Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
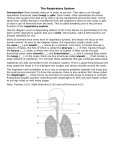
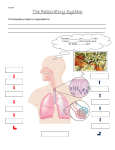
Topic 19: OXYGEN UPTAKE AND TRANSPORT (lectures 29-30) OBJECTIVES: 1. Be able to compare air vs. water as a respiratory medium with respect to oxygen content, diffusion coefficient, viscosity and water loss. 2. Be able to give some examples of organisms that use passive diffusion for gas exchange and where they might be found. 3. Have a general understanding of specialized gas exchange organs such as gills, insect tracheae and air capillary and alveolar lungs. 4. Be able to describe the concept of partial pressure. 5. Be able to describe the role of respiratory pigments like hemoglobin and how they work (i.e., reversible oxygen binding). 6. Describe the three ways in which CO2 is transported in the blood. Virtually every animal must take up oxygen from its environment and release carbon dioxide; there a few exceptions, namely intestinal parasites, worms living in anaerobic muds etc. Terrestrial organisms obtain oxygen from the air while most aquatic organisms obtain oxygen form water (the exception, of course, being such air breathers as aquatic mammals, birds, reptiles). Water: Air: O2 O2 CO2 CO2 These two respiratory media (air & water) impose rather different constraints on oxygen uptake: 1. oxygen content- oxygen content of air is at least 30 times higher than water; oxygen content of water decreases with increasing temperature and salt content 2. diffusion coefficient (D)- recall the diffusion equation (J = (D x A x C)/d); the D value for oxygen in air is 10,000 times higher than in water; oxygen does not diffuse 1 very rapidly in water; this means that most large, active aquatic organisms must move the water (ventilate= convective movement of respiratory medium) across their gas exchange surface) 3. viscosity of the medium- viscosity is a measure of the resistance to flow of a medium; water is much more viscous than air. Ventilation of water across gas exchange surface requires greater expenditure of energy 4. water content- water vapor content of air may be very low so air breathers often lose large amount of water during respiration ( respiratory water loss) Gas Exchange Mechanisms I. Passive diffusion across the body surface 1. aquatic animals- because the D value for oxygen is so low, passive diffusion works only in small animals and/or in large, thin, sluggish species like sea anemones. 2. Terrestrial animals- characteristic of earthworms, land planarians, slugs; there also is a group of lungless salamanders who rely on gas exchange across their skin (cutaneous gas exchange); these animals are restricted to moist habitats II. Specialized gas exchange organs which may or may not be ventilated- (a) gillsfleshy, blood-filled structures bathed in water, (b) tracheal system- system of tubes in insects with direct connection to the exterior and (c) lungs- internal gas exchange structures in a few fish, terrestrial amphibians, reptiles, mammals and birds. Gills: gills of more active, complex animals are internal to the extent that they are protected; by the operculum in the case of fish and the carapace in the case of crustaceans. Fig. 42.20- fish gills are complex and have a very high surface area (why?); each gill consists of a number of arches with each arch having many filaments and each filament containing many lamellae. Lamellae are very thin and contain a rich blood supply. The blood flows in the opposite direction of water passing over the gill (fig. 42.21); this creates a counter-current flow which optimizes the amount of oxygen that can be extracted from the medium. Ventilation is produced by pumping of the mouth cavity or simply by swimming with the mouth open which is called ram jet ventilation (as seen in tuna, sharks) Insect tracheal system (fig. 42.22): insects are unique in that they have a system of small tubes connecting to the surface by way of a valves called spiracles. These tubes called trachea break up into smaller tubes called tracheoles which actually travel all the way to the surface of cells ( one can find electron microscopy pictures of these tracheoles touching parts of cells where there is a cluster of mitochondria!). When spiracles are open, air flows passively into the system. Ventilation is generally not required for relatively sluggish insects due to the fact that the D value is so high in air. Flying insects do ventilate because their oxygen requirements are so much higher. 2 NOTE: The fossil record shows that flying insects attained rather large size in the past; these animals lived at a time when the oxygen content of air was ~28% as opposed to the present ~21%! Lungs: there are two types- (1) alveolar lungs (amphibians, reptiles and mammals) and (2) air capillary lungs (birds) Alveolar lungs (fig. 42.23); consist of a series of tubes which branch off into increasing smaller tubes that ultimate terminate in very tiny clusters of air sacs known as alveoli. Aveoli have the following characteristics: 1. membranes are very thin 2. they are covered by a dense mesh-work of capillaries where gas exchange takes place 3. the aveoli represent a very high surface area for gas exchange (a single human lung has 75 m2 of alveolar surface area) lungs are ventilated; that is, they inflate during inspiration and deflate during expiration. This process is accomplished by alternating contractions and relaxation of the diaphragm, a muscle located at the base of the thoracic cavityfig. 42.24 1. inspiration- diaphragm contracts; creates a negative pressure inside lungs; air is sucked in to inflate the lungs 2. expiration- diaphragm relaxes; creates a positive pressure in the lungs; pushing air out so that the lungs deflate Air capillary lungs (fig. 42.25); these lungs lack alveoli but instead have thousands of rigid tubes called air capillaries (or parabronchi) that run the length of the lung. The volume of the lungs never changes. Instead, the lungs are connected to a series of ventilatory structures known as air sacs. There are anterior and posterior sets of air sacs. These structures inflate during inspiration and deflate during expiration. What is unique about this system is that air passes through the parabronchi during both inspiration and expiration - the lung is continuously ventilated! This is a adaptation which permits high rates of gas exchange, this is essential for flight which is costly. Blood oxygen transport Gases flow from areas of high concentration to areas of low concentration; we normally refer to gas concentration by the term partial pressure. Partial pressure- the fractional pressure of a gas in a gas mixture; air is 21% oxygen and the total pressure of air at sea level is around 760 mm Hg so the partial pressure of oxygen is 0.21 x 760 = 160 mm Hg. Using partial pressure to express gas levels we call follow the changes in gas partial pressure as blood passes through the pulmonary and 3 systemic circulations (fig. 42.27). Both oxygen and carbon dioxide flow from areas of high partial pressure to areas of low partial pressure (always in the opposite direction). Oxygen flows from the air in the alveolus into the blood in the capillaries; what determines the amount of oxygen that can move into the blood? If you look at the blood leaving the lungs of a mammal, you will find that only about 5% of the oxygen the blood is carrying is dissolved in direction solution in the plasma; the rest (95%) is present in the red blood cells. In vertebrates, oxygen binds to a red pigment protein called hemoglobin (abbreviated Hb). Hemoglobin is a protein that contains an atom of iron (Fe) in its center. Hb has the capacity to bind oxygen at high partial pressures and release it at low partial pressures. This is known as reversible oxygen binding and can be shown as follows: high oxygen levels oxygen + hemoglobin oxygen-hemoglobin (deoxyHb) (oxyHb) low oxygen levels Most vertebrate hemoglobins have four subunit proteins each capable of binding one O 2 molecule so that each hemoglobin has the capacity to bind 4 O 2 molecules. Each red blood cell may contain several million molecules of Hb. The more red blood cells in a volume of blood, the greater the potential for carrying oxygen. Recall fig. 42.27, When the blood leaves the alveolus, the oxygen partial pressure is 104 mmHg; under these circumstances most of the molecules of Hb have oxygen bound to them. When the blood reaches the tissues its oxygen partial pressure falls to 40 mm Hg. The Hb can hold a much smaller amount of oxygen under conditions, much less than what was bound in the alveolus. Thus, oxygen is released by the Hb and it is taken up by the tissues. This is a manifestation of reversible oxygen binding. Hemoglobin is classified as a respiratory pigment because it has a color and it is involved in oxygen transport. There are a number of other respiratory pigments including the blue copper- containing pigment hemocyanin (molluscs, arthropods), the green pigment chlorocruorin (some marine worms) and the purple pigment hemerythrin ( some marine invertebrate groups). Carbon dioxide transport- fig. 42.29; roughly 7% carbon dioxide is transported as molecular CO2; 23% of CO2 is directly bound to Hb and the remaining 70% of CO 2 travels in the plasma as bicarbonate (HCO3-). At the site of oxygen delivery (in the cells and tissues of the systemic circulation), a large fraction of CO2 is goes through the following reaction as catalyzed by the enzyme carbonic anhydrase: CO2 + H2O H2CO3 which spontaneously dissociates to H+ + HCO34 At the site of oxygen uptake in the alveoli, the above reaction is reversed and the bicarbonate converts back to CO2. 5