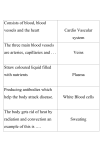
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
80 Central Nervous System Mechanisms Involving GABA Influence Arterial Pressure and Heart Rate in the Cat DANIEL J. WILLIFORD, BETTY L. HAMILTON, JANETTE DIAS SOUZA, THOMAS P. WILLIAMS, JOSEPH A. DIMICCO, AND RICHARD A. GILLIS SUMMARY Administration of the y-aminobutyric acid (GABA) receptor agonist, muscimol, into the third ventricle of anesthetized cats causes decreases in blood pressure and heart rate. To determine the brain areas involved, we administered muscimol into the (1) entire ventricular system, (2) lateral and 3rd ventricles, and (3) 4th ventricle. Muscimol 0.05-16.65 /ig administered into the entire ventricular system resulted in dose-dependent decreases in blood pressure, and at the two highest doses of 6.65 and 16.65 fig, significant reductions in heart rate. These changes in blood pressure and heart rate were mimicked when muscimol was localized to the 4th ventricle. Pressure and rate changes were not observed when muscimol was restricted to the lateral and 3rd ventricles. To determine the autonomic nerves involved in mediating the responses produced by muscimol administered into the 4th ventricle, bilateral vagotomy and bilateral stellate ganglionectomy were performed. Vagotomy had no effect on the responses evoked by muscimol, whereas stellate ganglionectomy prevented the decrease in heart rate without altering the effect of muscimol on blood pressure. To determine the efficacy of muscimol, the hypotensive and bradycardic effects of this agent were compared to those obtained with clonidine. The depressor response obtained with muscimol was greater than that seen with clonidine. In addition, muscimol produced an additional fall in pressure after a plateau effect had been obtained with clonidine; the converse was not observed. Pretreatment with the GABA receptor antagonist, bicuculline (25-100 pg) administered into the fourth ventricle in doses which had no effects on blood pressure or heart rate prevented the effect of muscimol. Treatment with bicuculline after a peak response had been obtained with muscimol restored pressure and rate to normal. These results indicate that activation of GABA receptors in the region of the hindbrain causes hypotension and bradycardia, and that both of these responses are mediated by a reduction in sympathetic outflow to the vasculature and heart. Circ Res 47: 80-88, 1980 Y-AMINOBUTYRIC ACID (GABA) may be an important neurotransmitter in central nervous system (CNS) pathways influencing activity in both divisions of the autonomic nervous system and thereby causing changes in cardiovascular function. Evidence for this is derived from studies using drugs that either interfere with or enhance CNS GABAergic mechanisms. Drugs known to interfere with the function of GABA, such as picrotoxin which inhibits the effect of GABA on chloride conductance (Ticku et al., 1978) and bicuculline which blocks the GABA receptor (Johnston, 1976), increase central sympathetic outflow and arterial pressure (DiMicco, 1978; Antonaccio et al., 1978a). These agents also increase parasympathetic outflow and decrease heart rate (DiMicco et al., 1977; DiMicco and Gillis, 1979; DiMicco et al., 1979). Similarly, a drug that inhibits the synthesis of From the Departments of Pharmacology and Anatomy, Georgetown University, Schools of Medicine and Dentistry, Washington, D.C. Supported by Grant NS-12566 from the U.S. Public Health Service and by a Grant-in-Aid from the American Heart Association with funds contributed in part by the American Heart Association, Nation's Capital Affiliate, Inc. Address for reprints: Daniel J. Williford, Department of Pharmacology, Schools of Medicine and Dentistry, Washington, D.C. 20007. Original manuscript received June 11, 1979; accepted for publication February 26, 1980. GABA in the CNS [thiosemicarbazide (Killam, 1957)] and an agent that has been found to block the release of GABA [tetanus toxin (Curtis et al., 1973)] increase sympathetic outflow (Taylor et al., 1977; Kerr et al., 1968). Conversely, substances that presumably act by directly stimulating GABA receptors (e.g., GABA and muscimol) decrease sympathetic outflow and arterial pressure and heart rate (DiMicco, 1978; Antonaccio and Taylor, 1977; Antonaccio et al., 1978b). In addition, representatives of a class of drugs thought to act in the CNS by increasing the affinity of the GABA receptor for GABA, the benzodiazepines (Costa, 1977), have been shown to decrease both sympathetic outflow and arterial pressure (Chai and Wang, 1966; Sigg et al., 1971; Antonaccio and Halley, 1975; Bolme and Fuxe, 1977). The purpose of the present study was to obtain more information on the influence of CNS GABAergic mechanisms in exerting control over arterial pressure and heart rate. Specifically, we used the selective GABA receptor agonist, muscimol, and determined: (1) where in the CNS muscimol was producing an effect that results in decreases in pressure and rate, (2) what division of the Downloaded from http://circres.ahajournals.org/ by guest on February 17, 2016 CNS GABA CONTROL OF CARDIOVASCULAR FUNCTION/ Williford et al. autonomic nervous system is involved in the decrease in pressure and rate induced by muscimol, (3) the efficacy of muscimol to lower pressure and rate as compared to the prototype CNS-active hypotensive agent, clonidine, and (4) whether or not the CNS actions of muscimol are a consequence of this agent interacting with GABA receptors. Methods Adult cats of both sexes and weighing from 1.9 to 3.3 kg were anesthetized with a-chloralose, 70-80 mg/kg, iv. A femoral artery and vein were cannulated for measurement of arterial pressure and systemic administration of drugs, respectively. The trachea was cannulated and all cats were artificially ventilated with room air. Animals then were immobilized with decamethonium bromide, 0.25 mg/ kg, iv, given every 45 minutes or as needed. Rectal temperature was monitored and maintained between 37° and 38°C by an infrared lamp. Blood pressure and lead II of the ECG were monitored continuously on a Grass polygraph. In some animals, the cervical vagus nerves were isolated and sectioned bilaterally. In addition, some cats were subjected to bilateral stellate ganglionectomy. This was accomplished by opening the chest through a lateral approach and removing both stellate ganglia. For intracerebroventricular administration of drugs, the cat's head was mounted in a David-Kopf stereotaxic apparatus. Three types of injections were performed in order to limit drug distribution. These were as follows: 1. for perfusion of the entire ventricular system, drugs were administered through a 26-gauge stainless steel spinal needle positioned in the left lateral ventricle at coordinates AP +11.5, HD +8.5, and RL +4.0, as indicated by the atlas of Snider and Neimer (1961). A short length of PE-160 tubing was inserted into the cisterna magna to provide an exit for displaced CSF. 2. for perfusion of the forebrain ventricles only (lateral and 3rd ventricles), injections were made through a needle placed in the left lateral ventricle as described above, with collection of perfusate through a cannula lodged in the Aqueduct of Sylvius, thereby preventing drug from reaching the 4th ventricle. 3. for perfusion of the 4th ventricle only, injections were made through a needle placed in the 4th ventricle at coordinates AP -8.5 HD —4.5 and RL 0 (Snider and Niemer, 1961). In all experiments, needle placement and drug distribution were confirmed by infusion of methylene blue dye and postmortem examination of the brain. All drugs infused intracerebroventricularly were dissolved in artificial cerebrospinal fluid (CSF) (Merlis, 1940, as modified by Feldberg et al., 1970) in a volume of 0.05 ml followed by a CSF flush of the same volume. Drugs administered systemically were dissolved in saline. The following drugs were used: a-chloralose (Es- 81 tablissements Kuhlmann), decamethonium bromide (Sigma Chemical Co.), muscimol (Biosearch), bicuculline methiodide (Pierce), clonidine HC1 (Boehringer Ingelheim), and y-aminobutyric acid (Sigma Chemical Co.). Statistical analysis was performed by using the grouped and paired £-tests, analysis of variance and Duncan's new multiple range test, and linear regression analysis by least squares. Criterion for statistical significance was p < 0.05. All values are reported as mean ± SEM. Results Effects of Muscimol Administered into the Left Lateral Ventricle on Arterial Pressure and Heart Rate According to Antonaccio and Taylor (1977), muscimol administered into the 3rd ventricle of anesthetized cats in doses ranging from 0.03 to 3.0 jug/kg produces dose-dependent reductions in arterial blood pressure and heart rate. In our study, a similar dose range of muscimol administered into the left lateral ventricle and allowed to perfuse the entire ventricular system of anesthetized cats produced a dose-dependent reduction in arterial pressure. Experiments were performed in seven animals and the data are illustrated graphically in the left side of Figure 1. Responses were observed with a dose as low as 0.05 ng and appeared maximal with a cumulative dose of 16.65 jug. Doses were treated in a cumulative fashion as the r'^ponse seen with each dose was maintained, and no evidence of recovery was observed over the duration of the observation period (30 minutes for the smaller doses and up to 2 hours for the larger doses). In terms of timeaction, arterial pressure began to fall within a minute or two after injection and the nadir was reached nearly always by 10 minutes after injection. Intervals between doses of muscimol therefore were approximately 10 minutes. The baseline mean blood pressures of these cats averaged 130 ± 6 mm Hg and the fall in pressure was to an average absolute value of 71 ± 8 mm Hg with the cumulative dose of 16.65 jug. Heart rate also was decreased by muscimol, but this was not reflected in the linear regression analysis of the data shown in the right side of Figure 1. However, using analysis of variance and Duncan's new multiple range test to analyze the data, a significant reduction in heart rate was observed with the two largest cumulative doses (6.65 and 16.65 jug). The baseline heart rates of these animals averaged 197 ± 10 beats/min and the fall in heart rate was to an average absolute value of 163 ± 5 beats/min with the cumulative dose of 16.65 /ig. The largest dose of muscimol accumulated in the intraventricular studies (16.65 jug) also was given as a single intravenous bolus injection to seven animals. By this route, muscimol produced no significant changes in arterial pressure or heart rate. Downloaded from http://circres.ahajournals.org/ by guest on February 17, 2016 82 CIRCULATION RESEARCH VOL. 47, No. 1, JULY 1980 +10 i - 1 -20 * -30 E -40 jS O "SO -60 -70 -801- 005 0.65 1.65 665 1665 - -80 I L 005 I 0.15 065 1.65 1 1 6.65 1665 Dose of Muscimol (/jg> 1 Changes in mean blood pressure (left panel) and heart rate (right panel) produced by muscimol administered in cumulative doses (indicated by the horizontal axis) into the entire ventricular system. Values represent the mean ± SEM of data obtained from seven cats. FIGURE Baseline values for arterial pressure and heart rate in these animals were 142 ± 7 mm Hg and 191 ± 7 beats/min, and at 10 minutes after injection of muscimol (the time at which the peak effect of muscimol was observed following intraventricular administration) the corresponding values were 138 ± 6 mm Hg and 189 ± 7 beats/min. Studies Designed to Localize the CNS Site of Action of Muscimol Two types of studies were performed and consisted of observing arterial pressure and heart rate changes following either (1) administration of muscimol into the left lateral ventricle and restricting the drug to the lateral and 3rd ventricles by cannulating the Aqueduct of Sylvius, or (2) administration of muscimol into the 4th ventricle. Data obtained from both types of studies are presented in Figure 2 and indicate significant dose-related decreases in pressure and rate when muscimol was administered into and restricted to the 4th ventricle, but no significant dose-related changes in pressure and rate when muscimol was administered into and restricted to the lateral and 3rd ventricles. The decrease in pressure and rate with the highest cumulative dose of muscimol administered into the 4th ventricle was similar to that seen when this dose was administered into the entire ventricular system. The absolute values attained were 84 ± 12 mm Hg (from 141 ± 10 mm Hg) and 154 ± 9 beats/ min (from 198 ± 16 beats/min). Peripheral Autonomic Nerves Involved in Mediating Pressure and Rate Changes Induced by Administration of Muscimol into the 4th Cerebroventricle The effect of bilateral vagotomy alone and vagotomy combined with bilateral stellate ganglionectomy on the heart rate and arterial pressure effects of muscimol were evaluated and the data are presented in Figure 3. As can be seen, bilateral vagotomy per se had no significant effect on responses produced by muscimol. Bilateral stellate ganglionectomy, however, prevented the dose-dependent decreases in heart rate normally observed when muscimol was administered into the 4th ventricle. This procedure, however, did not prevent the normally observed dose-dependent decreases in arterial pressure, although a slight attenuation of these responses was observed (see Fig. 3). Statistical analysis of the data revealed no significant differences between the hypotensive responses in cats with intact autonomic nerves, those with sectioned vagus nerves, or those with vagus nerves sectioned and stellate ganglia extirpated. Comparison of the Hypotensive and Bradycardic Effects of Muscimol with Those of Clonidine To obtain information on the potency and efficacy of muscimol administered into the 4th ventricle, we compared the depressor and bradycardic responses obtained with this agent to those ob- Downloaded from http://circres.ahajournals.org/ by guest on February 17, 2016 CNS GAB A CONTROL OF CARDIOVASCULAR FUNCTION/ WMiford et al. 83 +10,- 0- .E -10f5 % -20 <u 2: -30 -30 c -40 I O -50 -60 -60 -70 -70 -80 L 005 015 065 165 665 -SQL -L 005 1665 015 065 165 665 1660 Dose of Muscimol 2 Changes in mean blood pressure (left panel) and heart rate (right panel) produced by muscimol administered in cumulative doses (horizontal axis) into either the 4th ventricle (O) or into the lateral and 3rd ventricles (%). Values represent the mean ± SEM of data obtained from seven and six cats, respectively. FIGURE +10 +10 -KD — -20 -20 3 -30 -30 ro 55 -40 E -50 .£ -50 8, 2 6-60 O -60 -70 -70 -80 L- 005 OB 065 165 665 1665 -801- 005 015 065 165 665 1665 Dose of Muscimol (/Jg) 3 Effect of bilateral vagotomy (O) and vagotomy plus bilateral stellate ganglionectomy (9) on the changes in mean blood pressure (left panel) and heart rate (right panel) produced by cumulative doses (horizontal axis) of muscimol administered into the 4th ventricle. Values represent the mean ± SEM of data obtained from seven and four cats, respectively. FIGURE Downloaded from http://circres.ahajournals.org/ by guest on February 17, 2016 84 CIRCULATION RESEARCH +10 +10 r -10 -10 VOL. 47, No. 1, JULY 1980 -20 §5 £?-30 •|-30 £ 5,-40 Si 1-40 c 03 E-50 a,-60 £-60 I o -70 -70 -80 L 1.0 6.0 31.0 -80L IX) 31.0 60 Dose'of Clonidine <ug< FIGURE 4 Changes in mean blood pressure (left panel) and heart rate (right panel) produced by clonidine administered in cumulative doses (horizontal axis) into the 4th ventricle of vagotomized cats. Values represent the mean ± SEM of data obtained from eight cats. tained with clonidine, the prototype drug that acts primarily in the hindbrain to lower blood pressure and heart rate, (Schmidt, 1977). Clonidine was administered in three doses into the 4th ventricle of eight vagotomized cats, and the data are summarized in Figure 4. As can be seen, dose-dependent decreases in pressure and rate can be obtained with doses of 1, 6, and 31 fig of clonidine. The maximal effect of clonidine occurred with a cumulative dose of 31 jug (see Fig. 4). In five cats we observed that, once a maximal hypotensive effect had been obtained with clonidine, the additional administration of muscimol caused a further reduction in pressure of 18.4 ± 4.6 mm Hg (P < 0.05). When the converse experiment was performed, i.e., clonidine was administered into the 4th cerebroventricle of three animals in a dose of 25 jttg after a maximal hypotensive effect of muscimol had been attained, no additional decrease in arterial pressure occurred. Indeed, in all three experiments performed, only a pressor response occurred after the administration of clonidine. A comparison of the maximal effect produced by clonidine and muscimol on arterial pressure and heart rate when administered into the 4th ventricle is shown in Figure 5. The interesting finding was that muscimol produced a greater hypotensive effect than did clonidine. However, the heart rate effects of each agent were nearly equal. Role of GABAergic Mechanisms in Pressure and Rate Changes Induced by Administration of Muscimol into the 4th Ventricle Two types of experiments were performed to determine whether the hypotension and bradycardia produced by muscimol were caused by this Muscimol Clondre Muscimol Oomdne 5 Comparative effects of muscimol (seven cats) and clonidine (eight cats) on mean blood pressure (left panel) and heart rate (right panel). Control levels of pressure and rate are indicated by the unfilled histograms, and the responses of mean blood pressure and heart rate to cumulative dose regimens of muscimol (16.65 fig) and clonidine (31 fig) producing maximal responses are indicated by the cross-hatched histograms. FIGURE Downloaded from http://circres.ahajournals.org/ by guest on February 17, 2016 85 CNS GAB A CONTROL OF CARDIOVASCULAR FUNCTION/ Williford et al. Dose of GABA imgi 6 Peak changes in mean blood pressure (left panel) and heart rate (right panel) produced by GABA administered in increasing noncumulative doses into the 4th ventricle of seven vagotomized cats. Values represent the mean ± SEM. FIGURE agent activating GABA receptors in the hindbrain (i.e., in tissue in the vicinity of the 4th ventricle). The first type involved observing whether the neurotransmitter, GABA, could produce the hypotension and bradycardia seen with muscimol. GABA was administered into the 4th ventricle of seven cats subjected to bilateral vagotomy. Each animal received single doses of GABA consisting of 0.5, 2.5, and 10 mg. At these doses, GABA produced dosedependent reductions in pressure and rate (Fig. 6). These effects were transient (lasting from 10 to 30 minutes), and complete recovery to baseline values was attained before administering each subsequent dose. The second type of experiment involved testing the effect of the GABA receptor antagonist, bicuculline, on the hypotensive and bradycardic effects of muscimol. In some experiments, bicuculline was administered prior to muscimol to determine whether the responses could be prevented. In other experiments, bicuculline was administered after attaining the peak responses to muscimol to determine whether the responses could be counteracted. These experiments all were performed in animals subjected to bilateral vagotomy, and the drugs were restricted to the 4th ventricle. The data from the bicuculline pretreatment experiments appear in Figure 7. The magnitude of the decreases in pressure and rate with a single injection of 16.65 jug of muscimol given without bicuculline pretreatment is indicated from the vertical scales of the unfilled histograms in the figure. The cross-hatched histograms indicate the responses after administering 16.65 fig of muscimol to cats pretreated 10 minutes earlier with bicuculline in doses ranging from 25 to 100 /tg injected into the 4th ventricle. Bicuculline at these doses produced no significant changes in pressure or rate per se, but prevented the usual responses seen with 16.65 ng of muscimol. In the experiments in which bicuculline was tested for its capacity to reverse established responses obtained with muscimol, bicuculline in a dose of 25 fig was first administered into the 4th ventricle. Only slight and transient increases in pressure and rate were observed (see representative experiment depicted as Figure 8 and the unfilled 260 (- -1-80 -70 -70 •s 5220 60 X E 3-50 1 $-40 O m -30 c ,120 «3 <U 2 -10 o -20 ' -20 5-100 i t Muscrrol 2Spg FIGURE 7 Effect of bicuculline pretreatment on the hypotensive (left panel) and bradycardic responses (right panel) produced by muscimol in four animals. Standard errors of the mean are indicated on each histogram (see text for details); a = significantly different compared to muscimol alone. HO 0 10 20 30 ' 40 I I BMI Sa Musamd 50 _l 60 70 Time min FIGURE 8 Representative experiment depicting the interaction between muscimol and bicuculline (BMI) on heart rate (top panel) and mean blood pressure (bottom panel). (See text for details.) Downloaded from http://circres.ahajournals.org/ by guest on February 17, 2016 CIRCULATION RESEARCH 86 histogram of data summarized in Figure 9). After an interval of 30 minutes, muscimol in doses required to produce pronounced depressor and bradycardic responses (this varied from cat to cat and ranged from 2.5 to 16.65 fig) was administered (Fig. 8). Once responses reached the nadir and stabilized, 25 fig of bicuculline was readministered; this was observed to counteract both the hypotensive and bradycardic effects of muscimol (see Fig. 8). Readministration of muscimol abolished the antagonistic effect of bicuculline. The decreases in pressure and rate observed with muscimol were 73 ± 15 mm Hg and 44 ± 9 beats/min, respectively. Readministration of bicuculline produced increases of 64 ± 12 mm Hg and 38 ± 10 beats/min, as indicated by the cross-hatched histograms in Figure 9. Thus bicuculline administration completely reversed the hypotensive and bradycardic effects of muscimol. In addition, and as indicated by the histograms in Figure 9, bicuculline injected initially into the 4th ventricle produced only slight increases in pressure and rate, but when this agent was injected at a time when muscimol-induced changes in pressure and rate had occurred, pronounced rises in pressure and rate were observed. To test the selectivity of bicuculline to reverse muscimol-induced decreases in pressure and rate, bicuculline was administered to five vagotomized +80 r+70 - +70 +60 -<50 1 — cr i .30 +20 +20 +10 FIGURE 9 Effect of bicuculline treatment on the hypotensive and bradycardic responses produced by muscimol and clonidine. This figure illustrates the mean blood pressure (left panel) and heart rate effects (right panel) produced by bicuculline when administered alone (unfilled histograms—10 animals), after peak responses were obtained with muscimol (cross-hatched histograms—seven cats), and after peak responses were obtained with clonidine (filled histograms—five cats). Standard errors of the mean are also presented on each of the histograms. (See text for details.) a = Significantly different compared to changes produced by bicuculline alone; b = significantly different compared to changes produced by bicuculline in muscimol-pretreated group. VOL. 47, No. 1, JULY 1980 cats that had been given clonidine by injection into the 4th ventricle to lower pressure and rate. The depressor and bradycardic effects of clonidine (dose used was 31 fig) were 49 ± 7 mm Hg and 39 ± 13 beats/min, respectively. In contrast to what was observed with muscimol, bicuculline administration (25 jug) did not result in a reversal of these responses. Pressure increased by only 14 ± 4 mm Hg, whereas rate increased by 0.8 ± 0.6 beats/min. These values are depicted by the filled histograms in Figure 9. Discussion Muscimol administered into the entire ventricular system of chloralose-anesthetized cats decreased arterial pressure and heart rate. Effective doses ranged from 0.05 to 16.65 ng. This is not a new finding, as a similar dose range administered into the 3rd ventricle of chloralose-anesthetized cats had been reported by Antonaccio and Taylor (1977) to produce hypotension and bradycardia as well as dose-related reductions in renal sympathetic nerve discharge. Both studies indicated that these effects were due to a CNS rather than a peripheral site of action, because muscimol in doses that caused marked effects when administered by intracerebroventricular injection had no effect when given by the intravenous route. The new observation that has been made in the present study is that the site of action where muscimol causes reductions in pressure and rate is in the hindbrain (areas surrounding the 4th ventricle). We have excluded forebrain areas as a site of action by demonstrating that muscimol exerts no effect on arterial pressure and heart rate when restricted to the lateral and 3rd ventricles. It is not clear from our studies where muscimol acts in the hindbrain to produce these changes in pressure and rate. Our experiments also provide data indicating that muscimol produces hypotensive and bradycardic effects by activating GABA receptors in the hindbrain. This was demonstrated in the studies using bicuculline, an agent previously shown to act competitively to block the actions of muscimol and GABA at the receptor level (Johnston, 1976). Bicuculline given into the 4th ventricle in doses which had no significant pressure or rate effects prevented the hypotensive and bradycardic effects of muscimol. In addition, bicuculline administered into the 4th ventricle counteracted the fall in pressure and rate elicited by muscimol. The selectivity of action of bicuculline was shown by its inability to counteract hypotension and bradycardia produced by administration of a non-GABAmimetic drug, clonidine, into the 4th ventricle. Antonaccio and Taylor (1977) and Antonaccio et al. (1978b) also have used bicuculline to counteract the central cardiovascular actions of muscimol. However, the dose and method of administering bicuculline (0.5 mg/kg, iv, and 50 /xg/kg into the 3rd ventricle) results in significant pressure and rate effects per se (DiMicco and Gillis, Downloaded from http://circres.ahajournals.org/ by guest on February 17, 2016 CNS GAB A CONTROL OF CARDIOVASCULAR FUNCTION/ Williford et al. 1979). Hence the antagonism observed between bicuculline and muscimol in their study also could be interpreted as being due to a physiological interaction. That is, the fall in pressure and rate produced by one agent was being counteracted by the rise in pressure and rate induced by a second agent. The similarity between the CNS cardiovascular effects of muscimol and GABA also argue for a GABAergic mechanism being involved in decreases in pressure and rate evoked by muscimol. Both agents produced decreases in pressure and rate when administered into the 4th ventricle, and the arterial pressure effect was the most sensitive response produced by both substances. On the other hand, when clonidine was administered into the 4th ventricle, decreases in pressure and rate occurred in parallel. As pointed out by Antonaccio and Taylor (1977), muscimol is more potent than clonidine in producing centrally mediated decreases in pressure and rate. More importantly, it appears from our data that muscimol has greater efficacy in lowering pressure than does clonidine. This was shown by the larger reductions in pressure observed after muscimol as compared to clonidine, as well as by the depressor response observed with a single dose of muscimol after a plateau effect of clonidine had been attained. It should be pointed out that muscimol did not produce an additional slowing in heart rate after the peak effect of clonidine had been attained. This suggests that clonidine and muscimol cause a similar withdrawal of sympathetic tone to the heart but not to the vasculature. Both agents appear to have a relatively long duration of action, as the responses are maintained for at least 30 minutes and the doses have to be treated in a cumulative fashion. The division of the autonomic nervous system responsible for the bradycardic and hypotensive effect of CNS GABA receptor stimulation is the sympathetic nervous system. This was shown in the present study by noting the absence of any modifying effect on muscimol-induced bradycardia by bilateral vagotomy and abolition of the response by bilateral stellate ganglionectomy. Thus, cardiac slowing appears to be due entirely to withdrawal of sympathetic tone to the heart. Acute denervation of the heart however, had no significant effect on the hypotension produced by muscimol, suggesting that this response was due to withdrawal of sympathetic tone to the vasculature. Consistent with this suggestion are the earlier findings of Antonaccio and colleagues (Antonaccio and Taylor, 1977; Antonaccio et al., 1978b) that muscimol administered into the 3rd ventricle produced dose-dependent decreases in spontaneous discharge of renal sympathetic nerves. Finally, the data of the present study indicate that, whereas GABA receptor activation in the hindbrain produces pronounced decreases in pressure and rate presumably secondary to a reduction 87 in central sympathetic outflow, these receptors are not normally under the influence of GABA. This is indicated by the fact that administration of bicuculline into the 4th ventricle, in doses which blocked GABA receptors, had little or no effect on either pressure or rate. Thus, while a GABAergic mechanism is present in the hindbrain and may be considered as another approach to developing antihypertensive agents, this mechanism may not be operating under normal circumstances. References Antonaccio MJ, Halley J (1975) Inhibition of centrally-evoked pressor responses by diazepam: Evidence for an exclusively supramedullary action. Neuropharmacology 14: 649-657 Antonaccio MJ, Taylor DG (1977) Involvement of central GABA receptors in the regulation of blood pressure and heart rate of anesthetized cats. Eur J Pharmacol 46: 283-287 Antonaccio MJ, Kerwin L, Taylor DG (1978a) Effects of central GABA receptor agonism and antagonism on evoked diencephalic cardiovascular responses. Neuropharmacology 17: 597603 Antonaccio MJ, Kerwin L, Taylor DG (1978b) Reductions in blood pressure, heart rate and renal sympathetic nerve discharge in cats after the central administration of muscimol, a GABA agonist. Neuropharmacology 17: 783-791 Bolme P, Fuxe K (1977) Possible involvement of GABA mechanism in central cardiovascular and respiratory control. Studies on the interaction between diazepam, picrotoxin and clonidine. Med Biol 55: 301-309 Chai CY, Wang SC (1966) Cardiovascular actions of diazepam in the cat. J Pharmacol Exp Ther 154: 271-280 Costa E (1977) Some recent advances in the biochemical pharmacology of y-aminobutyric acid. In Neuroregulators and Psychiatric Disorders, edited by E Usdin, DA Hamburg, JD Barchas. New York, Oxford University Press, pp 372-380 Curtis DR, Felix D, Game CJA, McCulloch RM (1973) Tetanus toxin and the synaptic release of GABA. Brain Res 51: 358362 DiMicco JA (1978) Neurocardiovascular effects of the GABA antagonists picrotoxin and bicuculline in the cat: evidence for involvement of GABA in central cardiovascular control. Ph.D. Thesis, Georgetown University DiMicco JA, Gillis RA (1979) Neuro-cardiovascular effects produced by bicuculline in the cat. J Pharmacol Exp Ther 210: 1-6 DiMicco JA, Prestel T, Pearle DL, Gillis RA (1977) Mechanism of cardiovascular changes produced by activation of the central nervous system with picrotoxin. Circ Res 41: 446-451 DiMicco JA, Gale K, Hamilton BL, Gillis RA (1979) GABA receptor control of parasympathetic outflow to the heart: Characterization and brainstem localization. Science 204: 1106-1109 Feldberg W, Myers RD, Veale WL (1970) Perfusion from cerebral ventricle to cisterna magna in the unanesthetized cat. Effect of calcium on body temperature. J Physiol (Lond) 207: 403-416 Johnston GAR (1976) Physiologic pharmacology of GABA and its antagonists in the vertebrate nervous system. In GABA in Nervous System Function, edited by E Roberts, TH Chase, DB Towers. New York, Raven Press, pp 395-407 Kerr JH, Corbett JL, Prys-Robert C, Smith A, Spalding JMK (1968) Involvement of the sympathetic nervous system in tetanus. Lancet 2: 236-241 Killam KF (1957) Convulsant hydrazides. II: Comparison of electrical changes and enzyme inhibition induced by the administration of thiosemicarbazide. J Pharmacol Exp Ther 119: 263-271 Merlis JK (1940) The effect of changes in the calcium content of the cerebrospinal fluid on spinal reflex activity in the dog. Am J Physiol 131: 67-72 Downloaded from http://circres.ahajournals.org/ by guest on February 17, 2016 88 CIRCULATION RESEARCH Schmitt H (1977) The pharmacology of clonidine and related products. In Handbuch der experimentellen Pharmakologie, edited by F Gross, Springer-Verlag, Berlin-Heidleberg-New York, 39: pp. 299-396 Sigg EB, Keim KL, Kepner K (1971) Selective effect of diazepam on certain central sympathetic centers. Neuropharmacology 10: 621-629 Snider RS, Niemer WT (1961) A Stereotaxic Atlas of the Cat VOL. 47, No. 1, JULY 1980 Brain. Chicago, University of Chicago Press Taylor DG, Arbour KA, Antonaccio MJ (1977) Effects of bicuculline and thiosemicarbazide on inhibition of reflex and spontaneous sympathetic nerve potentials in the cat (abstr). Neurosci Abstr 3: 25 Ticku MK, Van Ness PC, Haycock JW, Levy WB, Olsen RW (1978) Dihydropicrotoxinin binding sites in rat brain: comparison to GABA receptors. Brain Res 150: 642-647 Instantaneous Femoral Artery PressureFlow Relations in Supine Anesthetized Dogs and the Effect of Unilateral Elevation of Femoral Venous Pressure WALTER EHRLICH, ROBERT W. BAER, RONALD F. BELLAMY, AND RANDALL RANDAZZO SUMMARY Instantaneous femoral artery pressure-flow (P/Q) relations were evaluated in consecutive 50-msec intervals of the pulseless flow changes during cardiac arrest in six anesthetized dogs and in two anesthetized dogs with a-adrenergic blockade. In all 245 P/Q graphs obtained under conditions of normal or elevated venous pressure, either with or without a blockade, the pressure-flow relations are linear, and the zero-flow intercept on the pressure axis—reached in less than 3 seconds after the onset of cardiac arrest—is markedly higher than the simultaneous venous pressure. We believe that the zero-flow intercept is the effective downstream pressure to arterial flow and that the reciprocal of the slope of the pressure-flow line indicates the arterial resistance. The elevation of femoral venous pressure raises the effective downstream pressure and the resistance to arterial flow in the same leg. The effective downstream pressure in the contralateral leg is raised also. a-Adrenergic blockade abolishes the reflex change in the contralateral leg, but a change in P/Q relations in the manipulated leg remains. We believe that the.central reflex change could be triggered by stretch receptors in the wall of the small veins transmitted to the arterioles through a-adrenergic receptors. The encroachment on the smallest arterioles by distended small veins and by the rise in interstitial fluid pressure might be the local mechanism by which venous pressure elevation directly changes arterial P/Q relations in the manipulated leg. CircRes 47: 88-98, 1980 IN 1933, Whittaker and Winton found in the isolated hindlimb of dogs linear pressure-flow relations with a positive zero-flow intercept on the pressure axis. In the intact hindlimb of dogs, Gomez and Veil (1936) observed that the femoral artery pressure fell during aortic occlusion to about 45 mm Hg. By increasing the sympathetic nervous tone, Pappenheimer and Maes (1942) shifted the projection of their straight femoral artery pressure-flow lines to higher pressures at zero-flow. Conversely, lowering sympathetic nervous tone shifted the projection of their lines to lower pressures at zero-flow values. From The Johns Hopkins University, Department of Psychiatry and Behavioral Sciences and Department of Environmental Health Sciences, Division of Physiology, Baltimore, Maryland and Walter Reed Army Institute of Research, Washington, D.C. Supported by Public Health Service Grant HL-10342. Address for reprints: Walter Ehrlich, M.D., The Johns Hopkins University, School of Hygiene and Public Health, Division of Environmental Physiology, 615 North Wolfe Street, Baltimore, Maryland 21205. Original manuscript received September 15, 1978; accepted for publication February 21, 1980. The femoral artery pressure-flow lines obtained by Green et al. (1944) also led to markedly positive zero-flow intercepts on the pressure axis, but the pressure-flow lines were convex to the pressure axis. Levy and Share (1953), Doyle (1953), and Levy et al. (1954) also obtained convex pressure-flow lines. These lines, however, point to a zero-flow intercept of about zero pressure. It is, of course, easy to dismiss the earlier reported linear pressure-flow relations and the positive zeroflow pressures with the argument that they were obtained with less advanced experimental technology than the convex pressure-flow relations with zero-flow intercepts at zero pressure obtained more recently. We believe, however, that the difference between the results of these two groups can be explained by the difference in experimental approach. Whittaker and Winton (1933) and Gomez and Veil (1936) followed the instantaneous fall of the femoral arterial pressure from continuous direct Downloaded from http://circres.ahajournals.org/ by guest on February 17, 2016 Central nervous system mechanisms involving GABA influence arterial pressure and heart rate in the cat. D J Williford, B L Hamilton, J D Souza, T P Williams, J A DiMicco and R A Gillis Circ Res. 1980;47:80-88 doi: 10.1161/01.RES.47.1.80 Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Copyright © 1980 American Heart Association, Inc. All rights reserved. Print ISSN: 0009-7330. Online ISSN: 1524-4571 The online version of this article, along with updated information and services, is located on the World Wide Web at: http://circres.ahajournals.org/content/47/1/80.citation Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which permission is being requested is located, click Request Permissions in the middle column of the Web page under Services. Further information about this process is available in the Permissions and Rights Question and Answer document. Reprints: Information about reprints can be found online at: http://www.lww.com/reprints Subscriptions: Information about subscribing to Circulation Research is online at: http://circres.ahajournals.org//subscriptions/ Downloaded from http://circres.ahajournals.org/ by guest on February 17, 2016