Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
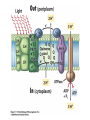
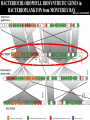
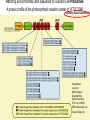
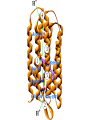
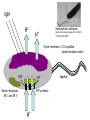
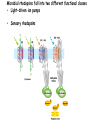
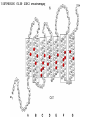
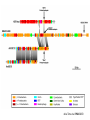
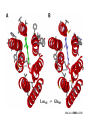
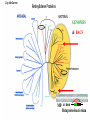
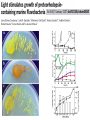
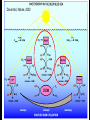
“Non-oxygenic microbial photophysiologies in the ocean: rhodopsin and bacteriochlorophyll based systems” Agouron Microbial Oceanography Summer Course 2007 Photophysiology in the sea Solar energy Photosynthesis Plants Algae, photosynthetic bacteria CO2 + carbon dioxide H2O water Chemical energy or heat N,P,S,Fe…. Respiration Animals Bacteria C6H12O6 + O2 organic carbon oxygen Dave Karl, Nature, 2002 OTHER SORTS of PHOTOTROPHY Type Electron donor C source Photolithoautotroph H2O, H2S, S0, H2 CO2 Photolithoheterotroph H2O, H2S, S0, H2 Organic substrate Photoorganoautotroph Organic substrate CO2 Photoorganoheterotroph Organic substrate Organic substrate Photomixotroph Mixed inorganic/organic Mixed inorganic/organic http://helios.bto.ed.ac.uk/bto/microbes/winograd.htm http://ecosystems.mbl.edu/SES/MicrobialMethods/Winogradsky/default.htm O2 Winogradsky column H 2S OXYGENIC PAs b a LOTS OF DIVERSITY IN BACTERIAL ANOXYGENIC PHOTOTROPHS ! General features of anaerobic photosynthetic bacteria Many grow photoorganotrophically in the absence of oxygen When growing phototrophically, derive most of their ATP from light Carbon sources used predominantly for reducing power, biosynthesis Many are capable of photoautotrophic growth 1% to 6 % of isolates from sand, seaweed, seawater, sediments Tokyo Bay Erythrobacter longus Erythrobacter sp. OCh114. (Roseobacter denitrificans) Roseobacter litoralis Shimada coined the term in 1995 : “Aerobic anoxygenic phototrophs ” Not capable of anaerobic phototrophic growth; most strict aerobes Wide variety and large amounts of carotenoids Relatively low amounts of bacteriochlorophyll a Appear not able use light as sole source of energy Light-induced oxid./reduct. of photosynthetic apparatus demonstrated Mostly organotrophic (carbon used for energy and as carbon source) Aerobic Anoxygenic Phototrophic Bacteria MICROBIOLOGY AND MOLECULAR BIOLOGY REVIEWS, Sept. 1998, p. 695ミ724. Yurkov and Beatty “Pump and probe” or Fast repetition rate fluorometry IRFRR Instrument Vent photosyn !!! Bcll-containing bacteria may contribute 2- 5 % photosynthetic electron transport in the upper ocean Kolber et al. 2000. Nature 407:178 “Photosynthetically competent anoxygenic phototrophic bacteria comprise at least 11% ofthe total microbial community” Kolber et al. Science 292:2492 P/I curves and CO2 fixation in NAP-1 isolate (a little more controversial…) Kolber et al. Science 292:2492 “Daily cellular rates of CO2 fixation about or 3% of the cellular carbon content...” Anapleurotic reactions ! • TCA cycle intermediates are used to provide carbon skeletons for other biomolecules. Cycle would halt if OAA is not replaced. • Anapleurotic reactions produce TCA cycle intermediates from pyruvate or PEP. Oceanic puf M/L phylogeny Béjà, Suzuki, et al. 2002. Nature 415:630-633 BACTERIOCHLOROPHYLL BIOSYNTHETIC GENES in BACTERIOPLANKTON from MONTEREY BAY BEJA et al, 2002 NATURE Matching Environmental DNA Sequence to Cultured Cell Proteomes: A protein profile of the photosynthetic reaction center of HTCC2080 Courtesy Steve Giovannoni Unpublished: Jang Cho Martha Degan Doug Barofsky Steve Giovannoni HTC Lab (LIONS)/ EHSC Mass Spec Lab Oregon State Univ. Cho et al. OM60 and Congregibacter littoralis HALOARCHAEA light H+ Halobacterium salinarum H+ (electron microscope image)9 0.5-1.2 um x 1.0-6.0 um in size10 Purple membrane = 2-D crystalline bacteriorhodopsin lattice ADP ATP Sensor rhodopsins SR I and SR II ATP-synthase H+ flagellae QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. The cycle can be formally described in terms of 6 steps : isomerization (I), ion transport (T), accessibility change (switch S). Retinal first photo-isomerizes from an all-trans to a 13-cis configuration followed by a proton transfer from the Schiff base to the proton acceptor Asp-85. To allow vectoriality, reprotonation of the Schiff base from Asp-85 must be excluded. Thus, its accessibility is switched from extracellular to intracellular. The Schiff base is then reprotonated from Asp-96 in the cytoplasmic channel. After reprotonation of Asp-96 from the cytoplasmic surface, retinal reisomerizes thermally and the accessibility of the Schiff base switches back to extracellular to reestablish the initial state. http://www.biochem.mpg.de/oesterhelt/photobiology/br.html Genome sequence of Halobacterium species NRC-1 Wailap Victor N et al., PNAS | October 24, 2000 | vol. 97 | no. 22 | 12176-12181 QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Microbial rhodopsins fall into two different functional classes • Light-driven ion pumps • Sensory rhodopsins LIBRARY CONSTRUCTION AND SCREENING SAR86 130 kbp BAC 1 1 “SAR86” 130kb GENOME FRAGMENT LIGHT-DRIVEN PROTON PUMPING IN E. COLI (via ТSAR86У PROTEORHODOPSIN ) Oded Beja OFF + + Expression of proteorhodopsin in E. coli ON + + 5 min pH 0.02 Proteorhodopsin Retinal ON OFF Béjà et al. Science 289: 1902-1906 (2000) Fast photcycle kinetics Phylogenetic distribution of proteorhodopsin variants SAR86 SUBGROUPS from the COASTAL and OPEN OCEANS Sabehi et al., Environ. Microbiol. 6:903(2004) Monterey Red Sea Hawaii Do different SAR86 phylotypes encode proteorhodopsins ? Phylogentic relationships of naturally occurring SAR86 ribotypes env. clone MB11B0, AY033326 env. clone MB11E0, AY033304 env. clone MB12D 0, AY033314 env. clone MB11G0, AY033311 89 BA C clo ne e BACR ed20 E0 9 BA C clo ne e BACHOT4 E0 7 env. clone OM10, U70693 env. clone OC S44, AF001650 env. clone KTc0917, AF173974 BAC clone EBAC31A08 AF279106 env. clone Z D0108, AJ400345 env. clone NAC11-19, AF245642 10 0 97 env. clone MB12G1, AY033317 env. clone CH AB-III -1,AJ240912 env. clone MB12G0, AY033328 env. cloneKTc1112, AF241654 82 env. cloneKTc1121, AF241653 env. cloneKTc1107, AF173975 10 0 env. clone ARCTIC97A-18, AF354613 env. cloneOC S5, AF001651 10 0 marine bacterium Z D0107, AJ400344 BAC clone EBAC27G05, AF268217 10 0 EB750-02H09, AY458632 marine bacterium Z D0433, AJ400356 BAC C lone EB000-65A11 ** ** *** 10 0 0.10 Sar86 - I SSU rRNA Sar86 - II Sar86 - IIIa Sar86 - IIIb de la Torre et al. PNAS 2003 (Pelagibacter) Venter et al., Environmental Genome Shotgun Sequencing of the Sargasso Sea, Science 394:66-74 (2004) ARCHAEA CRENARCHAEOTA EURYARCHAEOTA Methanobacteriales Thermoplasmatales Methanococcales ТMarine Group IУ Thermococcales Archaeoglobales ТMarine Group IIIУ ТMarine Group IIУ Methanopyrales ТMarine Group IVУ pSL12 Haloarchaea Thermoproteales Sulfolobales pJP33 Methanomicrobiales To Eucarya, Bacteria Depth-specific differences in proteorhodopsin variants Béjà et al. Nature 411:786-789 (2001) Leu105 -> Gln105 Man et al. EMBO J. 2003 Man et al. EMBO J. 2003 Sabehi et al. ISME J. 2007 Glu96 (cytoplasmic H+ donor) Asp85 (periplasmic H+ acceptor) Leu105 -> Gln105 Sensory rhodopsins lack the cytoplasmic proton donor - 22 of Sargasso Sea PR variants have either Thr (18), Ile (3), or Lys (1). Each appears linked in an operon to a putative sensory rhodopsin. Sensory rhodopsins in bacteria R ST R ST R ST R ST R, rhodopsin ST, Signal transducer (histidine kinase domain) Sharma,et al. TRENDS in Microbiology Vol.14 p. 463, 2006 Jay McCarren GENOMES & BACS PR-1 PR-2 MB_41B09 Betaproteobacterium “Typical” carotenoid (retinal) biosynthesis genes co-associated with PRs blh Exceptions: GII Archaea, Pelagibacter, a few others, PR unlinked to retinal biosynthetic operon CFB = PR-blh linkage (Pinhassi and colleagues) Proteorhodopsin photosystem gene organization PR crtE crtI crtB crtY blh Ipp moaE Jay McCarren/Chon Martinez PR,carotenoid and retinal biosynthetic gene co-evolution ? Jay McCarren crtB crtI crtB * * * * Gram - crtE * crtY * CFB Gram - Chon Martinez X X X X A single genetic event can confer phototrophy ~ 1e5 ATP/cell/min Distribution of PR photosystems among marine bacteria Life on Earth Today: The Foundation Solar energy Photosynthesis Plants Algae, photosynthetic bacteria CO2 + carbon dioxide H2O water Chemical energy or heat N,P,S,Fe…. Respiration Animals Bacteria C6H12O6 + O2 organic carbon oxygen Dave Karl, Nature, 2002