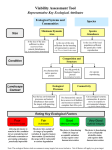
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Environmental change in the Kalahari: Integrated land degradation studies for non equilibrium dryland environments Andrew J. Dougill*, David S.G. Thomas1 and A.Louise Heathwaite * - The Environment Centre, University of Leeds, Leeds, LS2 9TJ, UK. - Sheffield Centre for International Drylands Research (SCIDR), Department of Geography, University of Sheffield, Sheffield, S10 2TN, UK. 1 - author for correspondence Recent decades have seen major intensification of cattle-based agricultural production in semiarid savanna ecosystems. In the Kalahari of Botswana, cattle production now occurs on privatized and fenced ranches. Patterns of ecological change, notably increased bush dominance, have been linked to increased cattle grazing intensity, but it remains contentious whether these changes represent land degradation. Uncertainty in ecological understanding stems from the dynamic, "non equilibrium" functioning of semiarid ecosystems. Given the inherent ecological variability of drylands, it is argued that degradation assessments should be based not on ecological observations alone, but on the study of long-term changes in pastoral production figures and on changes in the ecologically-determining factors of soil water and soil nutrient availability. Provided here is a framework incorporating soil and ecological changes at a range of scales that can enable us to distinguish drought-induced fluctuations from long-term ecological state changes. The results demonstrate that increased cattle use and associated ecological changes have not been caused by, nor are associated with, changes in soil water and nutrient availability. A model of ecosystem dynamics is presented that displays bush encroachment not as a definite form of land degradation. Encroachment may also be curtailed by resilience mechanisms found in protected ecological niches and also by the resilience of the nutrient poor sandy soils. Key words: Land degradation, Dryland pastoralism, Ecological modeling, Resilience, Bush encroachment, Soil hydrochemistry, Kalahari Issues of contemporary environmental change in dryland areas, and in particular their association with desertification, remain controversial in many respects due to their multi-faceted social and environmental dimensions. Desertification does, however, remain high on the global environmental agenda, as witnessed through the ratification of the United Nations Convention to Combat Desertification (CCD), and continued global efforts to assess the extent of desertification (Middleton and Thomas 1997). The CCD defines desertification as "land degradation in arid, semiarid and dry subhumid areas resulting from various factors, including climatic variations and human activities" (Middleton and Thomas 1997:viii). In this context, land includes the soil and water resources, as well as local vegetation cover upon which rural agricultural livelihoods are dependent. Environmental studies therefore need to be wide-ranging in their remit to produce a clear understanding of the nature and extent of desertification problems. The term degradation implies a reduction of the resource potential by one or a number of processes operating on the land (UNEP 1997). These definitions provide the context for contemporary environmental studies in drylands, implying that changes in the natural resource base of vegetation and soil systems can only be classified as desertification if they reduce the land’s value to human systems. One study has defined desertification in this context as "an effectively permanent decline in the rate at which land yields agricultural products under a given management system" (Abel and Blaikie 1989: 113). Conventional media images showing desertification as extensive wind or water erosion of soils serve to over-simplify and often confuse the most important environmental and social issues. Much academic debate has focused on how these images have exaggerated both the physical extent and the social impacts of dryland degradation, leading to the alternative view that desertification is an institutional myth (Warren and Agnew 1988, Hellden 1991, Thomas and Middleton 1994). Less debate, concern, and publicity has been afforded to changes in the dryland ecological resource-base (the savanna vegetation communities) which are less overt, and yet are responsible for much of the reduced agricultural potential, especially in areas used for intensive cattle ranching. The most notable example of this, and that investigated here for the Kalahari of southern Africa, is that of bush encroachment, the ecological transition to a state of increased bush dominance. This transition reduces grass cover in the landscape and can reduce economic outputs from grazing-based farming systems (de Ridder and Breman 1993, Quan et al. 1994), as both the total fodder availability and its heterogeneity are reduced (Scoones 1995). Bush encroachment has now become recognized as a threat to sustainable pastoralism in all major savanna regions throughout the globe. Indeed, Warren and Agnew (1988: 6) state "it has often been maintained that the most widespread problem on (dryland) pastures is not their complete degradation but their invasion by thorn scrub." Previous environmental studies of this phenomenon have implicitly linked it to the intensity of cattle grazing, with bush encroachment being associated with areas of intensive cattle use (e.g., Buffington and Herbel 1965, van Vegten 1983, Andrew 1988, Perkins and Thomas 1993a,1993b). Increased awareness of the existence of bush encroachment has not, however, been paralleled by advancements of our understanding of the underlying processes of change in relation to the controlling factors of rainfall variability, changes in fire regimes, the direct impacts of cattle grazing, nor the interrelationships between these factors. Yet without such environmental understanding, preventative and control measures aimed at reducing bush encroachment will remain difficult to provide. A number of different theoretical models have been proposed to explain the transition to greater bush dominance in savannas (e.g., Walker and Noy-Meir 1982, Westoby et al. 1989, Schlesinger et al. 1990). This paper starts by outlining differences in these ecological change models before assessing their applicability to, and implications for, privately owned cattle ranches on the flat extensive sandy plains of the Kalahari. The wider significance of these studies to other semiarid regions with similar environmental and land use characteristics, such as in Australia (Andrew 1988, Pickup 1995), Sahelian Africa (Warren and Khoghali 1992), and the southwest USA (Schlesinger et al. 1990, Archer 1990) are highlighted to demonstrate the wider applicability and complexity of current geographical debates associated with changes in the dryland ecological resource base. These debates are extended to examine the future role, and suggested methodologies for, integrated environmental monitoring of changes in soil and vegetation systems aimed at land degradation assessments, and for improved incorporation of environmental studies into the development of sustainable land use practices. Ecological change models Theoretical discussions about the causes of bush encroachment have developed as part of the rapid evolution of dryland research and the recent paradigm shift recognizing the "nonequilibrium" nature of ecological functioning (Behnke et al. 1993, Scoones 1995, Warren 1995). Due to the inherent rainfall variability and patchy fire regimes of drylands, there is now a recognized need to place short-term environmental studies within the context of the natural environmental variability that characterizes such areas. The environmental paradigm shift has important implications for determining what constitutes degradation and for models attempting to explain the interrelated causes of ecological transitions. Ecological change models require adaptation to incorporate the effects of variability in rainfall and fire regimes, factors which now form the focus of dryland ecological studies. In this section we discuss current understandings of the causes of bush encroachment in drylands, and detail the more comprehensive integrated studies needed to adapt environmental studies to the dynamic, highly variable nature of nonequilibrium theories. Conventional models derived to explain the processes leading to bush encroachment can be divided into two main groups: first, those that suggest bush encroachment occurs solely due to changes in grazing and fire regimes acting independently from soil factors (e.g., Westoby et al. 1989), and second, those that act through changes in the soil resource (e.g., Walker and NoyMeir 1982, Schlesinger et al. 1990). Given the ability for agricultural land use practices to modify grazing and fire regimes in relation to the environmental conditions experienced at a given time and place, this distinction has important implications for the provision of sustainable natural resource management strategies. Such adaptations have been termed "opportunistic management" (Westoby et al. 1989) and have been heralded as a flexible and sustainable way to manage natural resources through coordination between land use planners and local land managers (Scoones 1995). Adaptations such as rapid destocking in times of drought to prevent grass mortality (Mott et al. 1992) and the use of wet season fires to reduce bush dominance (Hodgkinson et al. 1984) will only be successful in controlling bush encroachment if it is these pressures alone that are the causal agents. The aim and importance of soil-based studies, on the other hand, is to determine whether bush encroachment occurs at least partly because of changes in the soil resource, including the depth profile and spatial distributions of soil water and nutrient availability (Walker and NoyMeir 1982, Schlesinger et al. 1990). If the ecological distribution of bushes has a soil hydrochemical causation independent of changes in the grazing and fire regimes, it suggests that the ecological dominance of bushes can remain in the landscape, then bush encroachment represents a permanent decline in the resource base for pastoralism. This is especially true given the findings of longer term ecological studies showing that bush species can recover rapidly from disturbances that cause die-back (Whitford et al. 1995). Consequently, the recent shift in emphasis of dryland degradation studies to consider the impact of management strategies on soil factors (e.g., Parr et al. 1990, Oldeman et al. 1990, Stocking 1995, Middleton and Thomas 1997) is important, and provides a context for the research reported here. This article builds on previous studies by moving away from investigating visible attributes of soil degradation to consider patterns of soil water and nutrient availability. Importantly, it is these factors that are recognized as the primary controls on the productivity and vegetation community structure (i.e., the ratio of bush to grass cover) within dryland savanna ecosystems (Scholes and Walker 1993). In this respect the "two-layer model of environmental change” (Walker and Noy-Meir 1982) has been widely used as the explaination of bush encroachment in the Kalahari (Skarpe 1990, 1991, Perkins and Thomas 1993a, 1993b) and in other regions. This model is now described, before moving to consider recent ecological developments that are important in land degradation debates. The two layer model In this model, the ecological balance between bush and grass production is determined by the relative availability of soil water and key nutrients (principally nitrogen (N) and phosphorus (P)) in different rooting zones in the soil profile. Ecologically, grasses out-compete bush species for water and nutrients in the topsoil (0 - 50 cm depth), while bush species have the competitive advantage in the subsoil below 50 cm (Walker et al. 1981, Belsky 1990). According to the two layer model, cattle grazing effects the bush:grass ratio by suppressing grass growth, and therefore topsoil plant water uptake, promoting soil water movement into the subsoil. At the same time, the increased mineralization of organic nitrogen inputs into the water soluble inorganic form of nitrate (NO3--N), caused by the fact that the surface inputs of cattle dung are more readily decomposed than residual plant litter, enhances leaching of this vital plant nutrient into the subsoil. Consequently, the two layer model predicts that areas of intensive grazing experience significant increases in moisture content and "plant-available" inorganic nutrient concentrations in their subsoil layers (Figure 1), with bush encroachment being the ecological consequence. The model implies that bush encroachment is linked to major soil hydrochemical changes that favor bush dominance into the future, irrespective of changes in grazing and fire regimes. However, changes in the nutrient mineralization and leaching processes that the model is dependent upon have not been observed in controlled laboratory or field process-based studies on Kalahari sand soils (Dougill et al. 1998a) leaving uncertainty about the actual extent of the soil-to-ecological-change links proposed by the two-layer model. Research in this paper examines the links between patterns of ecological change and soil profile hydrochemical factors through ranch scale measurements at sites experiencing different grazing intensities and ecological community structures. New ecological theories and land degradation assessments "Non-equilibrium" ecological theories are founded on the identification of the inherent dynamism of dryland ecosystems, where rainfall plays a greater role in plant growth than variations in grazing regimes (Holling 1973, Wiens 1977, Sandford 1983, Ellis and Swift 1988, Friedel et al. 1993). Vegetation changes are often reversible, such that even systems classified as severely degraded demonstrate rapid recovery characteristics, especially following the return of good rainfall after drought years (Tucker et al. 1991, Perkins and Thomas 1993a). Such studies suggest that semiarid ecosystems are well adapted to cope with, and to respond to, disturbance, due partly to the continuous natural (rainfall and fire) disturbances upon which their original community composition is dependent. This recent view of dryland ecosystems as resilient systems directly opposes the traditional representation of semiarid ecosystems as fragile "degradation prone" environments (Stoddart et al. 1975, Kharin 1990). Rather than grazing disturbances being automatically seen as necessarily increasing system instability and enhancing the risk of desertification, more integrated understanding of the causes and permanence of transitions between vegetation states is required. In particular, environmental monitoring of changes in vegetation and soil resources is essential to distinguish between drought-induced ecological fluctuations and permanent changes in vegetation community state. Non-equilibrium theory clearly dismisses the applicability to dryland savannas of traditional "climax and equilibrium" ecosystem models (Behnke et al. 1993, Warren 1995), necessitating alternative models of ecosystem functioning for these areas. Westoby et al. (1989) have formulated these, as "state and transition" models. These models suggest that dryland ecosystem dynamics can be described by a set of discrete states of the vegetation community, and a series of distinct transitions between states. State and transition models imply that environmental changes can occur very rapidly and may be triggered either by management actions, such as increased cattle stocking levels or managed burning, and/or by natural events including rainfall variability and fire. Such models therefore better represent the natural complexity of interrelations experienced in drylands, but they pose difficulties for the environmental scientist attempting to maintain clarity and simplicity in degradation debates, as well as for land managers seeking to intervene in the land use decision making process (Thomas 1997). It is, however, essential that the exact conditions and processes leading to bush encroachment in particular environments be fully understood so as to prevent declines in agricultural productivity. The acceptance of non-equilibrium ecological theory has a number of important implications for limited time-scale and small spatial scale studies that typify environmental investigations used to assess the existence of land degradation in dryland areas (Middleton and Thomas 1997). The main implications are summarized in Table 1, which details the implications of theories in terms of the need to move away from degradation assessments based solely on vegetation community changes, and also the specific implications for the findings of previous Kalaharibased environmental studies. Non equilibrium concepts need to influence the manner in which contemporary environmental studies are designed, with there being an increased focus on understanding the interrelated effects of rainfall, grazing, and fire regimes in affecting the ecological and soil resource bases. As Table 1 describes, this can be achieved by extending the time frame of ecological studies, by incorporating clear recognition of different spatial patterns of ecological change in relation to patterns in the driving forces of grazing, rainfall, and fire regimes, and/or by assessing the impact of grazing on soil hydrochemical factors. This paper represents a case study detailing how these implications have been incorporated into integrated environmental studies, in this case for eastern Kalahari ranches in Botswana. The Kalahari The Kalahari desert, thirstland, or sandveld, as it has variously been termed, can be defined in a number of ways using a range of criteria (Thomas and Shaw 1991). Here it is designated as the area covered by deep Kalahari Sand deposits, centered upon western and central Botswana but extending into neighboring countries (Figure 2). It is characterized by semiarid climatic conditions (Bhalotra 1987) and typified by very limited relief. Geologically, the Kalahari is an extensive basin infilled with nutrient deficient aeolian sediments (Thomas and Shaw 1990). Kalahari soils are therefore predominantly deep, structureless, fine sands, low in essential plant nutrients (Skarpe and Bergström 1986, Buckley et al. 1987a, 1987b) and with a low organic matter content (Perkins and Thomas 1993b). The high infiltration rates afforded by the sandy sediments, together with the lack of perennial rivers, make the Kalahari a location where natural water availability is limited to small short-lived accumulations in occasional pan depressions. It is this lack of surface water, not the climatic characteristics, that lead to its portrayal as a desert. Despite the absence of surface water, the mixed grass and shrub savanna (Weare and Yalala 1971) have resulted in the view that the Kalahari is an untapped grazing resource (Debenham 1952). Pastoral utilization of this ecological resource base has intensified since the 1950s as a result of the large-scale sinking of deep boreholes to exploit ancient groundwater reserves in the underlying sandstone aquifers (de Vries and von Hoyer 1988). The nature of this intensifying development program has been clearly linked to a series of national policy-driven structural changes in the agricultural economy. Botswanan agricultural development Since independence in 1966, the government of Botswana, supported by a range of international development aid and trade agreements, has voraciously pursued policies of expanding privatized "ranch-style" livestock production in the Kalahari. Successive agricultural policies have aimed to relieve pressure on the country’s more densely populated eastern hardveld, where land degradation caused by high livestock numbers is perceived to have caused environmental damage (see Abel and Blaikie 1989 and Biot 1993 for discussion). The first Livestock Development Project (LDP1), begun in 1970, established forty ranches in the Kalahari as a means of encouraging herd owners to commercialize their activities (Odell 1980). In 1975, the LDP1 was subsumed under the wide-ranging Tribal Grazing Lands Policy (TGLP), which aimed to enhance national economic and social development by reorganizing Botswana's livestock industry. In so doing, it was hoped to both improve the livelihoods of rural dwellers and to reduce the environmental degradation attributed to "overstocking" under the traditional communal-grazing systems in the eastern hardveld. Development policies in Botswana have matched those in many other parts of dryland Africa (Lane 1998) by using the well publicized principle of the "Tragedy of the Commons" (Hardin 1968) to fence off and privatize large portions of once communal land. The TGLP aimed to expand cattle production in the Kalahari through the creation of fenced leasehold ranches of a uniform size, each supplied with groundwater via a borehole. Ranches were to be leased from the state by cattle owners or syndicates who owned over 400 animals. Lessees would gain sole water rights for the groundwater boreholes and therefore de facto control of grazing and ecological resources within a ranch. It was hoped that by moving large livestock owners off communal lands, overgrazing would be reduced, giving the smaller farmers who remained the opportunity to improve their livelihoods. Overall, therefore, the TGLP was intended to alleviate environmental and social problems in one part of the country by encouraging development elsewhere. The social applicability of these policies have now been widely criticized (Campbell et al. 1991, Tsimako 1991, Peters 1995, Thomas and Sporton 1997), due largely to the inapplicability of assumptions underlying the TGLP. These included the view that the Kalahari was an uninhabited wilderness and that, with the allocation of private ranches, large cattle owners would no longer graze their cattle on communal lands. In reality, many designated areas were on lands used by rural peoples for seasonal grazing for over 200 years (Campbell et al. 1991, Moyo et al. 1993). In addition, ranch lessees have continued to use their dual grazing rights on both private and communal lands, thereby exacerbating social inequalities in terms of access to grazing land and livestock (Good 1992, Peters 1995). The social implications of the TGLP therefore include reduced status and increased poverty among rural dwellers (Thomas and Sporton 1997). What remains less clear is the extent and permanence of ecological changes on TGLP ranch blocks and consequently the long term environmental sustainability of the ranching practices promoted. It is these factors which form the focus of the integrated environmental study reported here. The framework for expansion of privatized ranches provided by the TGLP was strengthened further by the 1991 National Policy on Agricultural Development (Republic of Botswana 1991). This called for further increases in livestock productivity through improved management, and continued commercialization, the establishment of additional ranches and, significantly, the fencing of all communal lands. Given the uncertain environmental consequences of this move to complete privatization, it is vitally important to assess the environmental impacts of the 25 years of intensive grazing experienced on TGLP ranches. The changes seen on these ranches will act as an indicator of the threats to sustainable natural resource management faced in the future, further privatized, Kalahari. Study site: The Makoba ranches One of the original blocks of leasehold ranches, established by the TGLP in 1975, is the Makoba ranch block, in the eastern sandveld of the Central District (Figure 2). This area has a mean annual rainfall of approximately 400 mm. The inter-annual variability of over 35 percent (Bhalotra 1987) greatly regulates ecological changes. At the time the ranches were designated, vegetation communities consisted of mixed savanna with grassy plains dominated by tufted perennial grasses, including Eragrostis, Pogontharia and Stipagrostis species, and low shrubs, commonly of Acacia, Lonchocarpus, Grewia, Rhigozum, and Terminalia species (Perkins and Thomas 1993b). Each ranch covers a standard area of 6400 hectares and was initially served by a single, diesel-pumped, borehole obtaining groundwater from over 60 meters depth. Up until the early 1990s, the ranches were indistinguishable in their operation from traditional “cattleposts,” with the absence of any paddocks or perimeter fencing, implying cattle were free to roam throughout the rangeland surrounding the borehole. Therefore, ecological studies that began on the Makoba ranches in 1989 (Perkins and Thomas 1993a, 1993b) initially assessed the impacts of the shift to a more intensively grazed cattle-based system, in which the single borehole is effectively "the herder" (Jerve 1982). Given the increasing available grazing area with distance from a point water source grazing intensity therefore declines exponentially in all directions away from the borehole. This pattern of grazing intensity decline enables direct study of the relationship between grazing intensity and changes to soil or vegetation resource bases, a technique termed the "piosphere" approach (Georgiadis 1987, Andrew 1988). Ongoing studies throughout the 1990s have taken place through a period of further structural changes, in line with the aims of the National Policy on Agricultural Development in the Makoba ranches. This has seen further boreholes sunk on ranches and the partitioning of ranches into paddocks that are then used for rotational grazing in a manner similar to conventional western-world ranching schemes. One such ranch, Uwe Aboo, has been the site of detailed environmental studies throughout the 1990s. Uwe Aboo has seen two new boreholes drilled and the ranch has been partitioned into five paddocks (Figure 3) since the initial vegetation surveys of Perkins and Thomas (1993a). These additional development pressures and the spatial and temporal intensification of cattle grazing they cause need to be considered throughout our discussion of the results from our environmental studyof TGLP ranches. Integrated ecological and soil studies To account for the natural variability of dryland environmental systems, and to attempt to differentiate between drought-induced ecological changes and agriculturally-determined degradation, conventional, limited time-scale environmental studies (Table 1) need to investigate both the spatial patterns of vegetation community changes in relation to agricultural pressures, and the nature of changes in soil hydrochemical characteristics that fundamentally determine ecosystem structure and productivity. To achieve these dual objectives, our research in the Makoba ranches has integrated ecological ecological (Perkins and Thomas 1993a,1993b, Dougill et al. 1998b) and soil-based (Dougill et al. 1997, 1998a) monitoring on a variety of scales, ranging from the ranch scale of the 6400 ha blocks to the fine scale of localized bush and grass patterns and processes of water and nutrient movement in individual soil profiles (Table 2). The findings of the array of studies are synthesized here to highlight the information available through the integrated use of multi-source and multi-scale environmental information, and to provide a context for the development of an alternative model of bush encroachment. It is therefore first necessary to consider the results of ongoing ecological studies examining the spatial patterns of vegetation community changes, before considering the results of the soil and water study. Ecological change on TGLP ranches Initial ecological studies by Perkins and Thomas (1993a,1993b) demonstrated that vegetation community changes are clearly associated with grazing patterns on Makoba ranches they investigated. Changes in vegetation community structure, measured using the surrogates of bush cover and the bush:grass ratio, were directly correlated to the gradient of grazing impact decline away from a borehole. Spatially, the most extensive ecological change is that of bush encroachment, extending out to a distance of up to 2 km from a borehole, with the pattern appearing most expansive on the older boreholes (Perkins and Thomas 1993a). Given the initial adoption of only one borehole per 8 km square ranch within the TGLP, the limited spatial extent of bush encroachment ensured that substantial areas remained largely unaffected by such ecological changes. This "unencroached" zone remains largely grass-dominated, and as such has been termed the "grazing reserve" (Perkins and Thomas 1993a). Field-based ecological evidence suggestis that a "bush encroached grazing reserve" duality is the major ecological pattern on TGLP ranches, a view supported by preliminary satellite remotesensing studies (Ringrose et al. 1990, 1995, Trodd and Dougill 1998). Consequently, on a typical 6400 ha ranch block with a single borehole, ecological and fodder diversity remains high within the ranch. It is the maintenance of this fodder diversity, in terms of both bush and grass cover, which can explain the economic observations that, as yet, ecological changes have not significantly reduced pastoral production (Vossen 1990, White 1993). Indeed, bush cover provides an important fodder resource that can maintain herd sizes through times of drought and enable rapid restocking when good rains return. This economic viewpoint matches the current focus of the degradation definitions detailed earlier, but conclusions need to be carefully placed within the context of the continued environmental changes, structural reforms and grazing intensification on TGLP ranches. The recognition of bush encroachment as an expansive process and the relaxation of the 8 km spacing between boreholes (Tsimako 1991) implies that there is both an ecological and management potential for the coalescence of bush dominant areas. It is the formation of such bush-dominated landscapes that has significantly reduced agricultural outputs in other dryland areas, and would constitute degradation, whether environmentally or economically defined. It is this threat of ecological changes representing degradation in the future that implies that investigations of the physical mechanisms leading to bush encroachment, and ecological resilience mechanisms retaining grass cover in grazed rangelands, need to be understood more clearly. Bush encroachment in intensively grazed areas not only increases the ratio of bush to grass production, but typically increases the proportion of certain invading bush species. Notably, fine leaved Acacia species, Dichrostachys cinerea, and Terminalia sericia have been identified as the main encroachers on the Makoba ranches (Perkins and Thomas 1993a). At Uwe Aboo ranch, bush encroachment consists predominantly of an increase in the cover of Acacia mellifera and Acacia ataxacantha, at the expense of broad-leafed species of Lonchorcarpus nelsii and Grewia flava, which dominate unencroached areas. Associated changes in the composition of herbaceous species also occur, with increases in unpalatable species such as Perotis patens and Tragus berteronianus at intensively grazed sites, but also with certain more palatable species including Digitaria eriantha and Brachiara nigropedata (Perkins and Thomas 1993a). Explanation of the maintenance of nutritious grasses, even in intensively grazed areas, requires the analysis of fine-scale spatial patterns of bush and grass cover to examine controls in regenerative ecology (Vetaas 1992). Incorporation of studies at Uwe Aboo quantifying the detailed grass and bush patterns within 10 m2 quadrats at sites experiencing different grazing intensities (see Dougill et al. 1998b for methodological details and Figure 3 for site locations) matches the shift in applied ecological studies seen in many other savannas (e.g., Frost and McDougald 1989, Menaut et al. 1990, Belsky et al. 1993a, 1993b, Belsky 1994, Amundson et al. 1995). Fine scale ecological studies in the Kalahari (Dougill et al. 1998b) show that the role of sub bush canopy sites is vital in imparting ecological resilience. This is similar to the results of studies in East Africa (Belsky et al. 1989, 1993a, 1993b) and Southwest US (Archer 1990). At the end of a dry season, and despite competition from bushes for soil water and nutrients, subbush canopy sites are typified by significantly greater grass biomass than in neighboring "open" sites (Dougill et al. 1998b). High sub-bush canopy grass biomass is particularly associated with low growing thorny Acacia species that are the main encroachers on this ranch. Consequently, bush encroachment is not associated with the complete removal of grass cover, as grazing protected sub-bush canopy niches retain a grass seed resource of nutritious species. This seed resource maintains the ecological potential for the transition back to a state of grass dominance following disturbances causing bush die-back, whether these occur through natural events such as drought or intense fires, or through management actions such as burning or manual bush clearance. The ecological potential for a transition back to a state of grass dominance still depends, however, on rainfall and soil hydrochemical characteristics that determine the competitive dominance of different species (Scholes and Walker 1993). It is the remaining uncertainty over the relative importance of, and interrelations between, controlling factors of ecological transitions that underlies the need to extend environmental studies to include longer term vegetation monitoring using remotely-sensed satellite data (Trodd and Dougill 1998) and soil hydrochemical studies, such as those reported below. Soil system studies: Applicability of the two-layer model Soil-based studies investigating the links between grazing intensity, ecological change, and soil hydrochemical characteristics on the Uwe Aboo ranch have focused on both changes in the processes of soil water movement and nutrient cycling, and on the profile patterns of water and nutrient availability at sites of differing grazing intensities. This dual focus is vital to improving our understanding of how soil processes are affected by cattle grazing pressures. Detailed findings of process-based studies have been reported by Dougill et al. (1997, 1998a), and are simply summarized here. Specifically, studies examined the underlying assumptions of the twolayer model that nutrient mineralization into water soluble inorganic forms is increased with cattle dung additions, and that increased leaching of water and nutrients occurs due to herbaceous vegetation removal and dung additions. Studies refuted both these hypotheses for the nutrient poor, sandy soils of the Kalahari (Dougill et al. 1997, 1998a). These findings refute the assumptions of the two-layer model, alternatively suggesting that infertile sandy soils appear resilient to changes in soil water distribution and nutrient availability caused by increased cattle use. To examine whether these process-based conclusions are matched by the reality of soil hydrochemical characteristics observed in situ, field-based studies here aim to assess patterns of water and nutrient availability, and their profile partioning between topsoil and subsoil layers, at sites of different grazing intensity (and therefore ecological state). Ranch scale soil studies were carried out along a 3 km transect radiating from the main 1973 initiated borehole on the Uwe Aboo ranch. Sampling was based on a grazing gradient or piosphere approach, with study sites located 25, 100, 200, 400, 800, 1600 and 2800 m from the borehole, and in a neighboring ungrazed control site confined within the Makoba veterinary double cordon fence, which has been ungrased for over 25 years (Figure 3). At the seven sampling sites, three replicate samples were taken from the soil surface and at depths of 20 cm (topsoil) and 100 cm (subsoil) shortly after the onset of the first rains (c. 25 mm rainfall) in October 1993. More intensive sampling was conducted at the bush encroached site (400 m from the borehole) and the control site (Figure 3). This involved the collection of 23 bulked samples from each depth throughout the sampling period from September 1992 to October 1993. In addition, eight replicate samples were taken from each depth of the bush encroached and control sites following a series of intense storms during January 1994 (c.150 mm of rainfall). These samples enable us to examine differences in soil water and nutrient availability patterns between the sites for the critical ecological growth pulse following rainfall (Scholes 1990). In all cases, fresh soil samples were analyzed at the Botswana Government Soils Laboratory within 48 hours of sampling. Samples were split and a portion immediately analyzed for gravimetric water content. For soil nutrient assessments, standard extraction procedures were followed to provide surrogate measures of the inorganic nutrient concentrations available for plant uptake. For inorganic nitrogen, this means extracting with distilled water for nitrate (NO3-N) and six percent KCl solution for ammonium (NH4+-N), prior to filtration of the extractant (Allen 1989). Subsequently, NO3--N concentrations were measured using the colorimetric method of Cataldo et al. (1975, as summarized in Anderson and Ingram [1993]). NH4+-N concentrations were also assessed colorimetrically, using the modified Bertholet Reaction method (Anderson and Ingram 1993). Extractable concentrations of inorganic phosphate (PO43-P) were measured following a standard Olsen’s extraction procedure (Allen 1989) and colorimetric analysis using the molybdenum blue technique (Murphy and Riley 1962). Soil moisture characteristics Measurements of the gravimetric moisture distributions in the soil profile were taken to assess whether changes in vegetation cover, particularly reductions in the herbaceous grass cover, had increased the amount of water leached into the subsoil, as proposed by the two layer model. Figure 4 summarizes the soil moisture characteristics on the study transect during the 1993 dry season. The data suggest that there was no significant difference between samples taken from the control site and bush encroached site (confirmed by t-test comparison; p = 0.267 and 0.066 for 20 cm and 100 cm depths, respectively). The statistical similarity in subsoil moisture availability at bush encroached and control sites also characterized the pulses of high moisture availability that followed intense rainfall. After a series of intense storms during January 1994, the gravimetric moisture content at 100 cm depth remained similar at both intensively sampled sites (bush encroached = 4.10 percent ± 0.79 [n = 8]; control = 3.99 percent ± 0.92 [n = 8]). Hence, no significant increase in subsoil moisture availability was found for the bush encroached sites with its significantly reduced grass cover. These results combine with similarities between sites in other measured soil characteristics, including bulk density, field capacity and hydraulic conductivity (Dougill et al. 1997) thereby suggesting that intensive grazing, and associated vegetation changes are not linked to changes in soil water availability in Kalahari soils. Indeed, hydrological studies that trace water redistribution following storms and controlled water additions (Dougill et al. 1997) found that despite the high soil hydraulic conductivity (over 12 cm h-1), the relatively high gravimetric subsoil moisture contents (of approximately 1.0 percent at 1 m depth) and the restriction of soil water movement to matrix flow pathways, prevents any significant leaching losses beyond the rooting zone of savanna grass species. These hydrological findings directly oppose the hypotheses proposed in the two layer model. Soil inorganic nitrogen The availability of inorganic nitrogen in the soil is a principal cause of ecological productivity, and is therefore an important part of soil degradation assessments. In addition, changes to profile patterns of inorganic nitrogen availability have been proposed as vital in affecting vegetation community structure (Figure 1), due to the water soluble nature of NO3--N that leaves it prone to leaching into the subsoil, where bush roots are competitively dominant. Soil inorganic-N (NO3-N and NH4+-N) concentrations in topsoil and subsoil samples were assessed at all study sites to establish whether the decreased grass cover and increased inputs of cattle dung and urine at intensively grazed areas combine to increase the subsoil concentrations of inorganic-N, as proposed by the two layer model (Figure 1). Figure 5 displays the variations in subsoil extractable inorganic-N concentrations along the study transect. Results indicate that the inherent variability of soil inorganic-N concentrations overrides the effects of intensive grazing and/or different vegetation communities. No significant differences were found between soil inorganic-N concentrations at bush encroached and control sites (t-test comparison; p = 0.502 and 0.956 for 20 cm and 100 cm depths, respectively). Again, these findings oppose the hypothesis proposed in the two layer model. To explain the resilience to change in soil inorganic-N availability, it is necessary to consider the processes affecting the transport and transformations of nitrogen within soils. Process-based studies (Dougill et al. 1998a) have shown that these infertile sandy soils experience very low rates of net nitrogen mineralization (less than 0.1 mg N 100g-1 day-1) even with cattle dung additions to field-incubated mineralization columns (method of Raison et al. 1987). The low net mineralization rate combines with the rapid plant uptake of nutrients that is typical of semiarid vegetation (McNaughton and Chapin 1985, Gross et al. 1993), preventing the build up of elevated water-soluble nitrate concentrations in the topsoil. In addition, controlled leaching column experiments showed that even following grass removal, and therefore negligible plant nutrient uptake, the low nutrient status of Kalahari soils and the predominant water movement as matrix flow combine to allow nutrient adsorption onto soil particles (Dougill et al. 1998a). This adsorption onto topsoil particles and the low net mineralization rates combine to suggest that nitrogen cycling will remain topsoil-dominated even following the removal of grass cover and direct nutrient inputs in cattle dung and urine. These process-based findings can be used to explain the similarities recorded here in profiles of inorganic-N availability between bush-encroached and control sites (Figure 5). The full range of soil studies conducted at Uwe Aboo refute the claim that vegetation changes can be linked to increased subsoil inorganic-N concentrations (Walker and Noy-Meir 1982). Importantly, in terms of extending these studies to assessments of soil degradation it should also be noted that no significant differences (p > 0.05) in nutrient availability were found beyond the sparsely vegetated "sacrifice zone" (within 100 m of the water point), where increased nutrient concentrations are found. This increase was never matched by significant reductions in nutrient availability elsewhere. Total nitrogen levels (both organically and inorganically bound) remain high (over 500 mgN 100g-1) in both bush encroached and control settings (Dougill and Cox 1995), implying that plant availability of inorganic-N is dependent on the processes of mineralization and plant uptake, rather than on changes in the surface organic input loadings. Therefore, no evidence was found to link grazing intensity to reduced inorganic-N availability or changes in its profile availability, and no evidence of chemical soil degradation was recorded. Soil inorganic phosphorus It is widely recognized that there is an association between semiarid vegetation growth and inorganic-P availability (Penning de Vries et al. 1980, Medina, 1987, Scholes and Walker, 1993). It has also been suggested that grazing patterns in borehole-centered systems can lead to reduced inorganic-P concentrations in intensively grazed areas (Botkin et al. 1981). This could be caused by livestock reducing the organic phosphorus inputs by the consumption of grass, and subsequently depositing it, as dung, in areas of intensive use close to the borehole, and in kraals (fenced enclosures used to hold livestock at night) where they are constrained for up to eight hours a day (Tolsma et al. 1987, Perkins and Thomas 1993a). Therefore any study attempting to link vegetation changes with soil characteristics needs to quantify the influence of changes in the availability of phosphorus, chiefly in the form of extractable phosphate (PO43--P) concentrations, so as to assess whether chemical soil degradation has occurred due to reduced P availability. Figure 6 displays the change in topsoil PO43--P concentrations along the Uwe Aboo study transect. Results show that although there is a marked increase in PO43--P availability in the immediate vicinity of the water point, due to the concentration of dung inputs, there is no evidence of associated declines in PO43--P concentration in intensively grazed sites. No significant differences were noted between soil inorganic-P concentrations at the bushencroached and unencroached control sites (t-test comparison; p = 0.450 for topsoil samples where PO43--P is concentrated). Studies of phosphorus cycling in these soils (Dougill et al. 1998a) showed that net mineralization into inorganic-P was extremely slow (under 0.01 mgP 100g-1 day-1), such that synchrony between nutrient production and plant uptake prevents elevated PO43--P concentrations in any area with even limited plant growth. Furthermore, the maintenance of total-P concentrations an order of magnitude greater than inorganic-P concentrations in soils at both bush encroached and control sites (Dougill and Cox, 1995) implies that reductions in inorganic-P availability will not occur as the "mineralizable" pool of P remains high. Consequently, no evidence exists to link patterns of vegetation change with soil inorganic-P availability. Again microbial mineralization processes appear to control soil fertility, rather than subtle changes in the nature and extent of organic nutrient inputs to the soil surface. Ecological change in the Kalahari: Alternative models and future research implications The soil water and soil nutrient research reported here does not provide any evidence to link vegetation changes to key soil hydrochemical characteristics on Kalahari TGLP ranches, as previously proposed (Skarpe 1990, 1991, Perkins and Thomas 1993a). This study suggests that the two-layer model appears unlikely to be applicable to semiarid ecosystems based on sandy infertile soils. We suggest that changes in soil characteristics are not required to explain the changes in vegetation community structure. Consequently, alternative models of environmental change are required, focusing on the interactions of the other key ecological determinants, notably grazing levels, rainfall variability, and fire regimes. Whilst the link between bush encroachment and grazing intensity appears well proven (Perkins and Thomas 1993a, 1993b), uncertainties remain in our understanding of the causes and timing of this ecological transition. It is these uncertainties that presently hinder the incorporation of clear environmental conclusions into the development of agricultural management strategies aimed at reducing the extent of bush encroachment. Thus far, environmental studies have never clearly been used to suggest alternatives to the continued reliance on the land use policies of grazing levels set at fixed carrying capacities for ranches using a rotational grazing system, ideas that are based on conventional climax and equilibrium ecological theories. Only with the formulation of more applicable state and transition models and continued integrated environmental monitoring, can environmental studies be used, in association with social evidence, to produce more flexible agricultural development policies. In Figure 7 we provide a conceptual "state and transition" model that summarizes the present theoretical understanding of ecosystem changes in Kalahari pastoral systems. The variability in natural environmental conditions (rainfall and fire regimes) is incorporated as vitally important in both influencing ecosystem processes favoring bush establishment and encroachment (factors on the bottom half of Figure 7), but also in providing system resilience attributes that mitigate against any ecological changes, whether natural or grazing induced, becoming permanent states (factors noted on the top half of Figure 7). It is also clear, however, that ecosystem changes caused by intensified livestock grazing activities do favor the onset of bush encroachment. Ecological studies, thus far, have shown that bush encroachment occurred around all boreholes investigated, even those grazed for less than 10 years (Perkins and Thomas 1993a, 1993b). The fact that ecological studies show that the extent and composition of the bush canopy cover is not directly related to the age of a given borehole (Perkins and Thomas 1993a, 1993b) supports the contention, illustrated in Figure 7, that frequent disturbances from various agents including fires, drought, frost, lightning, wind, and wood harvesting, provides Kalahari rangelands with some degree of resilience to permanent ecological changes. The lack of changes in soil characteristics provide further support for this ecological resilience, a factor that is increasingly being portrayed as the key ecological attribute of dryland ecosystems (Behnke et al. 1993, Scoones 1995, Jeltsch et al. 1996). The evidence provided here is not meant to suggest a complete lack of ecological concern regarding land degradation in the Kalahari. Evidence from other parts of Botswana, such as on communal lands surrounding the village of Lethlakeng (Figure 2) that have been intensively grazed for over a century, show that shrubby bush encroachment can over time develop into stands of mature trees and bushes (a fact recognized by the final stage of the state and transition model, Figure 7). These larger bushes and trees offer less protection for underlying grasses, and can result in the demise of the niches protected against grazing through the ecological growth characteristics of certain invading species, especially Dichrostachys cinerea and certain Acacia species (including A. eriloba and A. mellifera) which can grow to over 3 m height. Therefore, Figure 7 portrays the bush encroachment presently seen in the eastern Kalahari, not as an "effectively permanent" change, but as a stage in a change of causation that may either lead to quasi-permanent ecological change, or be reversed depending on the environmental conditions experienced and the management strategies followed. The model provided in Figure 7 should also not be viewed as a definitive guide to the causes of ecological change for the Kalahari, due to the remaining uncertainties caused by the complex array of interactions between grazing, fire, and rainfall regimes, all of which are characterized by inherent spatial and temporal variability in drylands. Alternatively, the model offers a conceptual representation of the current environmental understanding regarding the processes of ecological change, and should therefore be used as a guide to the nature of integrated environmental studies needed to improve environmental understanding in the future. In particular, it highlights the need for environmental monitoring to continue assessment of the spatial and temporal extent of bush encroachment and die-back. Further information is required on the ecological impacts of different grazing levels through times of drought, and how these affect post-drought vegetation regrowth patterns. Further research must also assess the impact of fires (of differing intensities) on vegetation community structure, in association with differences in the rainfall and grazing regimes experienced before and after fire events. Such improved environmental monitoring could take the form of intensive ground-based ecological studies of selected "representative" sites, as suggested by the ongoing International Geosphere Biosphere Programme (IGBP) Kalahari Transect program; the incorporation of a greater spatial and temporal record of environmental information contained within the historical archive of satellite earth observation data; and/or greater use of the information available from fine-scale ecological studies and integrated soil and ecological studies, such as that undertaken at Uwe Aboo. All of these means of advancing environmental research methodologies face a number of potential problems, as well as opportunities, for refining state and transition models as currently formulated. Detailed monitoring of individual study sites, such as undertaken at Nylsvley in northern South Africa over the last twenty five years (Scholes and Walker 1993), requires intensive fieldwork programs and historical information on the nature of vegetation communities in the area. Unfortunately, such historical ecological information is lacking for the Kalahari, as with many other remote dryland regions, and therefore would require the initiation and continued research funding for selected sites. Other concerns exist in the selection of "representative" sites. This can be shown by the preference of previous ecological studies and the current IGBP Kalahari Transect to concentrate on nature reserve or national park sites, such as Nylsvley or Chobe National Park, due to their conservation value. Other well-studied sites include the government experimental ranches (e.g., Skarpe 1990, 1991), typified by lower grazing intensities; and on sites experiencing very visible soil erosion problems, such as the wind erosion experienced on saline soils surrounding the village of Rakops, Central Botswana (Ringrose et al. 1996). None of these sites, however, typify the land use intensification pressures imposed by agricultural policy (Republic of Botswana 1991) or the environmental situation of the majority of the Kalahari. In this regard, ongoing studies on the Makoba TGLP ranches, and also Ncojane TGLP ranches, as well as the in western Botswana (Smith et al. 1997) and the Tshabong TGLP ranches in the more arid southwest Botswana (White 1993), can hopefully be continued to provide the longer time frame of ground based ecological analysis required to further reduce environmental uncertainty. Satellite earth observation data offers the potential to highlight variations in vegetation communities from their electromagnetic reflectance characteristics over archived historical records extending back at a regional scale over the last three decades. Many studies have observed the ability of satellite imagery to assess vegetation abundance (e.g., Smith et al. 1990, Tucker et al. 1991). Problems exist, however, in using Earth observation data to measure properties of the ecological state and transition model, notably vegetation community structure and regional variations in rainfall and fire regimes. Spectral studies (e.g., Graetz and Gentle 1982, Franklin et al. 1993) have shown that both bush and grass cover cause shadowing and a "darkening" effect on bright soils, such as those of the Kalahari. While bushes cause greater shadowing, due to the limited dimensionality of reflectance data, it is not possible at this time for spectral Earth observation data to be used to estimate vegetation community structure (Trodd and Dougill 1998). Alternative approaches exploiting the information content available in the temporal domain, such as contrasting the different annual growth patterns of bushes and grasses, are being investigated for the Kalahari (Trodd et al. 1997), but as yet these cannot map spatial and temporal changes in patterns of vegetation community structure, as is needed to clarify state and transition models. Problems with these research methodologies imply that it is vital to examine the extensions that can be made to limited time-scale, and small spatial-scale, field-based studies. It is hoped that the integrated analysis of ecological and soil based studies on the range of scales provided here can highlight simple field methods that can be used to extend environmental understanding in other dynamic dryland environments. Table 2 summarizes the studies undertaken at Uwe Aboo and their scale of analysis. While this representation compartmentalizes a variety of dryland research efforts, it is essential to realize that all studies shared the aim of reducing uncertainties inherent in state and transition models and that the model we propose, and the implications for land degradation assessments discussed, stem from the integrated analysis of their findings. Greater use of soil-based studies, in particular their extension to incorporate preliminary investigations of the processes that regulate water and nutrient availability patterns, would improve environmental monitoring, modeling and management of savanna rangelands. Implications for the classification of bush encroachment as land degradation If land use and environmental circumstances do permit a quasi-permanent change of ecological state from mixed savanna to bush dominant savanna, there remains the question of whether such a change in the Kalahari is sufficiently extensive and economically damaging to be classified as a form of land degradation (de Queiroz 1993, White 1993, Dougill and Cox 1995). Answering this question demonstrates the difficulties faced in incorporating the natural variability element of non-equilibrium ecological theories into classifications of land degradation. It also reflects confusions with the meaning of the term land degradation (Dahlberg 1994). In this study we have utilized the definition of Abel and Blaikie (1989), which relates degradation to the goals of the management system adopted at the time of investigation. Whilst this definition has been questioned (Cox and Dougill 1996), it does match the inertia present in the management policies adopted in the Kalahari, and therefore appears justified on a practical basis. Importantly, it also links degradation to human use of the environment rather than to any notion of pristine "natural" conditions. Consequently, we restrict consideration to the possible impact on livestock outputs on the fenced ranches that are the core of present agricultural management policies. This ignores notions of alternative land uses and issues of equity of access to natural resources, but does relate to the current reality of Kalahari land-use policies. Previous economic studies of pastoral production in the Kalahari have shown that at a regional scale the majority of variability in livestock production can be explained by rainfall fluctuations during the previous two wet seasons (Vossen 1990, White 1993), implying that there is no economic evidence of land degradation. Soil studies reported here show additionally that no significant changes in soil hydrochemical characteristics have occurred in intensively grazed bush-encroached sites, suggesting that ecological changes may be spatially confined as well as reversible. In the low fertility Kalahari sandy soils, soil changes neither result from, nor lead to, bush encroachment. It therefore appears that ecological changes are not “effectively permanent,” since changes in grazing levels and/or environmental conditions could cause a return to grass dominance (Figure 7). Consequently, economic, ecological, and soil studies all combine to suggest that the ecological changes seen on Kalahari rangelands cannot, as yet, be equated to land degradation. Conversely, as all livestock both graze and browse in times of diminished availability of the preferred grass food source, the mixed bush and grass dominant landscapes produced on TGLP ranches with a single borehole provide for a variability in food resources that is valuable in times of drought (Scoones 1995). These arguments are not to suggest that we should be unconcerned about either livestock production or bush encroachment in the Kalahari. These concerns must be viewed as real, given the assigned link between bush encroachment and reduced pastoral productivity observed in other areas, including neighboring regions in Namibia (Quan et al. 1994) and South Africa (Dean and McDonald 1994). Future concerns result from the pervasive and expansive nature of bush encroachment (Perkins and Thomas 1993a, 1993b) and management decisions encouraged by the National Policy on Agricultural Development. That is, the increasing density of boreholes, which relaxes the informal “8 kilometer rule” on borehole spacing adopted in the TGLP, and the fencing of management units, pose definite economic threats even without the assertion that ecological changes are permanent. As management units decrease in size, the potential exists for them to become almost completely bush dominated, with a reduction in resilience mechanisms. A consequence might then be for disturbance mechanisms not to return the ecosystem to a position of grass dominance. Implications for management strategies in the Kalahari Environmental studies describing bush encroachment as an inevitable consequence of formalized cattle ranching (Figure 7) have a number of important implications for the agricultural management of Kalahari rangelands. If the prevention of more extensive, merged, areas of bush encroachment is to occur, three main factors require consideration and action. First, there is a need to consider reducing the physical defoliating effects of cattle grazing during drought events and the vital initial regrowth stage, when grasses are most susceptible to grazing-induced mortality (Coughenour et al. 1985, Mott et al. 1992). This could be achieved through the provision of rapid reaction destocking programs, as adopted in many other dryland African settings (Scoones 1995, Lane 1998), that ensure livestock owners gain an incentive to offtake animals at drought onset, and which provide the capital to enable rapid restocking in the post drought period (Toulmin 1995). Alternatively, regional programs could be initiated to allow movement of livestock to areas of adequate grass fodder supply, maximizing the regional herd size in relation to patterns of spatial forage availability caused by regional rainfall variations. Such cattle movements do occur on cattle trucks between ranches owned by the same person, but they have not reached the sophisticated level seen in some dryland regions. For instance, in Jordan modern communication systems are being used to allow trucked migratory movements over hundreds of miles to areas experiencing above average rainfall (Pearce 1995). This increased mobility would enable larger regional herds allowing the rapid restocking required in subsequent wet years, but it would require cooperation between livestock owners on a scale not presently seen in Botswana. Second, controls need to be imposed on the density of boreholes and the continued fragmentation of ranch blocks into smaller paddocks that will disturb the present balance of both bush dominant and grass dominant areas found on ranch blocks. At the very least, the financial costs of borehole drilling (which is often unsuccessful due to groundwater salinity problems) and fencing operations, the potential yield losses that could result from decreased heterogeneity of fodder resources, and the capital costs to implement bush clearance measures, need to be incorporated into clear long term cost-benefit analyses conducted before further structural reforms of ranches are initiated. In this regard, it is essential to use the increasing evidence from neighboring areas to demonstrate to land managers that increasing borehole densities does not translate directly to increased profitability. Finally, the ecological state and transition model proposed (Figure 7) implies that the role of fire can remain vital in preventing the continued expansion of bush encroachment. Further studies are required to clarify the applicability of this link and the nature of fires required to return ecosystems to a state of grass dominance for a number of growth seasons. Again, further investigation of this link requires improvements of environmental monitoring methods as suggested earlier, using long-term ecological studies, greater use of earth observation data, and/or fine scale ecological studies to examine more clearly interactions between environmental variability (rainfall and fires) and grazing levels. Conclusions Ecological studies in the Eastern Kalahari have characterized bush encroachment as the major change in vegetation communities associated with the intensification of cattle grazing over the last thirty years. However, the integrated studies reported here contest the immediate attribution of ecological change with land degradation. Degradation assessments involving economic analysis show no discernible impacts of ecological changes on livestock outputs (Vossen 1990, White 1993), and environmental studies suggest that vegetation community changes are not necessarily "effectively permanent." Studies reported here show that ecological changes have occurred independent of soil hydrochemical characteristics, and that significant grass cover remains even at intensively grazed sites. This implies therefore that bush encroachment can be reversed by pastoral management strategies and natural environmental variations of drought and fire (Figure 7). These conclusions, based on an array of ecological and soil-based studies on both ranch-level and finer scales, demonstrate the extra environmental certainty that can be attached to integrated studies, compared to solely ecological analysis. In this regard, the research presented here is a possible framework to be used to improve understanding of the causes and permanence of ecological changes for other dynamic dryland regions. Specific findings for Kalahari TGLP ranches suggest that, at present, Kalahari rangelands have not suffered land degradation per se, but are experiencing significant spatial changes in their ecological characteristics. With the continued push for further grazing intensification and land privatization, there remains a threat that livestock production will experience a marked decline in productivity in the years ahead as a direct result of the expansive process of bush encroachment. Indeed, ecological change without associated soil hydrochemical changes, and in turn declining livestock productivity, may turn out to be dual outcomes of management changes imposed on ranches by government agricultural policies. Acknowledgments The research reported here was conducted under the Botswanan Government Research permit Ref No. OP 46/1 XXXII (97). Andrew Dougill was in receipt of a University of Sheffield Postgraduate Scholarship, with additional fieldwork support from the Royal Geographical Society, Manchester Geographical Society, Duke of Edinburgh Trust, Gilchrist Educational Trust, and the Royal Society (Dudley Stamp Memorial Fund). Visits by David Thomas and Louise Heathwaite were funded by the British Council. In Botswana, the support provided by the University of Botswana is greatly appreciated. We would also like to thank the three anonymous reviewers for their comments and thoughtful reading of the manuscript. References Abel, N.O.J. and Blaikie, P.M. 1989. Land degradation, stocking rates and conservation policies in the communal rangelands of Botswana and Zimbabwe. Land Degradation and Rehabilitation 1:101-123. Allen, S.E. 1989. Chemical Analysis of Ecological Materials. London: Blackwell Scientific. Amundson, R.G., Ali, A.R. and Belsky, A.J. 1995. Micro-site effects of trees and shrubs in dry savannas. J. of Arid Environments 29:139-153. Anderson, J.M. and Ingram, J.S.I. 1993. Tropical Soil Biology and Fertility: A Handbook of Methods (2nd ed.). Wallingford: CAB International. Andrew, M.H. 1988. Grazing impact in relation to livestock watering points. Trends in Ecology and Evolution 3:336-339. Archer, S. 1990. Development and stability of grass/woody mosaics in a subtropical savanna parkalnd, Texas, U.S.A. J. of Biogeography 17:453-462. Behnke, R.H., Scoones, I. and Kerven, C., eds. 1993. Range Ecology at Disequilibrium: New Models of Natural Variability and Pastoral Adaptation in African Savannas. London: Overseas Development Institute. Belsky, A.J. 1990. Tree/grass ratios in East African savannas: a comparison of existing models. J. of Biogeography 17: 483-489. Belsky, A.J. 1994. Influences of trees on savanna productivity: tests of shade, nutrients and treegrass competition. Ecology 75:922-932. Belsky, A.J., Amundson, R.G., Duxbury, J.M., Riha, S.J., Ali, A.R. and Mwonga, S.M. 1989. The effects of trees on their physical, chemical and biological environments in a semiarid savanna in Kenya. J. of Applied Ecology 26:1005-1024. Belsky, A.J., Mwonga, S.M., Amundson, R.G., Duxbury, J.M. and Ali, A.R. 1993a. Comparitive effects of isolated trees on their undercanopy environments in high-rainfall and low-rainfall savannas. J. of Applied Ecology 30:143-155. Belsky, A.J., Mwonga, S.M. and Duxbury, J.M. 1993b. Effects of widely spaced trees and livestock grazing on understory environments in tropical savannas. Agroforestry Systems 24:120. Bhalotra, Y.P.R. 1987. Climate of Botswana. Part II. Elements of Climate. 1. Rainfall. Gaborone: Department of Meteorological Services. Biot, Y. 1993. How long can high stocking densities be sustained? In: Range Ecology at Disequilibrium, eds. Behnke, R.H., Scoones, I. and Kerven, C., pp. 153-172. London: Overseas Development Institute. Botkin, D.B., Mellilo, J.M. and Wu, L.S.Y. 1981. How ecosystem processes are linked to large mammal population dynamics. In: Dynamics of Large Populations, eds. C.W. Fowler and T.D. Smith, pp.373-387. New York: John Wiley and Sons. Buckley, R.C., Gubb, A. and Wasson, R.J. 1987a. Parallel dunefield ecosystems: predicted soil nitrogen gradient tested. J. of Arid Environments 12:105-110. Buckley, R.C., Wasson, R.J. and Gubb, A. 1987b. Phosphorus and potassium status of arid dunefield soils in central Australia and southern Africa, and biogeographic implications. J. of Arid Environments 13:211-216. Buffington, L.C. and Herbel, C.H. 1965. Vegetational changes on a semidesert grassland range from 1858 to 1963. Ecological Monographs 35:139-164. Campbell, A., Main, M. and Associates 1991. Western sandveld remote area dwellers. Gaborone: Noraid. Coughenour, M.B., McNaughton, S.J. and Wallace, L.L. 1985. Responses of an African graminoid (Themeda triandra Forsk.) to frequent defoliation, nitrogen and water: a limit of adaption to herbivory. Oecologia 68:105-110. Cox, J. and Dougill, A.J. 1996. Ecosystem dynamics and degradation in the Kalahari - A reply. Pastoral Development Network Paper 39e. London: Overseas Development Institute. Dahlberg, A. 1994. Contesting Views and Changing Paradigms: The Land Degradation Debate in Southern Africa. Oslo: Nordiska Afrikainstitutet Discussion Paper 6. Dean, W.R.J. and McDonald, I.A.W. 1994. Historical changes in stocking rates of domestic livestock as a measure of semiarid and arid rangeland degradation in the Cape Province, South Africa. J. of Arid Environments 26:281-296. Debenham, F. 1952. The Kalahari today. The Geographical Journal 118:12-23. de Queiroz, J.S. 1993. Range degradation in Botswana: Myth or reality? Pastoral Development Network Paper 35b. London: Overseas Development Institute. de Ridder, N. and Breman, H. 1993. A new approach to evaluating rangeland productivity in Sahelian countries. In: Range Ecology at Disequilibrium, eds. R.H. Behnke, I. Scoones and C Kerven, pp. 104-117. London: Overseas Development Institute. de Vries, J.J. and von Hoyer, M. 1988. Groundwater recharge studies in semiarid Botswana - a review. In: Estimation of Natural Groundwater Recharge, ed. Simmers, I. pp.339-347.. Dordrecht: D.Reidal. Dougill, A.J. and Cox, J. 1995. Land degradation and grazing in the Kalahari: New analysis and alternative perspectives. Pastoral Development Network Paper 38c. London: Overseas Development Institute. Dougill, A.J., Heathwaite, A.L. and Thomas, D.S.G. 1997. Cattle ranching and ecological change in the Kalahari, Botswana: a hydrological perspective. In: Sustainability of water resources under increasing uncertainty, pp. 469-477. Wallingford: International Association of Hydrological Sciences Publication no. 240. Dougill, A.J., Heathwaite, A.L. and Thomas, D.S.G. 1998a. Soil water movement and nutrient cycling in semiarid rangeland: vegetation change and system resilience. Hydrological Processes 12:443-459. Dougill, A.J., Trodd, N.M. and Shaw, M.J. 1998b. Spatial patterns of grass biomass in a grazed semiarid savanna: ecological implications. The North West Geographer 2:21-32. Ellis, J.E. and Swift, D.M. 1988. Stability of African pastoral ecosystems: Alternate paradigms and implicatiopns for development. J. of Range Management 41:450-459. Franklin J., Duncan, J. and Turner, D.L. 1993. Reflectance of vegetation and soil in Chihuahuan desert plant communities from ground radiometry using SPOT wavebands. Remote Sensing of Environment 46:291-304. Friedel, M.H., Pickup, G. and Nelson, D.J. 1993. The interpretation of vegetation change in a spatially and temporally diverse arid Australian landscape. J. of Arid Environments 24:241-260. Frost, W.E. and McDougald, N.K. 1989. Tree canopy effects on herbaceous production of annual rangeland during drought. J. of Range Management 42:281-283. Georgiadis, N.J. 1987. Responses of savanna grasslands to extreme use by pastoralist livestock. Ph.D. Thesis, Syracuse University, New York. Good, K. 1992. Interpreting the exceptionality of Botswana. J. of Modern African Studies 30:6995. Graetz, R.D. and Gentle, M.R. 1982. The relationships between reflectance in the Landsat wavebands and the composition of an Australian semiarid shrub rangeland. Photogrammetric Engineering and Remote Sensing 48:1721-1730. Gross, K.L., Peters, A. and Pregitzer, K.S. 1993. Fine root growth and demographic responses to nutrient patches in four old-field plant species. Oecologia 95:61-64. Hardin, G. 1968. The tragedy of the commons. Science 162:1243-1248. Hellden, U. 1991. Desertification: time for an assessment? Ambio 20:371-383. Hodgkinson, K.C., Harrington, G.N., Griffin, G.F., Noble, J.C. and Young, M.D. 1984. Management of vegetation with fire. In: Management of Australia's Rangelands, eds. G.N. Harrington, A.D Wilson and M.D Young, pp. 141-156. Melbourne: CSIRO. Holling, C.S. 1973. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics 4:1-23. Jeltsch, F., Milton, S.J., Dean, W.R.J. and van Rooyen, N. 1996. Tree spacing and coexistence in semiarid savannas. J. of Ecology 84:583-595. Jerve, A.M. 1982. Cattle and Inequality. A study in rural differentiation from southern Kgalagadi in Botswana. DERAP Publications No. 143. Bergen: Michelson Institute. Kharin, N.G. 1990. Recommendations on application in the Sahelian zone of the FAO/UNEP provisional methodology for desertification assessment and mapping. In Desertification revisited: proceedings of an ad hoc consultative meeting on the assessment of desertification. pp. 179-238. Nairobi: UNEP DC/PAC. Lane, C.R. 1998. Custodians of the Commons. Pastoral land tenure in East and West Africa. London: Earthscan. McNaughton, S.J. and Chapin, F.S. 1985. Effects of phosphorus nutrition and defoliation on C4 graminoids from the Serengeti plains. Ecology 66:1617-1629. Medina, E. 1987. Nutrients: Requirements, conservation and cycles of nutrients in the herbaceous layer. In Determinants of Tropical Savannas, ed. B.H. Walker, pp. 39-65. Oxford: IRL Press. Menaut, J.C., Gignoux, J, Prado, C. and Clobert, J. 1990. Tree community dynamics in a humid savanna of the Côte-d'Ivoire: modelling the effects of fire and competition with grass and neighbours. J. of Biogeography 17:471-481. Mott, J.J., Ludlow, M.M., Richards, J.H. and Parson, A.D. 1992. Causes of variation in seasonal response to defoliation in three tropical savanna grasses. Australian Journal of Agricultural Research 43:241-260. Moyo, S., O'Keefe, P. and Sill, M. 1993. The Southern African environment. London: Earthscan. Murphy, J. and Riley, J.P. 1962. A modified single-solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 14:318-319. Nellis, M.D. and Bussing, C.E. 1989. Desertification and sustainable development issues in Botswana. Papers and Proceedings of Applied Geography Conferences - State University of New York at Binghampton 12:99-106. Odell, M.L. 1980. Botswana's First Livestock Development Project: an experiment in agricultural transformation. Gaborone: Swedish International Development Authority. Oldeman, L.R., Hakkeling, R.T.A. and Sombroek, W.G. 1990. Global assessment of soil degradation. Wageninen: International Soil Reference Information Centre. Parr, J.F., Stewart, B.A., Hornick, S.B. and Singh, R.P. 1990. Improving the sustainability of dryland farming systems: a global perspective. Advances in Soil Science 13:1-8. Pearce, F. 1995. Shepherds or Wise men? New Scientist 2009:24-27. Penning de Vries, F.W.T., Krul, J.M. and van Keulen, H. 1980. Productivity of Sahelian rangelands in relation to the availability of nitrogen and phosphorus from the soil. In: Nitrogen Cycling in West African Ecosystems. Ecological Bulletin 33, ed. T. Rosswall, pp. 95-113. Stockholm: Swedish Natural Science Research Council. Perkins, J.S. and Thomas, D.S.G. 1993a. Environmental response and sensitivity to permanent cattle ranching, semiarid western central Botswana. In: Landscape Sensitivity, eds. D.S.G. Thomas and R.J. Allison, pp. 273-286. Chichester: J. Wiley and Sons. Perkins, J.S. and Thomas, D.S.G. 1993b. Spreading deserts or spatially confined environmental impacts? Land degradation and cattle ranching in the Kalahari desert of Botswana. Land Degradation and Rehabilitation 4:179-194. Peters, P.E. 1995. Dividing the commons: Politics, policy and culture in Botswana. Charlottesville: University of Virginia Press. Pickup, G. 1995. A simple model for predicting herbage production from rainfall in rangelands and its calibration using remotely sensed data. J. of Arid Environments 30:227-245. Quan, J., Barton, D. and Conroy, C. 1994. A preliminary assessment of the economic impact of desertification in Namibia. Windhoek: Directorate of Environmental Affairs, Research Discussion Paper no. 3. Raison, R.J., Connell, M. J. and Khanna, P.K. 1987. Methodology for studying fluxes of soil mineral-N in situ. Soil Biology and Biochemistry 19:521-530. Republic of Botswana 1991. National Policy on Agricultural Development. Government Paper No. 1 of 1991. Gaborone: Botswana Government Printer. Ringrose, S., Matheson, W., Tempest, F. and Boyle, T. 1990. The development and causes of range degradation features in southeast Botswana using multitemporal Landsat MSS imagery. Photogrammetric Engineering and Remote Sensing 56:1252-1262. Ringrose, S. Vanderpost, C. and Matheson, W. 1995. The use of integrated remotely sensed and GIS data to determine the causes of vegetation change in southern Botswana. Applied Geography 15:225-242. Ringrose, S., Chanda, R., Nkambwe, M. and Sefe, F. 1996. Environmental change in the midBoteti area of north-central Botswana: biophysical processes and human perceptions. Environmental Management 20:397-410. Sandford, S. 1983. Management of Pastoral Development in the Third World. Chichester: John Wiley and Sons. Schlesinger, W.H., Reynolds, J.F., Cunningham, G.L., Huenneke, L.F., Jarrell, W.M. Virginia, R.A. and Whitford, W.G. 1990. Biological feedbacks in global desertification. Science 247:1043-1048. Scholes, R.J. 1990. The influence of soil fertility on the ecology of southern African dry savannas. J. of Biogeography 17:415-419. Scholes, R.J. and Walker, B.H. 1993. An African Savanna: Synthesis of the Nylsvley study. Cambridge: Cambridge University Press. Scoones, I. 1995. New directions in pastoral development in Africa. In Living with Uncertainty: New Directions in Pastoral Development in Africa, ed. I. Scoones. pp. 1-36. London: Intermediate Technology Publications Limited. Skarpe, C. 1990. Shrub layer dynamics under different herbivore densities in an arid savanna, Botswana. J. of Applied Ecology 27:873-885. Skarpe, C. 1991. Spatial patterns and dynamics of woody vegetation in an arid savanna. J. of Vegetation Science 2:565-572. Skarpe, C. and Bergström, R. 1986. Nutrient content and digestibility of forage plants in relation to plant phenology and rainfall in the Kalahari, Botswana. J. of Arid Environments 11:147-164. Smith, M.O., Ustin, S.L., Adams, J.B. and Gilespie, A.R. 1990. Vegetation in deserts: I. A regional measure of abundance from multispectral images. Remote Sensing of Environment 31:126. Smith, R.D., Thomas, D.S.G. and Heathwaite, A.L. 1997. Cattle ranching, soil nutrients and desertification in an arid savanna. Paper presented at Open Savanna Biogeography Workshop. Leeds: University of Leeds. Stocking, M.A. 1995. Soil erosion and land degradation. In: Environmental Science for Environmental Management, ed. T. O'Riordan, pp. 223-242. London: Longman. Stoddart, L.A., Smith, A.D. and Box, T.W. 1975. Range Management. New York: McGrawHill. Thomas, D.S.G. 1997. Science and the desertification debate. J. of Arid Environments 37:599608. Thomas, D.S.G. and Middleton, N.J. 1994. Desertification: exploding the myth. Chichester: John Wiley and Sons. Thomas, D.S.G. and Shaw, P.A. 1990. The deposition and development of the Kalahari Group sediments, central southern Africa. J. of African Earth Sciences 10:187-197. Thomas, D.S.G. and Shaw, P.A. 1991. The Kalahari Environment. Cambridge: Cambrige University Press. Thomas, D.S.G. and Sporton, D. 1997. Understanding the dynamics of social and environmental variability: The impacts of structural land use change on the environment and peoples of the Kalahari, Botswana. Applied Geography 17:11-27. Tolsma, D.J., Earnst, W.H.O. and Verwey, R.A. 1987. Nutrients in soil and vegetation around 2 artificial water points in eastern Botswana. J. of Applied Ecology 24:991-1000. Toulmin, C. 1995. Tracking through drought: options for destocking and restocking. In: Living with Uncertainty: New directions in pastoral development in Africa, ed. I. Scoones. pp.95-115. London: Intermediate Publications Limited. Trodd, N.M. and Dougill, A.J. 1998. Monitoring vegetation dynamics in semiarid rangelands: use and limitations of Earth observation data to characterise vegetation structure. Applied Geography 18: Trodd, N.M., Dougill, A.J. and O'Connell, N.M. 1997. Monitoring vegetation dynamics in semiarid ecosystems: the influence of vegetation structure on the relationships between rainfall and NDVI. In: Observations and Interactions, Proceedings of the Annual Conference of the Remote Sensing Society, pp. 373-378. Nottingham: Remote Sensing Society. Tsimako, B. 1991. The Tribal Grazing Land Policy (TGLP) Ranches: Performance to Date. Gaborone, Botswana: Ministry of Agriculture. Tucker, C.J., Dregne, H.E. and Newcomb, W.W. 1991. Expansion and contraction of the Sahara desert from 1980 to 1990. Science 253:299-301. Middleton, N.J. and Thomas, D.S.G. (eds) 1997. World Atlas of Desertification. 2nd edition, London: Edward Arnold/Nairobi: UNEP). van Vegten, J.A. 1983. Thornbush invasion in a savanna ecosystem in eastern Botswana. Vegetatio 56:3-7. Vetaas, O.R. 1992. Micro-site effects of trees and shrubs in dry savannas. J. of Vegetation Science 3:337-344. Vossen, P. 1990. Algorithm for the simulation of bare sandy soil evaporation and its application for the assessment of planted areas in Botswana. Agricultural and Forest Meteorology 30:173188. Walker, B.H. and Noy-Meir, I. 1982. Aspects of the stability and resilience of savanna ecosystems. In: Ecology of Tropical Savannas, eds. B.J. Huntley and B.H. Walker, pp. 556-590. Berlin: Springer-Verlag. Walker, B.H., Ludwig, D., Holling, C.S. and Peterman, R.S. 1981. Stability of semiarid savanna grazing systems. J. of Ecology 69:473-498. Warren, A. 1995. Changing understandings of African pastoralism and the nature of environmental paradigms. Transactions, Institute of British Geographers 20:193-203. Warren, A. and Agnew, C. 1988. An Assessment of Desertification and Land Degradation in Arid and Semiarid Areas. London: International Institute for Environment and Development. Warren, A. and Khoghali, M. 1992. Assessment of Desertification and Drought in the Sudanosahelian region 1985-1991. New York: United Nations Sudano-Sahelian Office. Weare, P.R. and Yalala, A. 1971. Provisional vegetation map of Botswana. Botswana Notes and Records 3:131-152. Westoby, M., Walker, B.H. and Noy-Meir, I. 1989. Opportunistic management for rangelands not at equilibrium. J. of Range Management 42:266-274. White, R. 1993. Livestock development and pastoral production on communal rangeland in Botswana. Gaborone: Botswana Society. Whitford, W.G., Martinez-Turanzas, G. and Martinez-Mena, E. 1995. Persistence of desertified ecosystems: explanations and implications. Environmental Monitoring and Assessment 37:319322. Wiens, J.A. 1977. On competition and variable environments. American Scientist 65:590-597. Table 1. Change or Degradation? Issues for studies of dryland degradation in light of nonequilibrium ecological theory. Issue Temporal Change: Vegetation in constant flux. Implication Degradation cannot be assumed from vegetation change alone. Relevance to Kalahari studies Changes in vegetation communities used to infer degradation (e.g., Nellis and Bussing 1989, Ringrose et al. 1990, Skarpe 1990; deQueiroz 1993) should be discounted. Some consideration of ecological determining factors (soil water and nutrient availability, fire and grazing) is required. Spatial Change: Complex mosaics of vegetation may occur. Necessary to distinguish change caused by management practices from natural heterogeneity. Some changes clearly ascribable to management (e.g., sacrifice zone by boreholes; Perkins and Thomas 1993a). Other changes need longer term studies of dynamics to assign a cause. Thresholds: Changes in ecosystem state may occur rapidly due to management actions and/or natural events. Need to investigate ecological changes in the context of thresholds dependent on climatic conditions and land use practices. Need for long term ecological monitoring to assess when thresholds are crossed. Vitally important are the ecological implications of interactions between grazing levels, drought events, and fire occurrence. Table 2. Research framework followed in Makoba TGLP ranches. Mode of study Fine scale Ranch scale Soil based Process studies: water redistribution, mineralization and Leaching (Dougill et al. 1998a). Profile soil water and nutrient availability patterns in relation to grazing intensity on Uwe Aboo (reported here). Bush and grass cover patterns, and interrelations, for 10 m2 quadrats (Dougill et al. 1998b). Links between patterns of ecological community structure and grazing intensity on eight ranches (Perkins and Thomas 1993a,b). Spectral reflectance Patterns of reflectance Monitoring of spatial studies on bush and grass canopies (Trodd and Dougill 1998). change away from boreholes (Trodd and Dougill 1998). Ecological Remote sensing Regional scale -- -- patterns of change in temporal response to rainfall, as an indicator of vegetation community structure (Trodd et al. 1997). Figure 1. Links between cattle grazing, ecology and soil hydrochemical characteristics as proposed in the two-layer model (Walker and Noy-Meir 1982). Figure 2. Location of study area. Figure 3. Uwe Aboo ranch in 1993 showing study transect. Figure 4. Spatial variation in gravimetric water content with distance from borehole. Plots display mean and 95 % confidence intervals for topsoil and subsoil samples. Figure 5. Spatial variation in subsoil inorganic-N concentrations with distance from borehole. Plots display mean and 95 % confidence intervals. Figure 6. Spatial variation in topsoil inorganic-P concentration with distance from borehole. Plots display mean and 95 % confidence intervals. Figure 7. State and transition model of ecological change on Kalahari rangelands.