Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Nervous system network models wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Development of the nervous system wikipedia , lookup
End-plate potential wikipedia , lookup
Neuroanatomy wikipedia , lookup
Optogenetics wikipedia , lookup
Synaptic gating wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Embodied language processing wikipedia , lookup
Electromyography wikipedia , lookup
Caridoid escape reaction wikipedia , lookup
Muscle memory wikipedia , lookup
Proprioception wikipedia , lookup
Circumventricular organs wikipedia , lookup
Channelrhodopsin wikipedia , lookup
Premovement neuronal activity wikipedia , lookup
Neuromuscular junction wikipedia , lookup
Synaptogenesis wikipedia , lookup
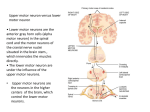
Commentary on slides for lecture 15 1. The muscles in your body are made up of muscle cells called myofibers each one formed by the fusion of muscle precursors cells, the myocytes. Each cell is thus multinucleated; as the contractile elements of the fiber form, the myonuclei are pushed to the outside of the cell. There are two major types of muscle fiber in the body: tonic and twitch. Some of the large muscles of the trunk are made up of tonic fibers. In these the degree of contraction is proportional to fiber depolarization; tonic fibers do not produce action potentials. The second kind of muscle fiber is the twitch fiber. When the twitch fiber is depolarized to threshold (via secretion of acetylycholine from the presynaptic terminal), an action potential is produced. This action potential invades an extensive system of intracellular membranes called the sarcoplasmic reticulum. Calcium rushes out of the SR (via voltage gated calcium channels) and causes the myosin and actin filaments to slide past one another and contract. Energy is required for muscle fibers to relax, hence rigor mortis. 2. An adult mammalian muscle fiber is innervated by one presynaptic terminal. If sufficient neurotransmitter is released the fiber will contract; if not, nothing happens. The all or nothing nature of the contraction of individual twitch muscle fibers produces a challenge for the production of smooth and coordinated movement by the central nervous system. This challenge is met in a variety of ways. First, the CNS can control how many motor units are active as a movement is being produced. The motor unit is all the muscle fibers that are innervated by a single motor neuron. Unit size can vary significantly from one or just a few fibers in the hand to hundreds or thousands of fibers in the large muscles of the thigh. 3. Individual muscle fibers are grouped together into bundles that make up the functional muscle of the body such as the gastroconemius of the leg. The muscles of the limbs are connected to the bones via tendons and, depending on their placement, can control different aspects of movement. For example, the sartorius muscle of the leg insery both into a bone of the thigh and a bone of the calf; it assistas in bending of the leg as for example when you sit cross legged and was named after tailors who used used to sit in this position. For example, we distinguish between leg extensors (that stretch the leg out) and leg flexors that abduct the leg. Both extension and flexion are caused by contraction of twitch muscle fibers; to produce a smooth movement is is often necessary to co-activate muscle that act in opposition. Muscles that produce the same sort of movement (extension or flexion) are called honomymous muscles while those that produce opposite effects are called heteronymous muscles (these are also known as synergists and antagonists, respectively). Muscle fibers that are next to each other in a particular muscle are innervated by motor neurons that are next to each other in the spinal cord. Motor pools (groups of motor neurons) that innervate adjacent muscles also lie next to each other in the spinal cord. There is thus a motor map in the CNS that closely resembles the somatotopic map that we saw in sensory systems. 4. To control movement effectively the CNS has to be provided with ongoing information about muscle length. This sort of information is particularly important, for example, when picking up an object of unknown weight. Two specific sensory systems provide this information to the CNS: one is an organ located within the muscle tendon called the Golgi tendon organ. The second is a specialized sensory structure within the muscle itself called the muscle spindle. The muscle spindle is an encapsulated structure that contains very fine muscle fibers, the intrafusal fibers, and the endings of neurons located in the dorsal root ganglia, the Ia afferent fibers. The intrafusal fibers are arranged in parallel with the main or extrafusal muscle fibers but do not contribute to the work done by the muscle. Instead, as we shall see, they help to maintain the sensitivity of the spindle over a wide range of muscle lengths. 5. Because the muscle spindle is sensitive to muscle length, it can be activated very effectively by a rapid muscle stretch such as that produced by hitting the fiber with a rubber mallet. The Ia afferents from a particular muscle fiber synapse on the motor neurons (alpha motor neurons) that innervate that muscle. The synapse is excitatory and can lead, for example, to leg extension as shown here. This kind of synaptic relation between Ia afferents and a particular muscle is preserved all the way up to motor cortex and is known as local sign. 6. The degree of contraction of the intrafusal fibers is controlled by a specialized group of motor neurons in the spinal cord called gamma motor neurons. 7. When the muscle fiber is stretched, the Ia afferent fibers are squeezed and fire a volley of action potentials. These excite the alpha motor neurons causing the muscle to contract. When the main muscle contracts, it shortens and the spindle is not longer stretched; we say that the spindle is unloaded. In this state it can no longer convey information about fiber length. However, the dynamic range of length over which stretch information can be provided can e increased if the gamma motor neurons fire at the same time as the alpha motor neurons; this scenario is called alpha-gamma co-activation. 8. The second sense organs related to muscle function is the Golgi tendon organ. This organ consists of the endings of dorsal root ganglion neurons called Ib fibers. These interdigitate with the braid-like strands of the tendon and fire when the muscle contracts and force is exerted on the tendon. The Gogi tendon organs help to convey information about the position of the limbs in space. In some diseased states the DRG neurons that provide muscle information die leaving the visual system as the sole source of information about limb position. These patients have severe disturbances in gait. 9. The spinal cord circuits that subserve sensory feedback to motor neurons are not just important because of their participation in simple reflexes such as the stretch reflex. These small circuits are the basic building blocks upon which larger aspects of the motor program are based and are used at all levels of motor control from central pattern generation through initiation of voluntary movement by motor cortex. 10. Two further levels of coordination at the level of the spinal cord deserve notice. First the action of synergistic and antagonistic muscle on one side of the body need to be coordinated in order to actually create movements. This coordination is produced by inhibitory interneurons (shown in black above) that are essential components of spinal reflex circuits. In the extensor reflex for example pain afferents from the foot excite flexors while, simultaneously, inhibiting ipsilateral extensors via Ia inhibitory interneurons. The second level of coordination is bilateral; coordination between the ipsilateral and contralateral halves of the spinal cord. In the crossed-extensor reflex shown here, the input from sensory afferents also excite extensor muscles on the opposite side of the spinal cord and inhibit, via inhibitory interneurons, the activity of flexor motor neurons. In diagramming these spinal circuits it is important to distinguish afferents from efferents as these terms describe the flow of information. In general the af- prefeix means “to” and the ef- prefix means from. Thus effernts from the ventral horn of the spinal cord (from the alpha motor neurons) provide innervation to the muscle fibers (they afferent the muscle fibers). Afferents from the DRG innervate neurons in various layers of the spinal cord itself. 11. The circuitry that underlies spinal reflexes is generally studied in cat spinal cord. A diagram of the experimental set up is shown here. Ia afferents from the flexor or extensor muscles can be stimulated separately via stetch and activation recorded extracellularly using en passant electrodes, The activity of motor neurons can be recorded with an intracellular electrode and the mono- or di-synaptic nature of afferent innervation distinguished by timing, production of IPSPs and EPSPs and pharmacology. 12. To finish our consideration of spinal circuits let us examine two in detail starting with the circuitry active under the influence of the Ia afferents from muscle spindles. When the muscle spindle is excited by stretch, Ia afferents excite flexor muscles via their motor neurons and inhibit extensor muscle via the excitation of a particular class of interneurons, the Ia inhibitory interneurons. This circuit can be activated locally (from the muscle) or by descending commands from higher levels of the CNS. Not shown in this diagram is the concurrent activation of gamma motor neurons that maintains sensitivity to stretch. 13. For Ib afferents, the nerve terminals of DRG cells insert into the “braids” of the tenson linking a particular muscle to the bone. When these braids are compressed, as the muscle contracts and pulls on them, action potentials are generated in the Ib fibers. 14. A dedicated circuit conveys information from the Ib afferents. One of the functions of this circuit is to limit muscle length and prevent tearing. Thus this circuit will, for example, will oppose activation of the flexor via inhibition (Ib inhibitory interneuron) and via activation of the extensor (Ib excitatory interneuron). 15. This circuit will also oppose activation of the flexor via activation of the extensor (Ib excitatory interneuron). 16. The final spinal reflex circuit we will consider involves a particular interneuron called the Renshaw cell. This interneuron is activated by axon collateral of alpha motor neurons. It serves to decrese the time that a motor neuron is excited. Renshaw cells are believed to sharpen up the pattern of excitation of motor neurons in a pool that are activated by descending input. This sharpening up involves the same kinds of circuits that lead to lateral inhibition in sensory systems. 17. Within the length of the spinal cord there is an orderly map representing all the muscle of the body. 18. The spinal cord is made of of a central region or grey matter that contains neuronal cell bodies surrounded by white matter, the myelinated axons that are travelling up and down the cord, entering the dorsal horn and leaving the ventral horn. Because there are more muscles in the arms and legs than in thoracic segments, the size of the grey matter is larger at these levels giving rise to the brachial and lumbar enlargements. Remember here the developmental mechanisms that contributes to these enlargements: a reduction of ontogenetic cell death in these regions associated with increased numbers of muscle targets. Within the spinal cord and the medulla there are networks of interneurons that contribute to rhythmic behaviors such as walking, breathing, and talking. These networks are called central pattern generators. 19. One of the jobs carried out by central pattern generators is to reconfigure combinations of motor neurons to produce different movements. Some idea of this reconfiguration can be gained from looking at movements of the fore and hindpaws of a cat as it walks, trots, paces and gallops. 20. The alternating activity of flexors and extensors has to be changed in order to go from walking to galloping. 21. How are these motor patterns produced? In theory, they could be activated sequentially by sensory feedback from muscle or cutaneous receptors. One way to examine this question is to record nerve activity during a particular pattern, such as walking, with and without (deafferentation) any sensory input. In the absence of sensory input the basic pattern is the same although the timing is not as tight. This kind of result gave rise to the idea that the basic circuitry for generating the motor program resides in the spinal cord and does not require sensory input. What kinds of circuitry could generate alternate activity, for example, in extensors or flexors. Here is an example taken from the flight muscles of locusts (elevator and depressor muscle that control wing beat). If excitatory input produced reciprocal inhibition, and if one excitatory interneuron leads the other, alternating bouts of activity in motor neurons could result from a constant barrage of activity to the excitatory interneurons. The input need not be symmetrical and involved excitation but could instead be the result of disinhibition. 24. A variety of models have been proposed to generate rhythmic behavior. One very common finding, however, is that in addition to participation of interneuronal circuits, there are substantial contributions from pacemaker neurons, neurons that have an endogenous firing rhythm (due, for example, to a leak current). The outputs of pacemakers neurons can be filtered by groups of interneurons to generate different rhythmic patterns. 25. These sorts of circuits have been worked out in detail in a number of different vertebrate and invertebrate systems. One set of examples are the swimming behaviors of sharks, lampreys and tadpoles. In swimming we also have to account for segmental coordination of muscle activity. 26. In the lamprey, Sven Grillner and his colleagues have outlined spinal cord circuits that can be activated by descending inputs from the brain stem or by sensory input from the skin.The circuits involve coordination across the midline using disinhibition. 27. Another very well understood motor system is the stomatogastric ganglion of lobsters and crabs. This is the ganglion that is responsible for powering the gastric mill that chops food items up. Only about 30 cells are involved and their wiring diagram is understood in its entirety. In this system different patterns of activity can be produced by neuromodulators that are capable of reconfiguring the activity of the entire network.