Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
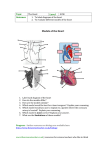
Chapter 11 Multi-Step Tumorigenesis -11.1 ~ 11.5 -11.9 ~ 11.17 - May 22 & 24, 2007 In the survival of favoured individuals and races, during the constantly-recurring struggle for existence, we see a powerful and ever-acting form of selection. ~ Charles Darwin, 1859 ~ Tumor progression Normal cells evolve into cells with increasingly neoplastic phenotypes through a process termed tumor progression. Tumor progression is driven by a sequence of randomly occurring mutations and epigenetic alterations of DNA that affect the genes controlling cell proliferation, survival, and other traits associated with the malignant cell phenotype. How many different sequential changes are actually required in cells and tissues in order to create a human cancer? 11.1 most human cancers develop over many decades of time Figure 11.1 The Biology of Cancer (© Garland Science 2007) 11.2 Histopathology provides evidence of multi-step tumor formation The notion of human tumor development as a multi-step process has been documented most clearly in the epithelia of the intestine. Microanatomy of the normal intestinal wall villi columnar epithelium basement membrane mesenchymal core (fibroblasts, endothelial cells, macrophages, etc.) The epithelial layer is the site of most of the pathological (disease-associated) changes associated with the development of colon carcinomas. Figure 11.5 The Biology of Cancer (© Garland Science 2007) Four major types of new tissue growth 肥大 增生 grow faster thicker epithelium 不正常發育 惡性增生 (腫瘤形成) Kleinsmith, L. J. Principles of Cancer Biology. Pearson Education, Inc. 2006 Fig. 1-3 Normal colonic epithelium → adenomas crypts (腺窩) adenomatous crypt normal crypts Figure 11.6 part 1 of 2 The Biology of Cancer (© Garland Science 2007) Pre-invasive adenomas (pedunculated polyps) 含肉莖的息肉 Adenomatous growths are considered benign, in that none has broken through the basement membrane and invaded underlying stromal tissues. Figure 2.15a The Biology of Cancer (© Garland Science 2007) adenoma → invasive carcinoma → metastatic carcinoma normal liver tissue evidence for adenoma-to-carcinoma progression Figure 11.6 & 8 The Biology of Cancer (© Garland Science 2007) Two more evidences for adenoma-to-carcinoma progression hundreds of polys on the wall of the colon from an individual with familial adenomatous polyposis (FAP) → high risk of colon cancer FAP individuals contain a hereditary mutation of adenomatous polyposis coli (APC) tumor suppressor gene Figure 11.8b & 7.22 The Biology of Cancer (© Garland Science 2007) Multi-step tumorigenesis in a variety of organs (carcinoma in situ) prostatic intraepithelial neoplasia cervical intraepithelial neoplasia Figure 11.7 The Biology of Cancer (© Garland Science 2007) ductal carcinoma in situ 11.3 Colonic growths accumulate genetic alterations as tumor progression proceeds LOH in colon cancers 17p: p53 5q: APC Figure 7.14 The Biology of Cancer (© Garland Science 2007) Molecular alterations during human colon tumor progression (~ 40-50 %) (~ 90 %) * (~ 60 %) *(~ 50-70 %) * tumors bearing K-ras oncogenes rarely have mutant p53 alleles, and vice versa. The precise contribution of hypomethylation to tumor progression remains unclear; some evidence suggests that it creates chromosomal instability. Figure 11.10 The Biology of Cancer (© Garland Science 2007) Alternative genetic paths during colon cancer progression almost always the 1st inactivation in the progression Individuals with familial adenomatous polyposis (FAP) start here ! Figure 11.11a The Biology of Cancer (© Garland Science 2007) Darwinian evolution and clonal successions Figure 11.12 The Biology of Cancer (© Garland Science 2007) Paradox - A mutated, activated H-ras oncogene from a human bladder carcinoma could fully transform NIH3T3 mouse fibroblasts. → the acquisition of a single ras oncogene (a point mutation on ras) sufficed to convert NIH3T3 cells to a transformed, tumorigenic state. - Why is cancer formation in humans a multi-step process ? 11.9 Multiple lines of evidence reveal that normal cells are resistant to transformation by a single mutated gene - The NIH3T3 cells are not truly normal. They constitute a cell line, a population of cells that have been adapted to grow in culture and to proliferate in an immortalized fashion. - The results obtained from primary rat and hamster primary cells (transfection of ras oncogene) were very different from those observed previously with NIH3T3 cells. These primary cells were not susceptible to ras-induced transformation. - Single mutations are not sufficient for the development of cancers. 11.10 Transformation usually requires collaborration between two or more mutant genes - A line of human promyelocytic leukemia cells carry both an activated N-ras and an activated myc oncogene. - When a myc oncogene was introduced together with an H-ras oncogene into rat embryo fibroblasts, the cells become transformed and tumorigenic. Figure 11.23 The Biology of Cancer (© Garland Science 2007) The ras and myc oncogenes have complementary effects on cell phenotype ras: anchorage independence round and refractile appearance loss of contact inhibition (cytoplasmic mitogenic signaling cascade) myc: immortalization reduction of dependence on growth factors (perturb cell cycle control machinery) or a mutant p53) Not all “ras-like” or “myc-like” members elicit identical effects in cells. Table 11.1 The Biology of Cancer (© Garland Science 2007) Table 11.2 The Biology of Cancer (© Garland Science 2007) 11.11 Transgenic mice provide models of oncogene collaboration and multi-step cell transformation mouse mammary tumor virus (MMTV): a retrovirus that specifically targets mammary tissues. The viral promoter is expressed at significant levels only in mammary glands, to a lesser extent , in salivary glands. Figure 11.24a part 1 of 2 The Biology of Cancer (© Garland Science 2007) Figure 11.24a part 2 of 2 The Biology of Cancer (© Garland Science 2007) Oncogene collaboration in transgenic, cancer-prone mice Even with mutant ras + myc expressed in the great majority of mammary cells from early in development, tumors did not appear in these mice until 4 weeks after birth. Careful analysis of rat cells transformed by ras + myc, sooner or later, acquire a mutation or methylation event that leads to inactivation of the p53 tumor suppressor pathway. Figure 11.24b The Biology of Cancer (© Garland Science 2007) Collaboration of myc and bcl-2 family genes (anti-apoptotic) in cancer development myc and bcl-2 (bcl-XL) genes are driven by immunoglobulin G (IgG) promoter and lymphomas/plasma cell tumors are scored. Figure 9.22b, c The Biology of Cancer (© Garland Science 2007) 11.12 Human cells are constructed to be highly resistant to immortalization and transformation - Human cells rarely, if ever, become immortalized following extended serial passaging in culture. - The introduction of paired oncogenes (such as ras + myc) could transform primary rodent cells, however, such pairs consistently fail to yield tumorigenic human cells. a possible explanation: - The cells of laboratory mice carry extremely long telomeric DNA (up to ~ 40 kb) and express telomerase activity. - In contrast, normal human cells have far shorter telomeres (5~10 kb), and most human cell types lack telomerase activity. Figure 11.25 The Biology of Cancer (© Garland Science 2007) Human cells + hTERT gene (prevent telomere erosion) + SV40 large T gene (inactivate both pRb and p53) immortalization + ras morphological transformation in culture (still unable to form tumors in nude mice) + SV40 small T gene (perturb the function of protein phosphatase 2A, or PP2A) tumorigenic Five distinct intracellular pathways involved in human cell transformation 1. Mitogenic signaling pathway controlled by Ras (Ch 6) 2. Cell cycle checkpoint controlled by pRB (Ch 8) 3. Alarm pathway controlled by p53 (Ch 9) 4. Telomere maintenance pathway controlled by hTERT (Ch 10) 5. Signaling pathway controlled by protein phosphatase 2A Figure 11.25 The Biology of Cancer (© Garland Science 2007) alterations are commonly seen in human cancers Figure 11.43 The Biology of Cancer (© Garland Science 2007) Why are mouse and human different? a speculation: - The cells in a mouse pass through about 1011 mitosis in a mouse lifetime, while those in a human body pass through about 1016 cell cycles in a human lifetime. (Mice have only about 0.1% as many cells as humans and live on average only about 1% of a human lifespan.) - Would the 105-fold more cell divisions in humans create a greater lifetime cancer risk ? - Have human cells and tissues evolved to have additional anti-tumor mechanisms than those of small, short-lived mammals, such as mice ? 10.9 Telomeres play different roles in the cells of laboratory mice and in human cells - Mouse telomeres are so long that they are never in danger of eroding down to critically short lengths during the lifetime of a mouse. - Therefore, laboratory mice do not relay on telomere length to limit the replicative capacity of normal cells. - Telomere erosion is also not a mechanism for constraining tumor development in laboratory mice. 11.13 Non-mutagenic agents, including those favoring cell proliferation, make important contributions to tumorigenesis Are all mutagens carcinogens? Are all carcinogens mutagens? Indication of the importance of nonmutagenic (nongenotoxic) carcinogens came from the study of “tumor promotion” in 1940s. Figure 11.27 The Biology of Cancer (© Garland Science 2007) Induction of skin cancers requires certain combinations of initiators and promoters carcinogens, e.g., benzo[a]pyrene (BP), 7,12-dimethylbenz[a]anthracene (DMBA), 3-methylcholanthrene (3-MC) TPA – 12-O-tetradecanoylphorbol-13-acetate, a skin irritant prepared from the seeds of croton (巴豆) oil also called PMA (phorbol-12-myristate-13-acetate) Figure 11.28 part 1 of 2 The Biology of Cancer (© Garland Science 2007) (a mutant) from (E): The effects of promoter seem to be reversible, thus, it may exert a nongenetic effect on the papilloma cells. from (F): Additional treatments of the promoter seem to push the papilloma into a promoterindependent state. Figure 11.28 part 2 of 2 and 11.29 The Biology of Cancer (© Garland Science 2007) Explanation of the combined effect of initiation and promotion After 40 years, mutagen drive the proliferation of ras* cells The skin tumors that emerge invariably bear point-mutated H-ras oncogenes, indicating that this mutation confers a strong selective advantage on cells in the skin. Figure 11.30 The Biology of Cancer (© Garland Science 2007) Why does TPA (a tumor promoter) promote tumor formation? TPA acts as a functional mimic of diacylglycerol (DAG), which activates protein kinase C. Figure 6.16 The Biology of Cancer (© Garland Science 2007) DAG activates protein kinase C α (PKCα ) Figure 11.31 The Biology of Cancer (© Garland Science 2007) Induction of inflammation by TPA wild type TPA was initially chosen because it is an irritant of mouse skin and thus an inducer of localized inflammation. Figure 11.36 The Biology of Cancer (© Garland Science 2007) PKCα transgenic mice (keratin-5-promoter driven) neutrophil infiltration 11.14 - 17 Toxic agents, mitogenic agents and chronic inflammation can act as tumor promoters cytotoxic agents : damage cells → causing the proliferation of the cells that have survived the toxic effects mitogenic agents : i.e.,steroid hormones Estrogen and progesterone are involved in the programming the proliferation of cells in reproductive tissues chronic inflammation – tissue damage from inflammation – long-term anti-inflammatory drug use links to reduced cancer incidence Sidebar 11.14 Hepatitis B virus infections lead to hepatocellular carcinoma (HCC) in Taiwanese government employees 1975: 22,707 men – 3,454 were HBsAg + 19,253 were HBsAg – 1986: 153/3,454 HBsAg + men died of HCC 9/19,253 HBsAg – men died of HCC The relative risk of dying from HCC if one carries HBsAg in the blood is 98.4. Chronic inflammation leading to cancer Inflammatory cells Inflammatory cells Figure 11.35 The Biology of Cancer (© Garland Science 2007) B型肝炎病毒的慢性感染為何 會演變成肝硬化和肝癌? 病毒感染 對抗病毒的 免疫反應 肝臟再生 肝細胞損傷 B型肝炎病毒的慢性感染為何 會演變成肝硬化和肝癌? 病毒感染 對抗病毒的 免疫反應 肝臟再生 肝細胞損傷 DNA 突變 ↓ 肝癌 Fibroblast 修補 ↓ 肝硬化 強度不足 Table 11.3 The Biology of Cancer (© Garland Science 2007)