLE the eye discs but differed from their dorsal counterparts; they were significantly less frequent, and were found predominantly behind the MF (Fig. 1H). Cells in the posterior regions of the eye are taken out of the cell cycle and ‘frozen’ into the ommatidial lattice earlier than their anterior counterparts. Thus, the absence of anterior/ventral clones suggests that perduring slp gene product allows mutant cells to survive for a time (the posterior clones; white arrowheads in Fig. 1H), but that more extensive proliferation leads to further dilution of the gene product, followed by loss of the cells. Surviving ventral clones were also rounded compared with their dorsal counterparts (Fig. 1H), suggesting an adhesive difference between wild-type and slp ventral cells. Further support for the perdurance phenomenon came from clones induced at 48-72 hours AEL; now they occurred on both sides of the MF, but still showed smoother borders than their dorsal counterparts (white arrowheads in Fig. 1I). These data suggest that Slp function is required for the growth/survival/persistence of cells in the ventral region of the disc. In an attempt to generate large slp patches, we generated slp clones in a Minute background, but these clones were not appreciably different from those generated in a wild-type background (not shown). To determine whether the slp cells were removed by apoptosis, slpS37A clones were induced in the presence of the baculovirus apoptotic caspase inhibitor P35 (Hay et al., 1994). Again, these clones were similar to those induced in wild-type tissues, and thus the under-representation of ventral slpS37A cells does not appear to result from programmed cell death. Development 134 (4) Slp and Iro are mutually repressive transcription factors Since Slp and Iro are transcription factors expressed in abutting domains of the second-instar disc (Fig. 1E), we examined the ability of one to regulate the expression of the other. When slp1 was ectopically expressed in dorsal clonal patches (marked by the coexpression of GFP), a correlating cell-autonomous suppression of mirr transcription was observed (Fig. 2A). Some of the Slpexpressing cells showed residual mirr-lacZ expression (inset in Fig. 2A⬙), but this represented significantly reduced staining, suggesting that these cells are in the process of extinguishing mirr expression. Furthermore, the lacZ gene product has long perdurance. Thus, Slp appears to be an effective suppressor of iro gene expression. When mutant clones for all three genes of the iro gene complex (iroDFM3) were induced in dorsal regions, slp transcription was coincidentally upregulated (Fig. 2B). However, slp was typically expressed in the center of such clones and not at their peripheries (Fig. 2B). This observation suggests: (1) that Iro expression normally functions to repress dorsal expression of slp; and (2) that the creation of an Iro+Iro– interface generates an influence that locally suppresses slp expression (the absence of slp expression at the clone periphery). When mirr was ectopically expressed in ventral clones, a correlating suppression of endogenous slp transcription occurred (Fig. 2C). Again, a non-autonomous effect occurred and slp expression in the cells surrounding the clone was also Fig. 3. (A,B) Double staining of fng in situ hybridization (red) and mirr-lacZ expression (green) in second (A) and mid-third (B) instars. Arrowhead in B indicates the interface between fng and mirr expression. (B⬘) Arrows show the potential Fng+-Fng– interface in the ventral anterior regions of the eye disc as evidenced by fng expression. (C-E⬙) Regulation of fng expression by Slp and Iro as evidenced by fng in situ hybridization. (C) Clones ectopically expressing slp (green, arrows) show upregulation of fng transcription (red). (D) Clones ectopically expressing mirr (green, arrow) downregulate fng transcription (red). (E) When slp and mirr are co-expressed ventrally, fng transcription (red) is repressed in ventral (arrowhead), but not upregulated in dorsal (arrow) regions. DEVELOPMENT 662 downregulated. Immediately surrounding the clone was a region devoid of slp-lacZ expression, and surrounding that was a band of cells with reduced expression (inset in Fig. 2C⬙). Thus, Iro is an effective repressor of slp expression. Furthermore, this experiment provides another example of an Iro+-Iro– interface generating an influence that locally suppresses slp expression. The sub-viability of ventral slp clones prevented an effective assessment of any role that Slp might play in suppression of ventral iro expression. These above experiments indicate: (1) that Slp and Iro are mutually repressive transcription factors; (2) that Iro actively suppresses slp expression in the dorsal domain; and (3) that an Iro+Iro– interface generates an slp-repressing influence, which we show below to be N activity. The roles of Iro and Slp in regulating fng expression Fng is expressed exclusively in ventral eye disc cells (red in Fig. 3AB), and at the D-V midline a Fng+-Fng– interface is formed (arrowhead in Fig. 3B). Hitherto, Iro has been shown to repress fng transcription dorsally (Cho and Choi, 1998; Dominguez and de Celis, 1998; Papayannopoulos et al., 1998). However, when slp is ectopically expressed in dorsal cells, an upregulation of fng transcription occurs (arrows in Fig. 3C). Correspondingly, when iro is clonally expressed ectopically in the ventral territory, a downregulation of fng transcription occurs (arrow in Fig. 3D). This raised the question of whether Iro prevented dorsal fng expression indirectly, by repressing slp expression. When slp and mirr were coexpressed in clonal patches (both dorsally and ventrally), they behaved as mirr alone; there was no dorsal fng upregulation (arrow in Fig. 3E) and there was repression of ventral fng expression (arrowhead in Fig. 3E). Furthermore, we note that surviving ventral slp mutant clones do not show phenotypes typical of Fng+-Fng– interfaces (not shown), which would have arisen if Slp activated ventral fng expression. Thus, it appears that Slp does not promote the ventral expression of fng. Notch activity suppresses slp expression In second-instar eye discs, the Slp and Iro domains directly abut at the midline of the disc (Fig. 1E), and yet by the late third instar a gap opens between them (Fig. 1F). Furthermore, in the clonal experiments described above, whenever an Iro+-Iro– interface was generated, a local downregulation of slp transcription was observed RESEARCH ARTICLE 663 (Fig. 2B-C). Thus, wherever we observed an Iro+-Iro– interface, whether at the normal midline or in clonal experiments, there was a local downregulation of slp expression. Since Iro+-Iro– interfaces establish Fng+-Fng– interfaces (which lead to local N activation), we wondered whether N was the slp-repressing signal. When a constitutively activated form of the N protein (Nintra) (Struhl et al., 1993) was expressed in the ventral regions of the eye discs, a corresponding downregulation of slp expression occurred (Fig. 4A). Thus N activation appears sufficient to repress slp expression, suggesting that the gap that opens up between the Iro and Slp domains in the third-instar disc is caused by the local N activation at the midline interface. If this were true, then removal of N function should prevent the formation of the gap. To test whether endogenous N gene function was required for the downregulation of slp, Nts mutant (Shellenbarger and Mohler, 1978) larvae were grown at the permissive temperature until early third instar and then shifted to 31°C for 24 hours before dissection and staining. Such discs showed a complete absence of slp repression, and the Slp and Iro domains abutted along the length of the midline interface (Fig. 4C). The disc shown in Fig. 4C (Nts/Y) was treated identically to the control shown in Fig. 4B (Nts/+), but has a more immature appearance because N also functions in MF progression (Baonza and Freeman, 2001). These experiments show that ectopic N activity suppresses slp, and that loss of endogenous N activity prevents the normal downregulation of slp expression. Thus, we infer that N activity is the influence present at Iro+-Iro– interfaces that downregulates slp expression. The regulation of Dl and Ser expression In the eye, Iro+-Iro– interfaces generate corresponding Fng+-Fng– interfaces, which regulate the activity of the two N ligands Delta (Dl) and Serrate (Ser), leading to a local upregulation of N activity. One of the outputs of the N signaling is a local increase in the expression of its ligands, which thereby further strengthens the N activity (de Celis and Bray, 1997). In the second-instar eye disc, Dl was ubiquitously expressed (red in Fig. 5A), whereas Ser was expressed in the region of the D-V border (arrow in Fig. 5A) (Cho and Choi, 1998; Dominguez and de Celis, 1998; Papayannopoulos et al., 1998). In the third larval instar, N ligand upregulation occurred in two positions – one associated with the Iro+-Iro– interface, and the other not. These will be described below. Fig. 4. N activation downregulates slp expression. (A-A⬙) Clones of Nintra (green, arrows) show downregulation of slp-lacZ expression (red). (B,C) The effects of a 24-hour temperature shift on Nts/+ control discs (B) and Nts/Y experimental discs (C) stained for slp-lacZ (green) and Mirr (red). In the control, the gap (arrowhead) forms normally, but in the Nts/Y, slp transcription is unaffected and the gap fails to form. DEVELOPMENT Dorsoventral signaling in the fly eye 664 RESEARCH ARTICLE Development 134 (4) Fig. 5. N activity and expression of its ligands in second and third-instar eye discs. (A-A⬙) In late-secondinstar eye discs, Ser (arrow, green) is expressed in a broad swath about the D-V midline, whereas Dl (red) expression is more general. (B) Mid-thirdinstar N activation evidenced by N-GV/UAS-lacZ staining. Ahead of the wave front, strong N activity is in a line running from the D-V midline and curving up into the anterior dorsal region (arrow), and weak N activity is detected in the ventral part (arrowhead). (C) Ser (arrows and arrowheads, green) is expressed in a domain corresponding to the lines of both strong and weak N activation but occupying broader areas. (C⬘) Dl (arrows, red) is upregulated in a line corresponding to the profile of strong N activation, and DI expression can also be seen weakly in a position corresponding to the weaker N activation (arrowhead). (D) Late-third-instar N activity (blue) is detected in the region of the D-V midline and head capsule on both the dorsal (arrow) and ventral (arrowhead) side. (D⬘) Ser (green) is expressed in a broader region corresponding to N activity. (D⬙) Dl (red) is mainly expressed behind the MF, but is also expressed at the anterior extreme in both dorsal (arrow) and ventral (arrowhead) regions. The eye disc portion of the eye-antennal disc contains both the presumptive retina and head capsule. The presumptive retina occupies the majority of the tissue extending from the posterior margin of the disc to an anterior limit defined by the extent of Eyegone (Eyg) expression (blue in Fig. 6A-B) (Dominguez et al., 2004). It was found that the Iro+-Iro– interface runs as a straight line through the eye region, but as it moves into the region of the presumptive head capsule it curves sharply dorsal (Fig. 6B). Expression of Dl and Ser was upregulated about this border. Dl appeared as a stripe in the dorsal cells next to the midline, running as a straight line through the eye field and curving dorsally with the interface (red in Fig. 5C⬘). Ser expression was found to be more complicated: in the region of the eye, it filled the gap vacated by Slp expression (green in Fig. 6A-B), and as it entered the head capsule region it bifurcated into two ‘spurs’; the stronger one following the dorsally curving Iro+-Iro– interface, and the weaker one bending ventrally (green in Fig. 5C-D and Fig. 6A-B). Dl and Ser expression in the ventral spur To determine whether the ventral spur of Ser expression was associated with N activity, we stained a reporter line for N activity (N-GV) (Chung and Struhl, 2001). Strong N activity was found in the region of the Iro+-Iro– interface, both in the midline region of the eye and in the dorsally curving part in the presumptive head capsule (arrows in Fig. 5B,D). Additionally, however, N activity was detected in the region corresponding to the ventral spur of Ser expression (arrowhead in Fig. 5B,D). This expression is weaker than its dorsal counterpart, but it was consistently observed. In addition, a weak upregulation of Dl expression was often detected at this location (arrowhead in Fig. 5C⬘,D⬙). This ventral spur of N signaling is distant from and, by inference, independent of the Iro+-Iro– interface. However, an additional Fng+-Fng– interface appears to be present in the ventral anterior region of the eye disc (arrows in Fig. 3B⬘) which corresponds to the position of the ventral spur of N activity. Thus, this appears to be an Iroindependent Fng+-Fng– interface that regulates N signaling in the extreme ventral anterior of the eye disc. We note that the mirrlacZ line used in our experiments (mirrF7) is also a w+ enhancer trap for mirr (McNeill et al., 1997), and in the adult the pigment fills the dorsal eye stopping at the equator represented by the DV midline (Tomlinson, 2003). Thus, the Iro+-Iro– interface defines the midline of the eye, and the ventral spur of N activity appears to play no role in this patterning. Ser upregulation correlates with downregulation of slp expression In second-instar larvae, Ser is expressed about the D-V midline in both Slp- and Iro-expressing cells. Thus, Ser expression is not ‘generically’ suppressed by these transcription factors, However, Slp appears to play a role in regulating the subsequent upregulation of Ser. In the eye region, the Ser upregulation occurs in the gap left by the receding slp expression. N expression engenders both these effects: it upregulates Ser expression (Papayannopoulos et al., 1998) and downregulates slp expression. In the mid-third larval instar, a ‘peninsula’ of slp expression was seen to project from the main ventral body of expression and follow the dorsally curving Iro+-Iro– DEVELOPMENT Dl and Ser regulation about the Iro+-Iro– interface Dorsoventral signaling in the fly eye RESEARCH ARTICLE 665 Fig. 6. (A-A⬙) High levels of Ser expression (green) correspond with the loss of slp-lacZ (red) in thirdinstar eye discs. eyg-Gal4/UAS-GFP expression (blue) demarcates the eye field (dotted line indicates the anterior extreme of the eye); Ser expression anterior to this relates to patterning of head capsule tissues. Arrow in A⬘ indicates the downregulation of slp expression in the ventral spur corresponding with high Ser expression. (B-B⬙) Ser (green) is expressed in cells not expressing mirr-lacZ (red), except in the most anterior regions (arrow in B⬙). (C-D⬙) The roles of N, Slp and Iro in regulating Ser upregulation in third-instar discs. (C) Ectopic expression of slp (red, arrow) in the region of the gap shows a downregulation of Ser expression (green). Note that slp does not repress Ser completely. (D) Ectopic expression of mirr in the region of the gap (red, arrow) shows a downregulation of Ser expression (green). Note that the nonautonomous induction of Ser (arrowhead) is likely to be caused by the new Fng+-Fng– interface induced in the ventral cells. Slp represses Ser expression As there were two positions in the eye where Ser upregulation correlated with a downregulation of slp, we wondered whether this was a causal or a coincidental relationship. Since N activation is required to repress slp expression, and as Ser is required for that N activation, we inferred that Ser expression was required for the downregulation of slp. But was slp downregulation required for the upregulation of Ser? slp expressed under a (non-N-inducible) heterologous promoter in the region of the gap induced a strong reduction in Ser staining (arrow in Fig. 6C). This was a reduction, not an extinction of Ser expression, but the effect was clear and reproducible. Furthermore, expression of mirr also repressed Ser expression (arrow in Fig. 6D). Thus both Slp and Iro are able to suppress the upregulation of Ser. DISCUSSION We report here that Slp and Iro are mutually repressive transcription factors expressed in the respective ventral and dorsal domains of the developing eye. In the second instar the two directly abut at the D- V midline (Fig. 1E), but N signaling directs the local downregulation of slp, leading to the emergence of a gap between the two expression domains (white arrowhead in Fig. 1F). N signaling also instructs the upregulation of its ligands: Dl expression is increased in dorsal cells next to the midline, and Ser is expressed in a wider ventral domain that corresponds to the gap left by the receding slp expression. The removal of Slp (the formation of the gap) appears necessary for the effective upregulation of Ser. Below we discuss these results and address related issues. slp mutant clones slp clones in the dorsal region of the developing eye appear normal and healthy which suggests that there is nothing intrinsically wrong with slp mutant cells. Yet, when induced in the ventral (Slpexpressing) domain, the clones are sub-viable (white arrowheads in Fig. 1H-I). Furthermore, the cells that survive are likely to carry perduring gene function, so we suspect that fully mutant ventral slp cells do not survive (arrow in Fig. 1G-H). Neither the use of the Minute technique, nor the presence of the apoptotic inhibitor P35, affected the survival of the clones. These results suggest that the slp mutant cells do not suffer a simple growth disadvantage with respect to their Slp-expressing neighbors, nor are they removed by apoptosis. Surviving clones (white arrowheads in Fig. 1H) show smooth borders, which suggests that they are ‘immiscible’ with their Slp+ neighbors, and that some adhesion mechanism may be DEVELOPMENT interface (on its anterior side) (red in Fig. 6A). The ventral spur of Ser upregulation cut through this peninsula, and here a clear and correlating downregulation of slp expression was observed (arrow in Fig. 6A⬘). RESEARCH ARTICLE responsible for the removal of the cells. However, we did not observe extrusion of mutant tissue from the epithelium that would be expected if ‘immiscibility’ was responsible for the loss of the mutant cells. The relationship between Slp and Iro Slp and Iro are mutually repressive transcription factors: when one is expressed in the domain of the other, it suppresses the other’s expression. In dorsal iro mutant clones, slp is transcriptionally activated (Fig. 2B), indicating that Iro acts to prevent dorsal expression of slp. The reciprocal experiment of examining iro transcription in ventral slp clones was confounded by our inability to generate effectively null slp clones. However, ventral slp clones induced at all stages throughout larval development showed no evidence of mirr-lacZ expression (not shown). Furthermore, we note that the gap is made from ventral cells that turned off slp expression, and that iro expression does not subsequently turn on in those cells. We can view the relationship between Slp and Iro in one of two ways. First, both proteins function in their respective domains to suppress the expression of the other, and technical problems have prevented us from detecting the ventral suppression of iro by Slp. It this were the case, then both would be likely to function to ensure the emergence of a sharp interface between the two domains, in the manner described in the Introduction for vertebrate neural tube cell fates. The second possibility is that Iro represses dorsal slp expression, but Slp does not repress ventral iro expression. In this second model, what would be the function of Slp vis-à-vis Iro? Since Iro suppresses slp (Fig. 2B-C) then this suggests that the ‘ground state’ of the eye tissue is to express slp, and that it is excluded from dorsal expression by Iro. Wingless (Wg) is required for Iro expression (Heberlein et al., 1998), and in the embryonic eye disc primordium slp is expressed in the majority of the eye primordium, being excluded from the dorsal extreme (not shown). wg is expressed in the dorsal region (data not shown) and we speculate that Wg promotes dorsal Iro expression which suppresses the dorsal expression of Slp. Ventral Slp may then function to repress any ventral iro expression engendered by diffusing Wg signaling. The mutual repression of the two transcription factors would then lead to cells adopting either the Slp–/Iro+ or Slp+/Iro– state. Immiscibility effects between the cells would then lead to the formation of the long straight midline border, and in this regard we note that the surviving slp mutant clones round up when surrounded by Slp+ cells (arrowheads in Fig. 1H). Slp and the upregulation of Ser We have described (in two different positions in the eye disc) that loss of slp expression correlates with the upregulation of Ser. One position at which this occurs is the gap that forms next to the Iro+Iro– interface. The other is in the ventral region of the presumptive head capsule ahead of the eye field; here a spur of Ser expression shows a corresponding loss/reduction in slp expression (arrow in Fig. 6A⬘). Not only does the reduction of slp expression correlate with Ser upregulation, but also restoration of Slp to the region of the gap prevents the high levels of Ser normally found there (Fig. 6C). Thus, reduction in slp levels appears necessary for the increase in Ser levels. We note that slp transcription is monitored here using slplacZ, and as lacZ has significant perdurance, the gap is probably larger than that indicated, and any overlap between Ser and slp is likely to be less than that suggested by the images. In wing D-V patterning there is no known ‘equivalent’ of Slp – that is, a transcription factor that initially abuts the D-V interface but is then pushed back by the action of N signaling. Thus, this Development 134 (4) role of Slp appears specific to the eye, and this raises the question of its function. One explanation is that it acts as a brake on the N signaling. A major difference between the eye and wing is that a wave of differentiation sweeps the eye, and the midline N signaling is required specifically ahead of the wave front (the MF). A delicate balance may be required between the amount of N signaling ahead of the MF, and the speed with which the MF progresses across the disc. Slp may provide that control. As a repressor of Ser upregulation, Slp could act as a delay mechanism – slowing the increase in Ser, and thereby slowing the increase in N activation. We thank Richard Mann for comments on the manuscript; Ken Irvine, Maria Leptin, Richard Mann, Helen McNeill, Gary Struhl, Leslie Vosshall, Myriam Zecca, The Developmental Studies Hybridoma Bank and the Bloomington Stock Center for materials; and Natalya Katanayeva for the germline transformation. A.S. personally thanks Yoshinori Tomoyasu for helpful discussion. This work was funded by grant NIH R01EY012536 to A.T. References Baonza, A. and Freeman, M. (2001). Notch signaling and the initiation of neural development in the Drosophila eye. Development 128, 3889-3898. Becker, H. J. (1957). Uber Rontgenmosaikflecken und Defektmutationen am Auge von Drosophila und die Entwicklungphysiologie des Auges. Z. Induk. Abst. Vererb. Lehre. 88, 333-373. Blair, S. S., Brower, D. L., Thomas, J. B. and Zavortink, M. (1994). The role of apterous in the control of dorsoventral compartmentalization and PS integrin gene expression in the developing wing of Drosophila. Development 120, 18051815. Bruckner, K., Perez, L., Clausen, H. and Cohen, S. (2000). Glycosyltransferase activity of Fringe modulates Notch-Delta interactions. Nature 406, 411-415. Cavodeassi, F., Modolell, J. and Campuzano, S. (2000). The Iroquois homeobox genes function as dorsal selectors in the Drosophila head. Development 127, 1921-1929. Cho, K. O. and Choi, K. W. (1998). Fringe is essential for mirror symmetry and morphogenesis in the Drosophila eye. Nature 396, 272-276. Chung, H. M. and Struhl, G. (2001). Nicastrin is required for Presenilin-mediated transmembrane cleavage in Drosophila. Nat. Cell Biol. 3, 1129-1132. Curtiss, J. and Mlodzik, M. (2000). Morphogenetic furrow initiation and progression during eye development in Drosophila: the roles of decapentaplegic, hedgehog and eyes absent. Development 127, 1325-1336. de Celis, J. F. and Bray, S. (1997). Feed-back mechanisms affecting Notch activation at the dorsoventral boundary in the Drosophila wing. Development 124, 3241-3251. Diaz-Benjumea, F. J. and Cohen, S. M. (1993). Interaction between dorsal and ventral cells in the imaginal disc directs wing development in Drosophila. Cell 75, 741-752. Dominguez, M. and de Celis, J. F. (1998). A dorsal/ventral boundary established by Notch controls growth and polarity in the Drosophila eye. Nature 396, 276278. Dominguez, M., Ferres-Marco, D., Gutierrez-Avino, F. J., Speicher, S. A. and Beneyto, M. (2004). Growth and specification of the eye are controlled independently by Eyegone and Eyeless in Drosophila melanogaster. Nat. Genet. 36, 31-39. Gomez-Skarmeta, J. L., Diez del Corral, R., de la Calle-Mustienes, E., FerreMarco, D. and Modolell, J. (1996). Araucan and caupolican, two members of the novel iroquois complex, encode homeoproteins that control proneural and vein-forming genes. Cell 85, 95-105. Grossniklaus, U., Pearson, R. K. and Gehring, W. J. (1992). The Drosophila sloppy paired locus encodes two proteins involved in segmentation that show homology to mammalian transcription factors. Genes Dev. 6, 1030-1051. Grumbling, G. and Strelets, V. (2006). FlyBase: anatomical data, images and queries. Nucleic Acids Res. 34, D484-D488. Haines, N. and Irvine, K. D. (2003). Glycosylation regulates Notch signalling. Nat. Rev. Mol. Cell Biol. 4, 786-797. Hay, B. A., Wolff, T. and Rubin, G. M. (1994). Expression of baculovirus P35 prevents cell death in Drosophila. Development 120, 2121-2129. Heberlein, U., Borod, E. R. and Chanut, F. A. (1998). Dorsoventral patterning in the Drosophila retina by wingless. Development 125, 567-577. Irvine, K. D. (1999). Fringe, Notch, and making developmental boundaries. Curr. Opin. Genet. Dev. 9, 434-441. Irvine, K. D. and Wieschaus, E. (1994). fringe, a Boundary-specific signaling molecule, mediates interactions between dorsal and ventral cells during Drosophila wing development. Cell 79, 595-606. Jessell, T. M. (2000). Neuronal specification in the spinal cord: inductive signals and transcriptional codes. Nat. Rev. Genet. 1, 20-29. DEVELOPMENT 666 Klein, T. and Arias, A. M. (1998). Interactions among Delta, Serrate and Fringe modulate Notch activity during Drosophila wing development. Development 125, 2951-2962. Lawrence, P. A. and Struhl, G. (1996). Morphogens, compartments and pattern: lessons from Drosophila? Cell 85, 951-961. Lee, T. and Luo, L. (1999). Mosaic analysis with a repressible cell marker for studies of gene function in neuronal morphogenesis. Neuron 22, 451-461. McNeill, H., Yang, C. H., Brodsky, M., Ungos, J. and Simon, M. A. (1997). mirror encodes a novel PBX-class homeoprotein that functions in the definition of the dorsal-ventral border in the Drosophila eye. Genes Dev. 11, 1073-1082. Moloney, D. J., Panin, V. M., Johnston, S. H., Chen, J., Shao, L., Wilson, R., Wang, Y., Stanley, P., Irvine, K. D., Haltiwanger, R. S. et al. (2000). Fringe is a glycosyltransferase that modifies Notch. Nature 406, 369-375. Munro, S. and Freeman, M. (2000). The notch signaling regulator fringe acts in the Golgi apparatus and requires the glycosyltransferase signature motif DXD. Curr. Biol. 10, 813-820. Panin, V. M., Papayannopoulos, V., Wilson, R. and Irvine, K. D. (1997). Fringe modulates Notch-ligand interactions. Nature 387, 908-912. Papayannopoulos, V., Tomlinson, A., Panin, V. M., Rauskolb, C. and Irvine, K. D. (1998). Dorsal-ventral signaling in the Drosophila eye. Science 281, 20312034. RESEARCH ARTICLE 667 Riechmann, V., Irion, U., Wilson, R., Grosskortenhaus, R. and Leptin, M. (1997). Control of cell fates and segmentation in the Drosophila mesoderm. Development 124, 2915-2922. Sato, A., Kojima, T., Ui-Tei, K., Miyata, Y. and Saigo, K. (1999). Dfrizzled-3, a new Drosophila Wnt receptor, acting as an attenuator of Wingless signaling in wingless hypomorphic mutants. Development 126, 4421-4430. Shellenbarger, D. L. and Mohler, J. D. (1978). Temperature-sensitive periods and autonomy of pleiotropic effects of l(1)Nts1, a conditional notch lethal in Drosophila. Dev. Biol. 62, 432-446. Struhl, G. and Adachi, A. (1998). Nuclear access and action of notch in vivo. Cell 93, 649-660. Struhl, G., Fitzgerald, K. and Greenwald, I. (1993). Intrinsic activity of the Lin12 and Notch intracellular domains in vivo. Cell 74, 331-345. Tomlinson, A. (2003). Patterning the peripheral retina of the fly: decoding a gradient. Dev. Cell 5, 799-809. Wehrli, M. and Tomlinson, A. (1998). Independent regulation of anterior/posterior and equatorial/polar polarity in the Drosophila eye; evidence for the involvement of Wnt signaling in the equatorial/polar axis. Development 125, 1421-1432. Yang, C. H., Simon, M. A. and McNeill, H. (1999). mirror controls planar polarity and equator formation through repression of fringe expression and through control of cell affinities. Development 126, 5857-5866. DEVELOPMENT Dorsoventral signaling in the fly eye