Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
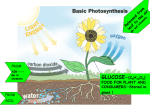
3.3 Photosynthesis E X P E C TAT I O N S Describe the energy transformations that occur in photosynthesis. Describe the role of enzymes in metabolic reactions in chloroplasts. Investigate and explain how the structure of molecules can influence metabolic rate. Compare the structure and function of chloroplasts and mitochondria. Photosynthesis is one of the most important chemical processes on Earth. Photosynthesis involves the use of energy from light to form carbohydrates. Organisms that manufacture their own food (autotrophs), such as plants, algae, cyanobacteria, and photosynthetic bacteria, do so through photosynthesis. Autotrophs form the base of the food chain for virtually all communities of heterotrophs, which must eat to obtain nutrients. The process of photosynthesis also produces the oxygen found in the atmosphere. Oxygen is used by organisms for many processes, such as aerobic cellular respiration. The overall equation for photosynthesis is as follows: 6CO2 + 6H2O + energy → C6H12O6 + 6O2 carbon water glucose oxygen dioxide The process of photosynthesis is believed to have originated in bacteria. Some of these bacteria were able to produce oxygen. Other bacteria were able to carry out a different form of photosynthesis but did not produce oxygen. In 2000, biochemists led by Dr. Carl Bauer at the University of Indiana found that non-oxygen-producing species (purple and green bacteria) are the most ancient photosynthetic bacteria. The oxygen-producing cyanobacteria that exist today (see Figure 3.19) evolved from a non-oxygen-producing bacteria called heliobacteria. As you may have noticed, the overall equation for photosynthesis is exactly the opposite of the equation for aerobic cellular respiration. This does not mean, however, that the reactions follow the same course in reverse. Photosynthesis requires structures and metabolic processes similar to those used in mitochondria: electron transport chains, dissolved enzymes, and a membrane-enclosed space for chemiosmosis. Figure 3.18 The process of photosynthesis is one of the most important of all life processes. Green plants use carbon dioxide and water to produce oxygen and food. Figure 3.19 Cyanobacteria eventually gave rise to the structures that carry out photosynthesis in the algae and green plants of today. Chapter 3 Cellular Energy • MHR 83 leaf cross section mesophyll cell opening for C O2 to enter leaf chloroplast vacuole nucleus cell wall granum lamella thylakoid stroma Structure of Chloroplasts In plant cells, photosynthesis occurs within chloroplasts. Chloroplasts have a double membrane and contain membrane pockets called thylakoids (see Figure 3.20). Thylakoids occur in stacked, parcel-like structures called grana (singular granum), which are held together by support structures called lamellae. The stroma, a thick, enzyme-rich liquid, fills the interior of each chloroplast. Mesophyll cells in the leaves of plants are specialized for photosynthesis and contain numerous chloroplasts. These cells provide the chloroplasts with the two important ingredients necessary for photosynthesis — carbon dioxide and water. Gas exchange (oxygen and carbon dioxide) occurs through pores on the underside of leaves, and water is delivered via veins that extend to the roots of the plant. Within the grana, solar light energy is captured by the thylakoids. This energy is used to form ATP molecules, which fuel the production of carbohydrates. These carbohydrate molecules are then used to synthesize glucose — the molecules used in cellular respiration. The thylakoid membrane in the chloroplast is the site of ATP production, using chemiosmosis and complex structures functionally similar to those found in mitochondria. Stages of Photosynthesis As the previous section suggests, there are two main stages of photosynthesis: the photo and synthesis stages. The first stage of photosynthesis converts 84 MHR • Unit 1 Metabolic Processes inner membrane outer membrane Figure 3.20 Structure of a chloroplast solar energy into chemical energy. The second stage uses this energy to produce PGAL, which is then used to form glucose (see Figure 3.21). The photo reactions require light and are called lightdependent reactions. The synthesis reactions do not require light directly, and are called lightindependent reactions. However, light seems to be important in activating enzymes in both the photo and synthesis reactions. sunlight photo reactions A synthesis reactions organic molecules B Figure 3.21 Two stages of photosynthesis. The first stage (A) consists of reactions that require light energy. The second stage (B) involves the synthesis of glucose molecules. As light strikes the leaf of a plant, the energy is captured by pigments in the chloroplasts. These pigments, known as chlorophylls, absorb various wavelengths of visible light (see Figure 3.22). The two most important types of chlorophyll are chlorophyll a and chlorophyll b. Photosynthesis is most active at light wavelengths of about 400 nm to 450 nm and 650 nm to 700 nm. The colour of chlorophyll, green, is a result of the absorption of mainly blue and red parts of the visible light spectrum. In the following MiniLab, you will extract chlorophyll from leaves and examine the colour and properties of both types of chlorophyll. B Chlorophylls a and b absorb certain wavelengths of visible light. chlorophyll a yellow and green transmitted visible spectrum prism Relative light absorption A Visible light represents only a small segment of the electromagnetic spectrum. chlorophyll b solution of chlorophyll white light 400 450 500 550 600 650 700 Wavelength (nm) Figure 3.22 The relative absorption of light by chlorophylls a and b. MINI LAB Photosynthetic Pigments In this lab you will investigate the colours of chlorophylls, the photosynthetic pigments found in many plants. You will need to produce a concentrated extract of chlorophylls. The materials you will require are: 15 g fresh or frozen spinach, 50 mL isopropyl alcohol, food blender, 100 mL beaker, funnel, filter paper, strong light source (for example, slide projector). 2. What wavelengths of visible light are absorbed by the chloroplasts? 3. When you viewed the solution, the effect you saw is called fluorescence. What colours and wavelengths were produced? Using the food blender, grind the spinach with 50 mL of isopropyl alcohol. Filter the extract through several layers of filter paper in the funnel. Then, in a darkened room, shine a strong beam of light at a sample of the filtered extract. Observe the colour of the chlorophylls by viewing the sample at a slight angle and then at a right angle to the beam of light. Describe the appearance of the chlorophylls as viewed from both angles. Analyze 1. What colours of the visible light spectrum are absorbed by chlorophyll as part of the photo reaction? Photosynthetic pigments Chapter 3 Cellular Energy • MHR 85 Light energy is absorbed by a network of chlorophyll molecules known as a photosystem (see Figure 3.23). These chlorophyll molecules are known as antenna pigments because they collect and channel energy. This energy causes electrons in the chlorophyll molecules to become energized. Energy from these electrons is passed from one chlorophyll molecule to another in the photosystem. Eventually the energy reaches the reaction centre, a specific chlorophyll a molecule. Only one in 250 chlorophyll molecules forms a reaction centre. A unit of several hundred antenna pigment molecules together with a reaction centre is called a photosynthetic unit. The large number of antenna pigment molecules in each photosynthetic unit allows the reaction centre to be supplied with the greatest possible amount of energy. Once the energy has reached the reaction centre, an electron acceptor receives the energized electron. Energy from these electrons is used to move H+ ions into the thylakoid interior for ATP production. ADP + ATP n s p o rt s y st e m Pi e− n tr a reaction-centre chlorophyll a electron acceptor tr o chlorophyll molecules electron acceptor sun ele c light photosystem 700. After the electron acceptors receive the energized electrons from the reaction centre, the electrons flow through an electron transport system. Here the electrons are passed from one electron carrier to another. Some of these carriers are cytochrome molecules. As the electrons pass through the system, they release energy that is used to phosphorylate ADP molecules to produce molecules of ATP. This process is called cyclic photophosphorylation, because after the ATP molecules are produced the electrons are cycled back into the photosystem. Only ATP molecules are produced by photosystem 700. energy of electron Photosystems e− reaction– centre chlorophyll a pigment complex photosystem 700 Figure 3.24 The cyclic electron pathway. In photosystem 700, Figure 3.23 A photosystem works by passing light energy from one molecule of chlorophyll to another. Cyclic Electron Pathway There are two types of photosystems. Photosystem 700, which absorbs light 700 nm in wavelength, is used by some photosynthetic bacteria. This photosystem contains molecules of chlorophyll a, which is found in cyanobacteria and all photosynthetic eukaryotes (such as green plants). Figure 3.24 shows how electrons pass through 86 MHR • Unit 1 Metabolic Processes electrons are recycled after their energy is used to form ATP molecules. WEB LINK www.mcgrawhill.ca/links/biology12 To find out more about how photosynthetic bacteria perform photosynthesis, go to the web site above, and click on Web Links. Note the types of bacteria that use photosynthesis to produce ATP. Prepare a chart that compares and contrasts the methods by which bacteria and plants perform photosynthesis. Non-cyclic Electron Pathway Photosystem 680 absorbs light 680 nm in wavelength. The shorter the wavelength of light, the higher its energy. Therefore, photosystem 680 is more powerful than photosystem 700 because photosystem 680 can capture higher-energy light. In addition to chlorophyll a, photosystem 680 contains molecules of chlorophyll b. It also contains molecules of chlorophyll c, chlorophyll d, and accessory pigments such as carotenes, xanthophylls, and anthocyanins. The pigments other than chlorophyll a aid in absorbing wavelengths of light not absorbed by chlorophyll a. Green plants, algae, and cyanobacteria (unlike other bacteria species) use both photosystems 680 and 700 to carry out photosynthesis. In this case, electrons from photosystem 680 are shunted to photosystem 700, as shown in Figure 3.25. The energy from electrons in photosystem 680 is used to produce ATP molecules. These electrons then move to photosystem 700 where, after becoming energized, they are taken up by NADP+ (nicotinamide adenine dinucleotide). After NADP+ accepts two electrons and a hydrogen ion (H+ ), it becomes the coenzyme NADPH. The production of NADPH and ATP are endothermic reactions, which require an input of energy. The ATP and NADPH molecules are then used in the synthetic steps to produce glucose. After ATP molecules are produced by photosystem 680, electrons that have passed through the electron transport system are not cycled back into photosystem 680. This type of ATP production is called non-cyclic photophosphorylation. However, photosystem 680 requires electrons to keep the photosystem operating. After photosystem 680 transfers an electron to the electron acceptor, photosystem 680 captures an electron from a Z enzyme. This enzyme is responsible for splitting water molecules into hydrogen ions and oxygen molecules and sun electron acceptor energy of electron sun NADP + e− electron acceptor NADPH H+ e− ele e − c tr on tra nsp ort sys tem ADP + Pi e− ATP reaction–centre chlorophyll a e− reaction–centre chlorophyll a photosystem 700 photosystem 680 e − H+ Z enzyme H2O CO2 2 H+ 1 O 2 2 Figure 3.25 The non-cyclic electron pathway. Electrons from water move from photosystem 680 to photosystem 700 and then to NADP+ . The ATP and NADPH molecules that are C16 H12 O6 synthesis reactions produced by these reactions fuel the synthesis reactions that form glucose. Chapter 3 Cellular Energy • MHR 87 channelling electrons to the electron acceptor (see Figure 3.25). This process is called photolysis because light energy is required to split bonds within the water molecule. All of the oxygen that we breathe, and all the oxygen in Earth’s atmosphere, has been generated through the photolysis stage of photosynthesis. In addition to passing electrons from water to chlorophyll molecules, the Z enzyme that performs photolysis also donates a hydrogen ion from the same water molecule to the reaction-centre of photosystem 680. This hydrogen ion joins the electron in its journey along the electron transport chain. The electron–hydrogen ion combination supplies energy to an electron transport chain comprised of cytochrome enzymes. This chain of enzymes in turn drives a proton pump, similar to the one you learned about in chemiosmosis in the mitochondrion. The photosynthetic proton pump, like proton pumps in the electron transport chain of the mitochondrion, moves H+ ions out of the stroma, into a membrane-enclosed space, as illustrated by Figure 3.26. Just as the inner membrane of the mitochondrion contains an ATP synthase complex that opens to the matrix, the thylakoid membrane of the chloroplast contains an ATP synthase complex where H+ ions flow through to the stroma and energize the phosphorylation of ADP. This process is called photophosphorylation. ELECTRONIC LEARNING PARTNER To learn how the intensity and wavelength of light can affect ATP/NADPH production in chloroplasts, go to your Electronic Learning Partner now. The thylakoid space serves as a reservoir for hydrogen ions. Every time the Z enzyme splits water to form two hydrogen ions, the thylakoid space receives them. Whenever photosystem 680 donates an electron to the electron transport system, giving up energy along the way to drive the proton pump, hydrogen ions move in from the stroma. A hydrogen ion gradient is formed when the thylakoid space contains more hydrogen ions than the stroma. The movement of hydrogen ions across the thylakoid membrane releases energy that is used in ATP synthesis. This gradient forces the hydrogen ions through the ATP synthase complex that resides on the membrane of the thylakoid body. This movement of hydrogen ions provides the energy required to join ADP and Pi in the chemiosmotic synthesis of ATP. photosystem II light photosystem I cytochrome complex antenna complex stroma light antenna complex + H e NADP + + H+ NADP reductase − e− e NADPH proton pump − Z enzyme Figure 3.26 Within the thylakoid H+ H2O H+ 1 O 2 2 thylakoid membrane + synthesis reactions + 2 H H+ H+ ATP + H thylakoid space H+ ATP synthase 88 MHR • Unit 1 Metabolic Processes ADP + Pi membrane, enzyme complexes pump hydrogen ions from the stroma into the thylakoid space. This process forms a hydrogen ion gradient. As hydrogen ions flow down the gradient and back into the stroma through the ATP synthase complex, ATP molecules are formed. As you can see in this diagram, photosystem 680 is also called photosystem II. Photosystem 700 is also called photosystem I. membrane are used during the synthesis reactions to produce organic molecules from carbon dioxide. ATP and NADPH molecules are formed on the thylakoid membrane by means of the ATP synthase complex and the NADP reductase complex, respectively (see Figure 3.26). The ATP and NADPH The electrons from photosystem 680 energize electrons that travel through the electron transport chain in the thylakoid to pump protons. The electrons lose energy after they move through the electron carriers. At photosystem 700, the electrons are re-energized by light energy. These two electrons now move along the final carriers of the electron transport chain to the NADP reductase complex. Here, two electrons are transferred to NADP+ , which also combines with a hydrogen ion to form the reduced NADPH, as shown in Figure 3.26. Both chloroplasts and mitochondria use chemiosmosis to produce ATP. These organelles also rely on an electron transport chain to power proton pumps and move electrons to an electron acceptor that removes them (such as water or NADPH). The proton pumps create the hydrogen ion gradient that both organelles use to make ATP. Chloroplasts and mitochondria even share the basic construct of an ATP synthase complex, which is remarkably similar in both structures. However, there are some differences in the way that phosphorylation occurs in the two organelles, as summarized in Table 3.3. Table 3.3 Differences in phosphorylation between mitochondria and chloroplasts Oxidative phosphorylation in mitochondria Electrons in the electron transport chain are supplied by the oxidation of food molecules. food energy → ATP molecules Photophosphorylation in chloroplasts Electrons in the electron transport chain are extracted from water during photolysis and passed on to pigment molecules (driven by captured solar energy) to donate them to the chain. light energy → ATP molecules The Calvin Cycle In photosynthesis, both the NADPH and the ATP produced by the photo reactions in the thylakoid Inner membrane pumps hydrogen ions from the matrix to the intermembrane space. Intermembrane space serves as proton reservoir. Thylakoid membrane pumps hydrogen ions from the stroma to the thylakoid interior. Thylakoid interior pools ions. ATP synthase resides between the membrane and the matrix, producing ATP molecules as hydrogen ions move back into the matrix from the intermembrane space. ATP synthase bridges the thylakoid membrane and its interior, producing ATP molecules as hydrogen ions move into the stroma. 3 CO2 Metabolites of the Calvin Cycle 3 RuBP C5 6 PGA C3 carbon fixation RuBP ribulose bisphosphate PGA 3-phosphoglycerate PGAP 1,3-bisphosphoglycerate PGAL glyceraldehyde-3-phosphate 6 ATP 3 ADP + 3 Pi Calvin cycle These ATP 3 molecules ATP were produced by the photo reaction. 6 ADP + 6 Pi reduction re-formation of RuBP 6 PGAP C3 5 PGAL C3 6 NADPH 6 PGAL C3 Figure 3.27 The Calvin 6 NADP + There is a net gain of one PGAL. 1 PGAL C3 These ATP and NADPH molecules were produced by the photo reactions. Glucose phosphate and other organic compounds cycle produces one molecule of PGAL for every three molecules of CO2 that enter the cycle. PGAL is used to form glucose and other organic compounds. Chapter 3 Cellular Energy • MHR 89 molecules formed then leave the thylakoid membrane and enter the stroma, where a series of enzymes perform synthesis reactions in the Calvin cycle. The Calvin cycle is named after biochemist Melvin Calvin. In the late 1940s, Calvin led a team of researchers to determine the steps of this synthesis reaction. Every photosynthetic plant uses the Calvin cycle to form PGAL. PGAL is then used to synthesize many different molecules. Using PGAL as the building block, plants can synthesize amino acids and fatty acids. Other molecules that can be formed from PGAL include fructose phosphate, glucose, sucrose, starch, and cellulose. Although plants synthesize these molecules, not every plant uses the same metabolic pathway. The Calvin cycle has three distinct stages, as shown in Figure 3.27, on the previous page: 1. Stage 1: carbon fixation 2. Stage 2: reduction 3. Stage 3: re-formation of RuBP (ribulose 1,5 bisphosphate) These three stages will now be discussed. Stage 1: Carbon Fixation Carbon fixation is the initial incorporation of carbon into organic molecules. To eventually build complex molecules, such as glucose, plants must first attach carbon to smaller carbon-containing molecules. They do this by taking carbon dioxide from the atmosphere and attaching it to RuBP, ribulose bisphosphate, as shown in Figure 3.28. A six-carbon molecule is the product of this reaction, but this molecule is extremely unstable and immediately splits into two molecules of threecarbon PGA (phosphoglycerate). The enzyme RuBP H2C H2C O C O HC HC H2C O OH RuBP carboxylase O P − O H2C + H+ O C O P OH HC RuBP (Ribulose 1,5 bisphosphate) + H+ O C CO2 + H2O P OH HC P OH O − PGA (phosphoglycerate) Figure 3.28 In the Calvin cycle, C3 fixation produces two three-carbon PGA. The reaction is catalyzed by the enzyme RuBP carboxylase. 90 MHR • Unit 1 Metabolic Processes carboxylase catalyzes this reaction, as shown in Figure 3.28. This reaction is called C3 fixation because it produces two three-carbon molecules of PGA. This molecule then passes into the next stage of the Calvin cycle. C3 fixation is used by plants, such as rice, wheat, and oats, which occur mainly in temperate regions. To form a molecule of glucose (C6H12O6 ), six carbon atoms must be fixed. Figure 3.27 shows that nine molecules of ATP are required to fix the three carbon atoms in the PGAL that is available to be used for glucose production. Therefore, 18 molecules of ATP are needed to fix the six carbon atoms required to form a glucose molecule. In addition to carbon fixation, RuBP carboxylase oxidizes RuBP with O2 to form CO2 by a process called photorespiration. Photorespiration creates an inefficiency in the carbon fixation process, since both the oxidation of RuBP and carbon fixation are catalyzed by the same enzyme — RuBP carboxylase. Both oxygen and carbon dioxide compete to bind with RuBP. The Calvin cycle is an ancient process that developed in an atmosphere with little free oxygen. The rate of reactions in the Calvin cycle increases with temperature to about 25°C. Reaction rate levels out and declines when temperatures approach or exceed 37°C. At warmer temperatures, RuBP carboxylase is mainly involved in oxidizing RuBP, and very little carbon fixation occurs. Thus, plants that live in warmer climates have developed a different approach to fixing carbon. For example, C4 fixation is used by plants, such as sugarcane and corn. In these plants, the Calvin cycle takes place in bundle-sheath cells, as shown in Figure 3.29. Plants that use C4 fixation form the four-carbon oxaloacetate and malate in parenchyma cells. The malate moves into the bundle-sheath cells and a carbon is removed as CO2 . Inside the bundlesheath cells, there is a greater concentration of CO2 and a lower concentration of oxygen than in parenchyma cells at the surface of the leaf. This difference in concentration allows CO2 to have a greater opportunity to bind with RuBP carboxylase. As a result, the plant can fix sufficient amounts of carbon to produce glucose in the Calvin cycle. In tropical climates, where the temperature often exceeds 28°C, food crops such as corn and sugarcane are commonly grown. Crops that use C3 fixation, however, do not survive well in tropical climates because they fix relatively less carbon and form fewer glucose molecules. Thus, the types of crops that can be grown in warmer climates are limited mainly to plants that use C4 carbon fixation. WEB LINK www.mcgrawhill.ca/links/biology12 Deserts plants, such as cacti and aloe vera, use a third method of carbon fixation called CAM (crassulacean-acid metabolism) fixation. Why do desert plants require a different method of carbon fixation? What processes are involved in CAM fixation? To find out more about CAM fixation, go to the web site above, and click on Web Links. Prepare an illustrated information handout that explains the process of carbon fixation in desertdwelling species of plants. Stage 2: Reduction In the second stage of the Calvin cycle, the stroma performs the necessary enzymatic reactions that reduce PGA to form PGAL. This happens in two stages. First, ATP molecules donate phosphate groups to the PGA molecules, converting them to bisphosphoglycerate, or PGAP molecules (see Figure 3.31). Secondly, an NADPH molecule, which was produced during the photo reactions, donates a hydrogen ion and two electrons to PGAP. This reduces PGAP to glyceraldehyde phosphate, or PGAL — the building block for anabolic processes including the synthesis of glucose. The oxidized NADP+ can return to the thylakoid membrane to be reduced again. ATP ADP + Pi PGA PGAP NADPH PGAL NADP + + H+ Figure 3.31 During the reduction stage, PGAP is reduced to become PGAL. CO2 phosphoenolpyruvate (PEP) oxaloacetate CCC CCCC AMP parenchyma cell ATP pyruvate malate CCC CCCC pyruvate malate CCC CCCC bundlesheath cell A CO2 Calvin cycle sugar B Figure 3.30 Wheat (A), and corn plants (B) have Figure 3.29 Carbon fixation in C4 plants adapted in different ways to their climates to circumvent the problem posed by active-site competition in the fixation of carbon dioxide. Chapter 3 Cellular Energy • MHR 91 Stage 3: Re-formation of RuBP Recall from Figure 3.27 and Figure 3.28 that RuBP, ribulose bisphosphate, is required in the carbon fixation stage of the Calvin cycle. RuBP is used to produce PGA, needed for the reduction stage of the cycle. Because PGAL is needed to reform RuBP, the majority of PGAL molecules, do not contribute to glucose production. The Calvin cycle reactions must occur twice to create one molecule of glucose. This is because for every three times that the Calvin cycle reactions occur, five PGAL are used to re-form three RuBP, ribulose bisphosphate, as shown in Figure 3.32. Notice from Figure 3.27 that 5 three-carbon PGALs contain the same number of carbon atoms as 3 five-carbon RuBPs. 5 PGAL 3 RuBP 3 ADP + 3 Pi 3 ATP Figure 3.32 Re-formation of RuBP. As five molecules of PGAL become three molecules of RuBP, three molecules of ATP become three molecules of ADP + Pi . To summarize the synthesis reactions of the Calvin cycle: ■ Stage 1: Carbon fixation, which takes carbon atoms from atmospheric carbon dioxide molecules and incorporates these atoms into organic molecules. ■ Stage 2: Reduction, which involves the formation of PGAP and its reduction to PGAL. ■ Stage 3: Re-formation of RuBP, which uses most of the PGAL molecules formed in the reduction stage to produce RuBP. This is then used to form more PGA in the Calvin cycle. Glucose: The Ultimate Food Source After glucose is produced in the synthesis reactions, plant cells can use glucose for glycolysis, followed by aerobic respiration in the mitochondria. The products and intermediary molecules of aerobic respiration provide the carbon-based molecules necessary to build amino acids, as well as the precursors to nucleic acids and lipids. However, there are many other ways that plants use glucose, for example, the conversion of glucose to starch, the formation of cellulose from glucose, and the conversion of glucose to sucrose. THINKING LAB Metabolic Rate and the Structure of Molecules O O O O O Background The molecule used by plants, such as corn and potatoes, to store energy is called starch. Starch is a large polymer composed of about 1000 glucose molecules. The starch is formed through condensation reactions, which link together individual glucose molecules. The starch molecule may contain side-branches, as shown in the illustration. Before a plant can use starch in aerobic cellular respiration, the starch must be broken down into individual glucose molecules. Recall that glucose is the molecule that first enters glycolysis. The rate at which glucose is available to be used in respiration can affect how quickly the cell will carry out metabolic processes. In other words, the availability of glucose can determine a plant’s metabolic rate. You Try It 1. What reaction is needed to break down a starch molecule into individual glucose molecules? 92 MHR • Unit 1 Metabolic Processes O O O O O O O CH2 O O O O O O O A starch molecule 2. If sweet corn contains mainly glucose molecules and starchy corn contains mainly starch molecules, in which type of corn would you expect cells to have a slower metabolic rate? Explain briefly. 3. Discuss how the structure of molecules might affect metabolic rate. 4. How could you determine if corn is sweet or starchy without tasting it? 5. What is the energy storage molecule in animal cells? These three ways will now be described. Autotrophs, such as green plants, produce a molecule used for energy storage, called starch. The starch is a large, branched polysaccharide composed of hundreds of glucose molecules linked by condensation reactions. Plants convert glucose to starch in the stroma. During peak hours of bright daylight, plants may produce more starch than they can use. This starch is stored in cells and is ready to be broken down into glucose for use in cellular processes. In the Thinking Lab on page 92, you will consider how the structure of starch can influence metabolic processes. In another series of reactions, plants may form another kind of polysaccharide that is the building block of cell walls — cellulose. PGAL is first exported from the chloroplast into the cytoplasm where condensation reactions take place to link glucose molecules. The formation of sucrose (the transport sugar in plants) also occurs in the cytoplasm. In order for glycolysis and cellular respiration to take place in the cytosol and mitochondria of plants, glucose is required. Because plants cannot move glucose molecules through the phloem (vascular tissue that transports organic material), they convert PGAL to glucose in the cytoplasm of leaf mesophyll cells. Glucose and fructose are then converted to sucrose. Sucrose is a molecule of fructose covalently bonded to a molecule of glucose. After sucrose is formed it is actively transported to the phloem, and then moved to locations in the plant that metabolize glucose. Photosynthesis Versus Aerobic Cellular Respiration Both plant and animal cells have mitochondria and carry out aerobic cellular respiration. However, only plants use photosynthesis. The cellular organelle for photosynthesis is the chloroplast, while the cellular organelle for aerobic cellular respiration is the mitochondrion. Figure 3.33 compares the processes of photosynthesis and respiration. Both processes have an electron transport chain located on membranes in the chloroplast and mitochondrion. ATP is produced on these membranes through the process of chemiosmosis. In photosynthesis, water is oxidized and oxygen is produced. In aerobic cellular respiration, oxygen is reduced to form water. Reactions in the chloroplast and mitochondrion are catalyzed by enzymes. These enzymes help to reduce CO2 to glucose in the chloroplast and oxidize glucose to CO2 in the mitochondrion. photosynthesis H2O aerobic cellular respiration O2 membranes ADP O2 ATP chloroplast NADPH CO2 H2O mitochondrion NADP + enzymes C16 H12 O6 NAD + C16 H12 O6 NADH CO2 Figure 3.33 Photosynthesis versus cellular respiration. How are products produced by one organelle used by the other organelle? The organelles shown are not drawn to scale, a chloroplast is about four times larger than a mitochondrion. Chapter 3 Cellular Energy • MHR 93 SECTION 1. 2. 94 K/U REVIEW Draw a chloroplast and label the key structures. K/U Explain how plants capture solar energy. Why does photosystem 680 provide some adaptive advantages over photosystem 700? 3. K/U Write an equation to summarize the photo reaction of photosynthesis. 4. K/U What are the components of a photosynthetic unit, and what roles do they play? In which part of the chloroplast are photosynthetic units located? 9. K/U How many CO2 molecules need to be fixed to produce 2 PGAL molecules that can leave the Calvin cycle to form glucose? 10. I In an experiment, a team of researchers uses a heavy isotope of oxygen-18 to track the passage of oxygen through the process of photosynthesis. What results would you expect to find if the researchers initiated the reaction in an environment in which (a) the carbon dioxide contained the heavy oxygen? (b) the water contained the heavy oxygen? 5. What role does water play in the photo reactions of photosynthesis? 6. K/U List the three stages of the Calvin cycle. For each stage, identify the energy input and the product(s). MC You are hired to advise strawberry producers on ways to increase their harvest. Assume the plants are grown under fully controlled conditions, so you can alter the temperature, lighting, atmosphere, water supply, and nutrients. 7. What becomes of the atmospheric carbon fixed by a plant cell? (a) What conditions would you recommend to maximize the plants’ productivity? 8. Explain, with diagrams, how high levels of oxygen reduce the effectiveness of the Calvin cycle. Explain how alternative forms of carbon fixation avoid this problem. K/U K/U C MHR • Unit 1 Metabolic Processes 11. (b) Can you think of any mutation that would help increase the rate or efficiency of photosynthesis? Discuss your reasoning.