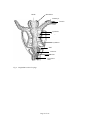
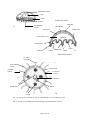
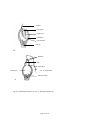
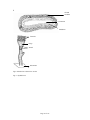
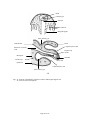
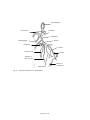
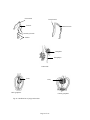
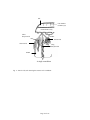
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Animal Diversity- I (Non-Chordates) Phylum Cnidaria (Old name- Coelenterata) Dr. Sukanya Lal Zoology Department, Ramjas College, University of Delhi Delhi – 110 007 26th June 2007 Page 1 of 115 Chapter index I. Phylum Cnidaria (Coelenterata): General characters 1. Introduction 2. Morphology 3. Reproduction II. Classification of Phylum-Cnidaria (Coelenterata) 1. 2. A. B. C. D. Introduction Classification Class: Hydrozoa Class Scyphozoa Class: Cubomedusae Class: Anthozoa I Subclass: Alcyonaria (Octocorallia) II. Subclass: Zoantharia (Hexacorallia) III. Obelia geniculata 1. Habit and Habitat 2. Structure A. Polyp B. Blastostyle C. Medusa 3. Histological structure of zooids A. Epidermis B. Mesogloea C. Gastrodermis / Endodermis 4. Statocyst 5. Gonads 6. Locomotion 7. Feeding 8. Excretion 9. Respiration 10. Reproduction A. Asexual mode of reproduction B. Sexual mode of reproduction 11. Alternation of generation and Metagenesis 12. Polymorphism. A. Polyps B. Blastostyle C. Medusa Figures 1-10 Page 2 of 115 IV. Aurelia aurita (Jelly-fish) 1. Introduction 2. Habit and Habitat 3. Structure A. Oral arms and Mouth B. Radial canals C. Velarium D. Sub-genital pits E. Gonads 4. Histological structure A. Epidermis B. Mesogloea C. Gastrodermis 5. Nervous System 6. Sense organs 7. Locomotion 8. Water circulation 9. Food and feeding 10. Digestive system 11. Respiration and Excretion 12. Reproduction 13. Life Cycle A. Planula larva B. Scyphistoma larva C. Strobilation D. Ephyra Larva E. Metamorphosis 14. Alternation of Generation Table 1 Figures 1-9 V. Polymorphism 1. Introduction 2. Class- Hydrozoa A. Order- Hydroida a. Hydra b. Obelia c. Bougainvillea d. Tubularia e. Hydractinia f. Vellela g. Porpita B. Modifications of polyp C. Modifications of medusa D. Order- Siphonophora Page 3 of 115 a. Physalia b. Diphyes c. Helistemma 3. Class Anthozoa a. Pennatula (Sea Pen) Figures 1-11 VI. Mesenteries 1. Introduction 2. Structure of Metridium (Sea Anemone) explaining Mesenteries A. Primary mesenteries or complete mesenteries B. Secondary mesenteries or incomplete mesenteries 3. Octocorallians 4. Hexacorallians 5. Examples of anthozoans showing different arrangement of mesenteries a. Alcyonium b. Edwardsia c. Gonactinia d. Halcampoides e. Halcampa f. Adamsia g. Haloclava h. Zoanthus i. Epizoanthus j. Cerianthus k. Antipathes (Black coral) l. Peachia m. Metridium 6. Formation of mesenteries 7. Significance and function of mesenteries Figures 1-17 VII. Corals and coral reefs 1. Introduction 2. Class: Anthozoa A. Subclass: Octocorallia or Alcyonacea a. Structure of Octocorallian coral i. Alcyonium (Dead man’s finger or soft coral) ii. Heliopora (blue coral) iii. Tubipora (Organ pipe coral) iv. Corallium (Red coral) v. Gorgonia (Sea fan) Page 4 of 115 B. Subclass: Hexacorallia (Zoantharia) a. Structure and formation of a Hexacorallian coral i. Fungia (Mushroom coral) ii. Madrepora (Horn coral) iii. Astraea (Star coral) iv. Meandrina (Brain coral) v. Astrangia (white coral) vi. Antipathes (Black coral) 3. Class: Hydrozoa i. Millepora (Fire coral or sting coral) 4. Coral reefs A. Development of coral reefs B. Types and structure of Coral reefs a. Fringing reefs b. Barrier reefs c. Atoll C. Theories explaining the formation of coral reefs a. b. c. d. e. Darwin’s Dana subsidence theory Stutchbury’s volcanic crater theory Samper Murray solution theory Submerged bank theory Daly glacial control theory 5. Significance of corals 6. Coral crisis A. Natural factors affecting coral growth a. Temperature b. Depth of the ocean c. Availability of light and presence of algae d. Sea storms B. Man made factors a. Increase in human population and development b. Aesthetic value c. Sea traffic d. Predators C. Coral Bleaching D. Coral Diseases Figures 1-18 VIII. Bibliography IX. Acknowledgements Page 5 of 115 I. Phylum Cnidaria (Coelenterata): General characters 1. Introduction: The phylum coelenterata is a Greek word where- ‘coel’ and ‘enteron’ stand for hollow and intestine respectively. It is also known as Cnidaria because of the presence of a unique organelle called nematocyst or cnidae (formed by special cells called cnidoblasts). The phylum Coelenterata contains more than 10,000 species, including jelly fish, fresh water Hydra, box jellies, sea anemones, soft coral and hard coral forming animals. Cnidarians are Eumetazoans (multicellular) with primarily radial symmetry. They constitute the lowest group of animals among the Eumetazoa. Cnidarians can be distinguished from sponges (see phylum- Porifera) as they have a distinct digestive cavity called coelenteron. They differ from ctenophores (see PhylumCtenophora) by having nematocysts, a polypoid stage and reproduce both sexually and asexually. 2. Morphology/ Anatomy: These fascinating animals are basically gelatinous in composition. A majority of them are marine water living except fresh water Hydra and Craspedacusta. Most of the animals are characterized by radial symmetry about an oralaboral axis. The body presents tissue level organization i.e. cells are organized into tissues which can perform various functions within an organism. The body wall of a cnidarian is diploblastic i.e. composed of only two layers of cells, the ectoderm and the endoderm. A layer of mesogloea generally intervenes between the ectoderm and the endoderm. Although the mesogloea itself is nonliving and gelatinous, it may contain living cells derived from embryonic ectoderm. Amoebocytes in the mesogloea of anthozoans probably play roles in digestion, nutrient transport and storage, wound repair and antibacterial defense. Phylum Cnidaria can be divided into four classes: the Hydrozoa (Hydra, Obelia), the Scyphozoa (jelly fish), Cubomedusae (box jellies), and the Anthozoa (sea anemones, gorgonians, sea pens, corals etc.). Cnidarians may have two basic body forms- polyps and medusae (also called zooids) which may be modified into different forms performing different functions and mutually benefiting each other. Hydrozoans bear polyps or medusa or both, adult of scyphozoans and class Cubomedusae are only medusoid forms, while all anthozoans are polypoid forms. Polyps are generally cylindrical, and are found attached to the main colony or to the substratum while medusae are free living. In polyps the mouth is surrounded by tentacles and faces upwards i.e. away from the substratum. In contrast, medusa is umbrella like, having ex-umbrellar and sub-umbrellar surfaces with the mouth prolonged centrally into a manubrium and facing towards the bottom. Tentacles are present on the periphery of the sub-umbrellar surface. They freely swim in water by rhythmic contractions. Their orientation is inverted in relation to that of the polyps. In all cnidarians, the mouth leads into a spacious cavity called the coelenteron. The coelenteron is enclosed by two–layered body wall (diploblastic condition) consisting of outer ectoderm and inner gastrodersm. Coelenteron is also called the gastrovascular cavity as it acts both as a digestive system and a circulatory system for the circulation of food, oxygen, excretory wastes and many other materials with in the body. It is lined by gastrodermis (also known as the endodermis) which is sometimes infolded to form radial septa or mesenteries. These mesenteries increase the surface area over which digestion may take place, because the primary function of the endoderm is digestion. Cnidarians Page 6 of 115 are mainly carnivorous although some soft corals have been found to feed also on phytoplankton. Colonial forms are fixed, so they cannot move actively from one place to another. They capture their prey with the help of tentacles encircling their mouth. Special stinging cells or cnidoblasts unique to this phylum are present abundantly on the tentacles and at the terminal ends of the mesenteries in anthozoans. Three general types of cnidoblasts are present in different cnidarians. Cnidoblasts having a cnidocil and nematocyst, which is a common type of stinging cells, is present in hydrozoans and scyphozoans. Besides this majority of anthozoans have spirocysts or ptychocysts. Spirocyst, as its name suggests, contains a spirally coiled thread inside. This thread, on discharge, releases sticky material. Spirocysts are present in most of the anthozoans in addition to nematocysts. Similarly, ptychocysts contain a tubule, which is haphazardly arranged inside the capsule, and is released. Ptychocysts are present in the tube dwelling anemones like Cerianthus. These can be discharged with great force for a variety of functions. Around 30 different types of cnidoblasts are reported to be present in different cnidarians. Many cnidarians have symbiotic photosynthetic algae i.e. zoochlorallae (in Hydra) and zooxanthellae (in marine cnidarians) within their gastrodermal cells or in their ectodermal cells. They live symbiotically with these algae and obtain additional nutrients from them. They benefit algae in turn by providing CO2 and metabolic products for photosynthesis. The nervous system is very primitive and lacks a central nervous system. In contrast it consists of a network of nerve cells and nerve processes which generally synapse on one another repeatedly and terminate at a neuromuscular junction. This type of a nerve network helps in immediate spread of excitation over the entire body of the animal when sensory cells are stimulated. As a result, the animal can change its orientation or react according to the situation. Muscle layers are derived from epitheliomuscular cells and endotheliomuscular cells that possess elongated, contractile bases anchored in the mesogloea. Circulatory, excretory or respiratory organs are completely lacking as the functions of these systems are performed by the gastrovascular cavity. A majority of the animals belonging to the class hydrozoa exhibit the phenomenon of polymorphism, which is defined as the occurrence of structurally and functionally different forms within the same organism during its life cycle. Different types of individuals are called zooids e.g. Obelia has polyps called feeding zooids or gastrozooids, while free living reproductive zooids are called medusae. Many anthozoan polyps secrete a calcareous skeleton made up of calcium carbonate (externally or internally), forming exoskeleton or endoskeleton respectively. This helps in deposition of calcareous skeletons called corals which helps in the formation of massive deposits in the sea called coral reefs. 3. Reproduction: Reproduction in cnidarians occurs asexually by budding, fragmentation or by pedal laceration, while sexual reproduction takes place by the sex cells produced by gonads. In some animals like Hydra, which have only a polypoid stage, gonads are developed from the epidermal cells to reproduce sexually in addition to the asexual reproduction by budding. Aurelia exists only as medusa; while Obelia has a characteristic life cycle, in which polyp and medusa alternates with each other during their life cycle. Medusa being a reproductive zooid carries four gonads, one of which is located along each of the radial canals. When the gametes mature, they are released by Page 7 of 115 rupturing of the gonad wall. Sperms fertilize the ova in the sea water and the zygote leads to the formation of a two layered, characteristic larval stage called the Planula larva. Planula larva has a ciliated ectoderm and freely swims in the water for some time before it settles and develops into a polypoid form. This polypoid form is tubular; bears the mouth and tentacles directed upwards and are attached to the substratum aborally. It multiplies asexually and either produces other polyps, which may lead an independent life to form a colony, or gives rise to free living medusoid stage bearing gonads. Hydrozoans thus exhibit a phenomenon of metagenesis i.e. an alternation of asexual and sexual phases during its life history. II. Classification of Phylum-Cnidaria (Coelenterata) 1. Introduction: i. They exhibit radial symmetry and are acoelomates (without coelom). ii. Tissue level (cells are organized into tissues and perform different functions) of organization is achieved in this lowest group of animals among the true metazoans. iii. Body wall is diploblastic (it is made up of two layers of cells, an outer ectoderm and inner gastroderm with a thin layer of mesogloea secreted by both these layers). iv. Special stinging cells or cnidoblasts, unique to this phylum are present. v. A single gastrovascular cavity (coelenteron), opening by a mouth (single opening) that serves as a mouth and anus. vi. Nervous system is in the form of a net work of nerve cells and nerve fibers present in the mesogloea. vii. Organism may be a sedentary (fixed) polyp, free living solitary medusa, or may bear both polyp and medusa in the same colony. viii. Asexual reproduction takes place by budding or fragmentation. ix. Sexual reproduction produces a characteristic, free living, ciliated planula larva. x. A phenomenon of Polymorphism is exhibited in hydrozoans and anthozoans.. xi. Many organisms form corals and help in the formation of coral reefs (a unique aquatic ecosystem). 2. Classification: A. Class: Hydrozoa: i. A majority of the hydrozoans are colonial and found in marine water except Hydra and few others which are fresh water living animals. ii. In solitary and colonial forms, polyps are predominantly nutritive zooids. Page 8 of 115 iii. Sexual zooids are called medusae and are produced by asexual budding. These are umbrella shaped and free swimming iv. Gonads are ectodermal and are borne by the medusa. However, in Hydra like forms, these are borne by polyps as they lack medusoid stage. v. Medusa possesses a circular shelf within the umbrella called velum. Hydrozoans are classified into following orders: a. Order: Trachylina i. All marine. ii. Contains most primitive animals of the class hydrozoa. iii. No polypoid stage, only medusoid zooids present. iv. Bell margin smooth. v. Gonads on radial canals or attached to manubrium. vi. Sense organs are tentaculocysts (modified tentacles) which are partly mesodermal in origin. Ocelli usually absent. vii. Planula larva develops into an actinula larva which directly develops into a medusa without intervening polyp stage. Examples: Aglaura, Liriope, etc. b. Order: Hydroida i. Solitary or colonial. ii. Well developed polypoid generation without gastric ridges. iii. Medusoid stage present or absent. iv. Velum is present in the medusa. v. Sense organs of medusa are ocelli and statocysts which are ectodermal in origin. Suborder 1: Limnomedusae i. Majority are fresh water animals. ii. Small solitary polyps present and medusae bud off from the sides. Examples: Craspedacusta Suborder 2: Anthomedusae (Athecate) i.Solitary or colonial hydrozoans. ii.Bell shaped, free medusa are common. iii.Polyp athecate (not surrounded by skeletal covering). iv.Statocyst absent. Page 9 of 115 Examples: Eudendrium, Hydractinia, Tubularia, Pennaria etc. Suborder 3: Leptomedusae (Thecate) i. Colonial hydrozoans. ii. Polyps are thecate i.e. they are enclosed by hydrotheca, and medusa produced by blastostyles is covered by gonotheca. iii. Free medusae are generally absent, and when present, are umbrella shaped and bear statocysts for equilibrium. Examples: Obelia, Companularia, Sertularia, Plumularia etc. Suborder 4: Chondrophora i. Pelagic, free swimming forms. ii. Polymorphic polypoid colonies. iii. These animals can be considered as large, single, inverted polyps. Examples: Vellela (float thin, with erect sail), Porpita (float disc shaped). c. Order: Actinulida i. Very small, solitary hydrozoans like actinula larva. ii. Medusoid stage is completely absent. Examples: Otohydra, Halammohydra. d. Order: Siphonophora i. Marine, pelagic, especially in warm seas. ii. Free swimming hydrozoan colonies comprising of polypoid and medusoid zooids. iii. Upper end of colony usually bears a supporting float or swimming bells. iv. Nematocysts many, large and powerful. v. Medusae incomplete, attached to stem or disc, rarely free. vi. Gonads in gonophores, which are not set free. Examples: Physalia (Portuguese Man of war, float inflated), Stephalia, Nectalia. e. Order: Hydrocorallina i. Colonial, polypoid hydrozoans. ii. Polyps of 2 forms, gastrozooids and dactylozooids. iii. Hydromedusae consisting of coenosarcal canals, ectoderm of which secretes a hard iv. Calcareous skeleton filling up the spaces of coenosarcal mesh work. v. Rudimentary medusae in the form of gonophores are generally present. Defensive and feeding polyps located within star-shaped openings on the skeleton. Page 10 of 115 Example: Millepora. B. Class Scyphozoa: i. All marine jelly fishes. ii. Chiefly free swimming medusae, bell or umbrella form, with strong 4 per radial symmetry and much gelatinous mesogloea. iii. No true velum, no stomodeum. iv. Gastric ridges present about mouth. v. Central gastrovascular stomach and radial canals with complex branches are present. vi. Notches in bell margin with sense organs rhopalia (tentaculocysts) having endodermal statoliths. vii. Medusae sexual, dioecious with endodermal gonads in pouches of the gastric cavity. viii. Polyp generation none or reduced (hytratuba and scyphistoma) producing medusae directly or by transverse fission. a. Order: Stauromedusae i. Sessile polypoid forms attached by a stalk on the aboral side of the trumpet shaped body. ii. Marginal sense organs (rhopalia) absent but 8 simple tentacles present. iii. Four cornered mouth with small oral lobes and manubrium is present. Examples: Haliclystus: Lucernaria etc. b. Order: Coronatae i. Free swimming scyphozoans found in warm waters. ii. Body cubical with four flat sides. Bell surrounded by circular furrow, above scalloped margin. iii. Four hollow inter-radial tentacles, four rhopalia at per radial positions. iv. Found mainly in deep waters. Examples: Periphylla, Nausithoe, Linuche, Atolla etc. c. Order: Semaeostomeae i Saucer-shaped bells having scalloped margins bearing tentacles. ii Mouth central, corners prolonged as four oral frilly arms. iii Gastrovascular cavity extends from central stomach to radial canals and circular canal at the bell margin. iv Chiefly in coastal waters Examples: Aurelia, Cynea, Pelagia, Aurelia, Stygiomedusa etc. Page 11 of 115 d. Order: Rhizostomeae i. With 8 large adradial, root like simple or branched oral arms with numerous suctorial mouths ii. Original central mouth is lost because of fusion of oral arms except in Stomolophus. iii. Gastrovascular cavity without septa. iv. Eight or more rhopalia (tentaculocysts), four are per-radial and 4 ad-radial. Examples: Cassiopea, Rhizostoma, Mastigias, Stomolophus. C. Class: Cubomedusae: Medusoid forms, with bell cubical and margin bent inward. i. Known as box jellies with complex eyes and potent toxin. ii. Tentacles 4 or in 4 groups. iii. Feed mostly on fish. iv. Planula larva develops into a fixed polypoid larva which metamorphoses into medusa. v. Found in tropical and subtropical waters, shores and open seas. Examples: Carybdea, Chironex, Chiropsalmus etc. D. Class: Anthozoa: All marine, solitary or colonial, includes sea anemones, corals and sea pens etc. i. All polypoid forms, sessile (fixed), solitary or colonial. ii. Oral disc flat, with hollow tentacles. iii. Mouth leading into stomodeum (gullet) lined by ectoderm, usually with ciliated groove (siphonoglyph) in the pharyngeal wall leading from mouth. iv. Enteron divided by vertical septa (mesenteries) bearing nematocysts on inner margins. v. Nematocysts do not possess an operculum like hydrozoans and scyphozoans and are called spirocysts or ptycocysts. vi. Mesogloea a connective tissue. vii. With or without skeleton. viii. Gonads (endodermal) in septa or mesenteries. ix. Planula larva develops from the fertilized egg and metamorphoses to form another polyp. x. Many anthozoans reproduce asexually by longitudinal or transverse fission. xi. Pedal laceration (in which parts of the pedal disc detach from the rest of the body and form a new individual) is also present. xii. Majority forms corals. Page 12 of 115 I Subclass: Alcyonaria (Octocorallia) i. With 8 pinnately branched tentacles and 8 single complete mesenteries or septa. ii. Modular colonies and are often polymorphic. iii. One central siphonoglyph is present. iv. Soft corals, polyps are supported by calcareous endoskeleton of spicules secreted by cells in the mesogloea. a. Order: Stolonifera i. Polyps arising separately from common stolon or mat. ii. Skeleton of separate calcareous spicules. iii. Sometimes fused as tubes. iv. Clavularia in California coast. v. Tubipora musica, organ pipe coral, in warm waters on coral reefs. Examples: Clavularia, Tubipora etc. b. Order: Telestacea i. Colony consists of simple or branched stems bearing lateral polyps ii. Skeleton consists of calcareous spicules. Example: Telesto. c. Order: Alcyonacea (soft corals) i. Polyps with lower parts fused in a fleshy mass and only oral ends protruding out. ii. Polyps are dimorphic in some forms bearing autozooids and siphonozooids. iii. Skeleton of separate limy spicules. iv. Found mostly in warm shore waters. Examples: Alcyonium, Gersemia, Anthomastus, Sarcophyton etc. d. Order: Helioporacea i. Skeleton massive made up of calcareous fibers. ii. Commonly known as blue corals. Examples: Heliopora, blue coral of Indo-Pacific region. e. Order: Gorgonacea i. Colony usually plant-like arising from a basal disc. ii. Axial skeleton of calcareous spicules made up of horn like gorgonin or both. iii. Polyp short. Examples: Corallium (red coral), used for jewelry; Gorgonia (Sea fan) Page 13 of 115 f. Order: Pennatulacea i. Colony fleshy, of one long axial polyp, with many dimorphic polyps arranged on both sides present on bare stalks. ii. Skeleton of limy spicules. Examples: Pennatula (Sea pen) feather like; Renilla (sea pansy), disc shaped; Umbellula. II. Subclass: Zoantharia (Hexacorallia) i. Tentacles few to many (usually in the multiples of six) and 6 pairs of primary mesenteries. ii. Many species are solitary and lack any protective covering (sea anemone), some are colonial but never polymorphic(true stony corals or scleractinian corals) iii. True stony corals secrete hard calcium carbonate exoskeleton. They may be reef building (hermatypic) or not (ahermatypic). iv. Siphonoglyph 1 or 2, associated with the pharynx. v. Many have endosymbiotic relationship with unicellular, photosynthetic zooxanthellae. a. Order: Zoanthidea i. No skeleton or pedal disc. ii. Polyps usually united by basal stolons. iii. Some solitary with stalked base iv. Many species on exterior of various invertebrates. Example: Zoanthus, Epizoanthus on hermit crab, Parazoanthus. b. Order: Actinaria i. All sea anemones. ii. No skeleton, polyps of same size, columnar with muscular wall and usually a pedal disc. iii. Stomodeum usually with siphonoglyph. iv. Mesenteries paired, often in multiples of 6. v. Found on rocks, on sand, or on invertebrates. vi. Sessile, but not fixed. vii. Essentially solitary. viii. Filaments with ciliated areas Examples: Halcampoides, Metridium, Anthopleura, Adamsia on hermit crab shells, Edwardsia in burrows. c. Order: Madreporaria or Scleractinia (Stony corals). Page 14 of 115 i. Exoskeleton compact, calcareous, polyp small or minute in cups on skeleton. ii. Tentacles in the multiples of six. iii. No siphonoglyph. iv. Muscles feeble. v. Mostly colonial in warm seas. Examples: Fungia (solitary), Acropora, Astrangia, Oculina. d. Order: Corallimorpharia i. No ciliated areas on filaments. ii. With capitate tentacles. iii. Usually in radial series. Examples: Corynactis, Ricordea. e. Order: Ceriantharia i. Solitary, slender, elongate, anemone like, living in vertical tubes. ii. Many tentacles in 2 circles. iii. No pedal disc, one siphonoglyph. iv. Solitary. Example: Cerianthus inhabits slime-lined vertical tubes in sea bottom and take out their tentacles and oral disc from the tube at the time of feeding. f. Order: Antipatharia (Black corals) i. Skeleton plant like, stems (some branched) composed of horny material and bear small polyps. ii. Tentacles 6 in number. iii. Found in deeper tropical waters. Example: Antipathes Page 15 of 115 III. Obelia geniculata (Sea fur) 1. Habit and Habitat: A typical example of class hydrozoa is Obelia geniculata. It is a colonial hydrozoan, commonly known as sea fur and is found attached to rocks, wooden piles, shells or seaweeds. The colony grows by budding from a single hydra-like individual. It consists of individuals (zooids) of three different forms-polyp, blastostyle and medusa attached to vertical branches. There are many polyps grown on the branching colony with the blastostyles grown in the axils of lower branches of the colony. Each blastostyle is a clubbed shaped extension of the main colony which by asexual budding gives rise to many small saucer shaped medusae. Each medusa is a free living form of the colony. It provides a means of dispersal and is able to reproduce the colony sexually. Obelia is one of the important cnidarians as it exhibits the phenomenon of polymorphism and alternation of generation also called as metagenesis. 2. Structure: Obelia is a microscopic, delicate and fur like colonial animal. It is whitish or brown in color. Its colony consists of zigzag stems arising from horizontal, hollow, branched, root like filaments called Hydrorhiza (Fig. 1). It forms a complicated meshwork over the surface of the substratum and helps in fixing the colony to the substratum (sedentary or fixed nature). From the horizontally running hydrorhiza, several, vertical branches arise called hydrocauli (hydrocaulus singular). Each hydrocaulus forms the main stem of a colony and is of 2 to 3 cm in height. It grows upright having alternate branching system. Each branch terminates into an individual (also called zooid) of a colony called polyp or a gastrozooid. Gastrozooid is so named as its main function is to feed the colony. Some of these branches may end up in the form of small buds representing young forms of the polyps. From the axils of each older branch arise cylindrical zooids, called blastostyle which are generally present towards the lower regions of the colony. Blastostyles, by asexual budding give rise to sexual zooids called medusae. Medusae are called sexual zooids because each one of them bears four gonads which are located on the per-radial position. When a medusa gets mature, it is released free from the blastostyle and swims freely in the water to carry out the process of sexual reproduction. The entire colony of the Obelia including its branches and zooids are covered by a chitinous external skeleton called perisarc and the innermost layer is called coenosarc. Coenosarc encloses a cavity within called coenosarcal canal. Basically, coenosarc consists of an external epidermis and an internal layer of gastrodermis with a thin sheet of noncellular mesogloea sandwiched in between them. As coenosarcal canal is continuous with the zooids and the colony, digested food, oxygen and even water is circulated and distributed through it to all the parts of the colony. Perisarc is tough, transparent, noncellular and cuticular covering. It is secreted by the epidermis which is present beneath it. It provides rigidity and physical strength to the colony. Perisarc was in direct contact Page 16 of 115 with the coenosarc in the beginning of development but gets separated from the coenosarc by a space, except at certain points at regular intervals. Because of the rigidity of the perisarc, colony and its branches cannot move except at certain points where circular rings or annuli formed by perisarc are present. The perisarc surrounding the polyp is called hydrotheca while surrounding the blastostyle is called gonotheca. A. Polyp: Polyp is also called as hydranth. Majority of the individuals of the Obelia colony are polyps. These are meant for the feeding of the colony and thus known as gastrozooids (Fig. 1 and 2). Number of polyps is more in Obelia colony as compared to blastostyle. Polyps resemble Hydra structurally. These are present at the terminal ends of almost all the branches of the colony. One end of the polyp is connected to the hydrocaulus while its free end is produced into conical structure, the manubrium (hypostome), which is around one third of the length of the polyp. Manubrium bears a small opening in the center called mouth. Mouth leads into a big cavity called gastrovascular cavity which is continuous within the body of polyp and with the coenosarcal canal of the hydrocaulus. Around the base of the manubrium are present numerous tentacles in a single circlet. Tentacles are long, pointed, and are projected beyond the manubrium. They bear batteries of nematocysts in their ectodermal cells. In Obelia tentacles are solid because of the presence of a single row of cylindrical and highly vacuolated endodermal cells. In contrast, tentacles of Hydra are hollow. Perisarc present on the outer surface of the polyp expands and forms a loose, protective conical, vase like structure around the polyp called hydrotheca. It is transparent and has a smooth outer margin. At the base of the polyp, hydrotheca is produced into a ring like horizontal platform or shelf. At the time of danger, polyp can retract suddenly by folding its tentacles within the hydrotheca and shelf checks the polyp from being pulled within the perisarc of the hydrocaulus. Perisarc forms three to four annuli at the base of the hydrotheca surrounding the polyp. These annuli help in swaying movements of the polyp to capture the prey. The basal disc of the polyp is continuous with the coenosarc of the stem to which it is attached. Though, each polyp is an individual, all the polyps are connected together by a tubular coenosarcal canal, through which the coelenteron continues. B. Blastostyle: Blastostyle is a zooid which asexually produces numerous medusa buds from its upper end. Blastostyles are produced in the axils of the vertical branches (hydrocauli). These are normally present towards the basal (older) branches of the colony (Fig. 1 and 3a). The number of blastostyle is less as compared to polyps in a colony. Every blastostyle consists of a central stem arising from the coenosarc. It is devoid of mouth and tentacles and has a reduced gastrovascular cavity, it cannot feed on itself, however, nutrients are supplied through the coenosarcal connection with the gastrozooid. It is enclosed in a long, cylindrical, transparent covering of perisarc called as the gonotheca. Gonotheca has a small opening, the gonopore towards its terminal end through which medusae can come out. Blastostyle with the gonotheca around is called Gonangium. The basal end of gonangium has two or three annular rings which provide flexibility to it as in the case of polyps. When medusae buds are fully formed, they are released out through gonopore into the surrounding water to lead a free swimming life. Page 17 of 115 C. Medusa: Medusa is a gonozooid which is produced by asexual budding from blastostyle. Mature medusae are present towards the terminal end of the blastostyle and younger ones are present towards the base of the blastostyle (Fig. 3b, c, and d). Each medusa carry four gonads, each one is located along each of the four per-radial canals on the subumbrellar side. When the gonads become ripe, they shed gametes by rupturing their wall. Medusae are produced from blastostyles in large numbers during spring and summer season. Medusa has an umbrella like structure, with the convex, exumbrellar surface and a ventral subumbrellar surface. The edge of the umbrella is produced inwards as a short double fold of integument to form a diaphragm like structure called velum. A medusa having velum is called craspedote and when velum is absent, it is called acraspedote (Aurelia). The rim of the medusa bears highly contractile tentacles with batteries of nematoblasts. A young medusa has only sixteen tentacles and the number increases as it grows. From the subumbrellar surface of the medusa arises a short hollow structure called manubrium bearing a four sided mouth at its terminal end. Manubrium appears like the handle of the typical umbrella. Medusa has a radial symmetry. If a circle is drawn with a four sided mouth in the center, then four radii arising from the angles of the mouth are named as per-radial positions.. The tentacles present at the per-radial positions are called per-radial tentacles. Bisecting the angle between two per-radii is known as inter-radial position. Radius passing through per-radial position and inter-radial position is named as ad-radial position. The tentacles present at inter-radial and the ad-radial position are called inter-radial and ad-radial tentacles respectively. A young medusa has four per-radial, four inter-radial and eight ad-radial tentacles. The mouth leads into a short gullet in the manubrium followed by a stomach occupying the central position of the subumbrellar surface of the medusa. The stomach continues into four thin, delicate radial canals which run towards the periphery of the medusa and are placed at equal distance from each other (Fig. 3d). At periphery of the medusa, the four radial canals open into a circular canal which runs all along the margin of the medusa. In medusa gastrovascular cavity is thus restricted to gullet, stomach, radial canals and circular canal. 3. Histological structure of zooids: The basic histological structure of polyp, blastostyle, medusa and coenosarc are similar and resembles Hydra. Therefore, the body wall of Hydra is discussed here briefly to understand the histology of the zooids. The body wall is made up of two layers of cells and is diploblastic. The outer and inner layers are known as ectoderm (epidermis) and endoderm (gastrodermis). A thin, delicate, transparent and noncellular material called mesogloea secreted by both epidermis and gastrodermis lies in between these layers. Medusa has an outer epidermis, and innermost gastrodermis with a thin layer of intervening mesogloea. Both exumbrellar and subumbrellar surfaces are covered with epidermis. The whole canal system is lined by a thin sheet of gastrodermis called as gastrodermal core, which is formed by the fusion of an upper and a lower layer of gastrodermis. Velum is composed of a double layer of epidermis enclosing a thin layer of mesogloea, but without gastrodermis. Tentacles are solid because of the presence of Page 18 of 115 vacuolated gastrodermal cells that are covered externally by epidermis bearing batteries of nematocysts. Exumbrellar surface (as it always faces up) has a very thick gelatinous mesogloea present between epidermis and gastrodermis and forms the main bulk of the body. Mesogloea contains lot of water and noncellular fibers. Muscular regions of the epitheliomuscular cells are better developed and are arranged in circular, longitudinal and radial tracts forming a well formed musculature. Tentacles have swollen regions at their bases because of the presence of interstitial cells. Interstitial cells are totipotent as they give rise to formation of any type of the cells depending upon the requirement of the individual tissues. These cells also replace cnidoblasts or nematoblasts (cells bearing nematocysts) which die soon after the discharge of the nematocyst. Nematoblasts are quite numerous on the margin of umbrella, tentacles and around the mouth. Nematocysts borne by nematoblasts help in paralyzing the prey and also protecting the colony by keeping away the enemies (see details below). Nerve cells and fibers are concentrated around the margin of the cell forming two nerve rings, inner and outer. The ex-umbrellar or outer nerve ring supplies the tentacles, while the subumbrellar nerve ring or inner nerve ring supplies the subumbrellar musculature and the statocysts. Inner nerve rings around the bell margin is associated with the rhythmic contractions of the medusa. A. Epidermis: Epidermis is thin and is made up of many different types of cells which are structurally and functionally differentiated (Fig.4). a. Epithelio-muscular cells: Each epithelio-muscular cell has outer broader end and narrow inner ends prolonged into unstriped muscle fibers. These cells are so named as they form the outer epidermis on one side and have muscular ends on the inner side, which are arranged parallel to the long axis of the body. Their contractions make the body and tentacles shorter and thicker. b. Sensory cells: Sensory cells are distributed all along the external surface of the body in between the epidermal cells. Their number is more on the tentacles and manubrium. They bear a delicate hair like sensory cilia at their free ends while their inner ends are withdrawn into fine fibers which meet the nerve net present below the epidermis. Sensory cells receive and transmit impulses from and to the nerve cells. These are sensitive to touch, temperature and chemicals and accordingly modified to perform different functions. c. Cnidoblast (nematoblast): Some of the cells of epidermis are specialized for the protection of the colony. These cells are called the cnidoblasts or nematoblasts and are specifically present only in coelenterates. Cnidoblasts are also known as stinging cells. These cells are aggregated in more numbers in the epidermis of the tentacles and such regions are called batteries of nematocysts (Fig. 5a). i. Structure: Each cnidoblast is a large cell enclosing a pear shaped cell called nematocyst. Its nucleus lies on one side of the cell. Nematocyst has a central cavity containing poisonous fluid, the hypnotoxin and is closed with a lid called operculum. Poisonous fluid or hypnotoxin is a mixture of proteins and phenol. The outer end of the nematocyst invaginates into a funnel shaped structure tapering into a coiled thread like Page 19 of 115 filament folded like a spring inside the cavity (Fig. 5b), the tip of the thread may be open or closed but the base is swollen to form the butt or shaft. There are three large spines on the butt; pointing inwards are called stylets or barbs. In addition, three spiral rows of minute spines called barbules or spinnerets are also present. There is few supporting muscle fibers arising from the basal region of the cnidoblast and are attached to the nematocyst. A small, pointed, sensory hair like structure called cnidocil is present close to the operculum and is projected outside beyond the epidermis. ii. Function: Cnidocil is sensitive to vibrations in the water and when a microorganism or prey passes close to the cnidarians, it stimulates the nematocyst to discharge (Fig: 5c). When this happens, the central cavity containing hypnotoxin is intensely compressed by the muscle fibers and the operculum opens up immediately. As a result, the coiled thread opens out with a great force turning inside out. As the thread is shot out, barbs and barbules penetrate the prey so that the hypnotoxin is injected and the prey is killed by the poison. Batteries of nematocysts are found on the tentacles, which all discharge together and paralyze or kill the enemy or prey. Nematocysts thus help not only in protection from enemies but also in capturing food. Some nematocysts can produce a burning sensation, itching or even death to higher animals. iii. Types of nematocysts: There are different types of nematocysts (Fig: 6a) present in different cnidarians performing different type of function. In fact all of them are known as stinging cells. - Penetrant type: It is the largest type of the cnidoblast and is present in Hydra. The tip of the thread is open and penetrates the tissues of the prey and injects poisonous fluid once it is discharged (Fig. 6b). - Volvent type: It is pear shaped. The tip of the thread is closed and is mainly used to hold the prey by coiling around it. - The Glutinants: They are of adhesive types and used by Hydra to stick its tentacles to the substratum when the Hydra is moving by looping. - The Streptoline Glutinant: It is oval and its thread bears minute spine like structures and they get coiled on the prey, once they are discharged. - The Stereoline glutinant: They discharge a straight unarmed thread. All these types of the nematocysts are discharged only once and then these cells die and are replaced by new cnidoblasts formed by interstitial cells. Spirocysts and ptychocysts are some of the other types of nematocysts which are present only in anthozoans (sea anemones and coral forming animals). - Spirocysts: Sea anemones bear spirocysts in their ectoderm and endodermal cells in addition to nematocysts. In their tentacles spirocysts are more in number as compared to nematocysts. Spirocysts consists of capsule with a single wall and a long thread. They help the anemone to capturing the food organisms that have hard surfaces e.g. small crustaceans attached to the substratum. -Ptychocysts: Large anemones like Cerianthus and Zoanthus which are solitary and tube dwelling, bear a nematocyst like organelle in their ectoderm. These animals are adapted to stay in the tubes lined by mucous and discharged threads and capsules of ptychocysts. Page 20 of 115 d. Interstitial cells (Totipotent cells): These cells are located in between the adjacent epitheliomuscular cells. These are small rounded cells having prominent nuclei with little cytoplasm. These cells are also called totipotent cells as they can differentiate into any type of cell depending upon the requirement of the body tissues. e. Nerve cells: There are both epidermal and gastrodermal nerve nets in cnidarians. Nerve cells are located beneath the epitheliomuscular cells and musculo-nutritive cells and lie parallel to both the layers. Each nerve cell consists of a nucleated cell with two to many processes called nerve fibers. These nerve cells are similar to multipolar neurons of higher animals but are very primitive in nature as they appeared for the first time in the coelenterates. The nerve cells form an irregular nerve net and synapse with other nerve cells. The network of nerve cells and fibers is dense in the region of the oral disc and the tentacles. In addition, nerve cells are in contact with the basal parts of the sensory cells of the epidermis and gastrodermis. f. Gland cells: These cells secrete mucus which helps in adhesion, food uptake and protection. In Hydra, their number is more at the basal disc for the attachment and for locomotion. g. Sex cells: During breeding season, interstitial cells of epidermis give rise to testis or ovary. Epidermal gonads are present only in the class hydrozoa. In scyphozoans and in anthozoans, gonads originate from the gastrodermis or endodermis. B. Mesogloea: It is very thin in Hydra and Obelia colony except medusa where it is thin only on the subumbrellar surface and very thick on the exumbrellar surface of the medusa which faces upwards. Nerve nets related to epidermis and gastrodermis are located towards the mesogloea. C. Gastrodermis / Endodermis: Gastrovascular cavity is lined throughout by the endodermis in hydrozoans. In Anthozoans, gastrodermis extends in the form of folds called mesenteries or septa with a thin sheet of mesogloea within. Types of cells present in the gastrodermis are: a. Musculo-Nutritive cells: Gastrodermis consist of long, columnar cells called musculo-nutritive cells. Some of the cells bear flagella to maintain a constant circulation of fluid and food within the gastrodermal cavity and within the coenosarcal canal. There are certain cells which form pseudopodia like structures to ingest food particles by phagocytic activity. Intracellular digestion within these cells completes the digestive process. Near the manubrium, these cells are modified into circular muscles to close and open the mouth. b. Gland cells: These are present within the gastrodermis, which secrete digestive enzymes within the gastrovascular cavity for the extracellular digestion of food. c. Nerve cells: These cells are like epidermal nerve cells and form a nerve net underneath the gastrodermis parallel to the mesogloea. d. Cnidoblasts: These cells are absent in the gastrodermis but some of them are also present in gastrodermis of mesenteries in anthozoans. 4. Statocyst: There are eight receptor organs i.e. statocysts located at the bases of eight ad-radial tentacles on the subumbrellar surface, just inside the margins of the bell (Fig. Page 21 of 115 3d). Each statocyst consists of a small fluid-filled sac which contains a movable, round particle of calcium carbonate, called statolith or otolith. Statolith is secreted by a large cell, the lithocyte. The statocyst is covered by a layer of epithelial cells which are sensory in nature and connected at the base with the nerve cells (Fig. 7a and b). The other free ends of these sensory cells have protoplasmic processes which touches the statolith. These help in maintaining the body position of the medusa in a right position while floating in the water, i.e. exumbrellar surface up and subumbrellar surface down. Besides there are two circular nerve rings; nerve ring I (exumbrellar nerve ring) and nerve ring II (subumbrellar nerve ring) which are simply the aggregations of the nerve cells and are present around the margin of the medusa. Nerve ring I supplies the nerves to the tentacles while nerve ring II supplies nerves to the subumbrellar musculature and the statocysts. When medusa is swimming and the body gets tilted, the statolith falls away from the processes of sensory cells. With the result nerve impulse is created in the sensory cells and transmitted to the nerve ring II and then to the nerve net present nearby. Epitheliomuscular cells of the stimulated side contract immediately and the medusa is brought back to its original position. Statocysts control the swimming movements of the medusa in the water and thus help in maintaining equilibrium by muscular coordination. 5. Gonads: Medusa is a sexual zooid as it bears gonads for the sexual reproduction (Fig. 3c and 3d). There is no sexual dimorphism between male and female medusa but it is certain that one medusa can behave either as male and will produce only sperms while another medusa acts as female by producing ova. Testis and ovaries are borne by different medusae, so they are dioecious. Each medusa bears four gonads which are located in the middle of each radial canal on the subumbrellar surface, so the position of gonads is per-radial. They can be easily noticed as bulging on the subumbrellar surface almost at equal distances between the manubrium and the velum (Fig. 3e). Once the medusa is released from the blastostyle through gonopore of gonotheca, the gonads get matured afterwards. Each gonad is an ovoid body with an outer epidermis which is present in continuation with the epidermis of the subumbrellar surface. It has an inner layer of gastrodermis which is continuous with that of the radial canal. The space between two layers is filled with mesogloea containing interstitial cells which may either differentiate into sperms or ova at the time of maturity. Germ cells are originated from the epidermis of the manubrium when it was attached to the blastostyle. Later on, these cells are transferred through gastrodermis into the gullet and then reach the gonads. When the gonads mature, epidermis ruptures and the sperms and ova are released in the water respectively. Sperms are minute and flagellated with the help of which they actively swim in the water. While the ova are large rounded cells which may either be released in the water for fertilization or may remain within the gonads and are fertilized by the incoming sperm there itself. 6. Locomotion: As Obelia colony is sedentary (fixed), so it cannot move from one place to another. There are annuli present at the base of the polyp and blastostyles which help them to undergo sideways movements and provide little flexibility to the colony. The polyps and their tentacles can undergo contraction and relaxation due to the epitheliomuscular cells present in the epidermis. Medusa is a free living zooid, once it is released from the blastostyle; it floats in the water with its exumbrellar surface upwards and subumbrellar surface facing down. It actively swims by the muscular action which Page 22 of 115 brings about opening and closing of the bell. This propels the water behind and the animal moves forward. They also float in the water passively by the water currents and winds blowing over the sea. At times their body may be disturbed or tilted with upside down or turned inside out. Statocysts present at the periphery help in maintaining the balance and position of the medusa. 7. Feeding: Polyps are called gastrozooids as they feed the whole colony. They are carnivorous and feed on microorganisms like nematodes, worms, crustaceans, small fish etc. The food is captured with the help of tentacles bearing batteries of nematocysts. Nematocysts kill the potential food and tentacles get the food within the mouth. Gland cells present in the gastrodermis release digestive juices within the gastrovascular cavity which break the food into smaller pieces and partly digest the food. Partly digested food is taken inside the musculo-nutritive cells and digested within the cells. Digested food is absorbed and circulated throughout the colony by the flagellated cells present in the gastrodermis. Medusa being free swimming can feed them, once they are released in the water. In medusa, the food is captured by the highly extensible mouth and then taken into gullet. From the gullet it is transferred to the stomach where digestive enzymes are released and digested food is then circulated through radial canals into the circular canal. 8. Excretion: There are no special organs for excretion. Nitrogenous wastes diffuse out into the water through external as well as internal surfaces. As there is only one opening i.e. mouth in both polyp and medusa, wastes released inside are thrown out of the mouth. 9. Respiration: Exchange of gases takes place through general surface of the body as the colony is present within water. As water also enters through mouth in the gastrodermal cavity and is circulated throughout the colony, each cell is in direct contact with gases dissolved in water. Oxygen diffuses inside and the carbon dioxide diffuses out in the water which moves out of the colony through mouth. 10. Reproduction: A. Asexual mode of reproduction: Obelia colony grows in size by budding of new horizontal stems (hydrorhiza) and vertical stems (hydrocauli), thereby increasing the number of zooids. Blastostyles form medusae (reproductive zooids) by budding. B. Sexual mode of reproduction: Medusa is a free swimming reproductive zooid which is formed by budding from the blastostyle. Medusa, once sexually mature is set free and released out through gonopore of gonangium into the water. A medusa is dioecious i.e. it bears either male or female gonads. There is no sexual dimorphism (male medusa and female medusa cannot be identified externally). Each medusa bears four gonads at the per-radial position on the subumbrellar surface. Each gonad has an outer ectoderm and inner gastrodermis. Ectoderm of the gonad is in continuation with the ectoderm of the subumbrellar surface while gastrodermis of the gonad is in continuation with the gastrodermis of the radial canals. The germ cells do not arise in the gonads but they develop from the interstitial cells of the ectoderm of the blastostyle at the time of budding of medusa. Later on these germ cells migrate through radial canals and take their position within the gonads. Gonads either give rise to sperms or ova. Mature Page 23 of 115 sperms are shed into the sea water by the rupture of gonads. Ova may be either released in the water or they may remain within the gonads to be fertilized. Medusa dies once it releases sex cells in the water. a. Fertilization: Fertilization may take place in the water if both ova and sperms are released in the water or sperms may fertilize the ova within the gonads of female medusa. b. Life cycle: A single sperm fertilizes one egg to form the zygote. Zygote undergoes complete (holoblastic) and equal cleavage to form a solid ball of cells, called morula. It develops into another stage called blastula with a cavity within called blastocoel. Two layered gastrula is formed by a process of delamination (from the inner surface of the blastomeres are cut off new cells which fill up the blastocoel at one pole). Thus the hollow blastula changes into a two layered gastrula with an outer ectoderm and inner endoderm. i. Planula larva: Gastrula becomes elongated and its outer ectoderm gets uniformly ciliated (Fig. 8), it has at first an inner solid core of endodermal cells which eventually split to develop a cavity called coelenteron. This larva has a distinct broad anterior end and a narrow posterior end. Its cells get differentiated into columnar ectodermal, sensory, gland, nerve cells and nematocysts. Nerve cells and sensory cells are present in great number at the anterior end. As planula larva is ciliated, it leads a free living life for sometime but it cannot feed on its own as it has no mouth. After a short span of free living, it settles down, attaches to the substratum which can be a stone, log of wood, or any solid object. After settling down, it undergoes metamorphosis and changes to next larval stage called hydrula stage (polypoid form). ii. Hydrula larva: Its attached end forms the basal disc while its free end develops a mouth on a manubrium with number of tentacles surrounding it. This larva resembles a miniature Hydra and thus called a hydrula larva (Fig. 9). Hydrula grows further by extensive process of budding of hydrocaulous and hydrorhiza and is soon converted into a complex branching colony of Obelia like parental colony. While free swimming medusa in the life cycle of a fixed Obelia colony helps in the dispersal of gametes and it is free swimming planula larva which disburses the species to newer locations. This doubly ensures that species do not over crowd in one area. 11. Alternation of generation and Metagenesis: In Obelia, two types of individuals, the fixed polyp and the free swimming medusa alternate successively so that polypoid generation asexually gives rise to medusae, which reproduces sexually to produce zygotes developing later on into polypoid form of larva. This hydrula larva develops into a new colony by asexual budding (Fig. 10). But it is not true alternation of generation as reported in plants which are discussed below. A. Alternation of generation: True alternation of generations is better explained in a plant (fern). Here the fern is diploid and known as sporophyte. It reproduces asexually producing haploid spores. Each spore develops in a haploid generation called gametophyte. The gametophyte produces haploid ova and spermatozoa which after fertilization, give rise to the diploid generation called sporophyte. So, the life cycle of a fern shows alternation of an asexual, diploid, sporophytic generation with the sexual, haploid gametophytic generation. This is known as true alteration of generation. Page 24 of 115 B. Metagenesis: In Obelia, both colonial, hydroid phase and free living medusoid phases are diploid. Medusa represents diploid phase because it originates from the diploid blastostyle by budding. The sexual generation is represented only by the haploid gametes which are in fact derived from the epidermis of the blastostyle ad then migrated to gonads of medusa. So medusa is not producing the gametes, but is meant only for dispersal of gametes. Therefore, in Obelia it is diploid hydroid colony which alternates with the free swimming diploid medusa. This type of alternation of generation is called metagenesis. Obelia thus does not show the true alternation of asexual phase with the sexual phase as mentioned in fern earlier. In brief, metagenesis is the regular alternation of the colonial, asexual, diploid, polypoid generation with the solitary, free moving, diploid, sexual, medusoid generation. 12. Polymorphism. : It is the phenomenon of occurrence of several structurally and functionally different individuals within a species. Any species having different individuals performing different functions for its survival and dispersal is called polymorphic. It results in the formation of a colony bearing different individuals with division of labor. Colony remains fixed to the substratum or may be free living with certain different modifications. Obelia is one of the very important organisms which exhibit the phenomenon of polymorphism as it bears three distinct types of zooids. A. Polyps: These are feeding zooids or gastrozooids, and are meant for the feeding of the whole colony. Food captured by gastrozooids is shared with other zooids via the common coelenteron present within the colony. B. Blastostyles: These are asexual polyps and produce medusae by budding from them. Once the medusae are fully formed, they are released out of them through gonopore. C. Medusae: These are diploid reproductive zooids and reproduce sexually by producing haploid sperms or ova in the water. They lead a free living life and help in dispersal of gametes and prevent overcrowding of the species. Page 25 of 115 Tentacles extended Polyp Tentacles contracted Hydrotheca Blastostyle Polyp retracted Gonopore Gonotheca Gonangium Medusa bud Perisarcal annuli Perisarc Hydrocaulus Coenosarc Hydrorhiza Fig. 1: External features of an Obelia colony. Page 26 of 115 Mouth Manubrium Nematocyst Tentacle Epidermis Mesogloea Endoderm Hydrotheca Shelf Coenosarc Perisarc Coenosarcal canal Fig. 2. Longitudinal section of a polyp. Page 27 of 115 Gonotheca Medusa . Gonopore Gonangium 3a Gonopore Gonotheca Medusa Blastostyle Coenosarcal canal Annuli 3b Fig. 3a. A Gonangium arising from the colony of Obelia. 3b. A gonangium showing blastostyle and budding medusa. Page 28 of 115 Exumbrellar surface Gonad Velum Exumbrellar surface Tentacles Epidermis Subumbrellar surface 3c Gastrodermis Mesogloea Radial canal Gastrodermal cavity Stomach Circular canal Tentacle Velum Manubrium Mouth Gonad Subumbrellar surface Per-radial tentacle Inter-radial tentacle Circular canal Ad-radial tentacle Location of statocyst Manubrium Mouth Radial canal 3d Gonad Fig. 3c: Lateral view of a medusa. Fig. 3d: Subumbrellar view of medusa (oral view) Fig. 3e. Lateral view of medusa showing positioning of gonads and other structures. Page 29 of 115 3e Gastrodermis Epidermis Gland cell Sensory cell Interstitial cells Epithelio-muscular cell Endothelio- gland cell Cnidoblast Germ cell Interstitial cells Sensory cell Nutritive cell Nerve cells Mesogloea Fig. 4: L.S. of Body wall of Hydra. Page 30 of 115 Discharged nematocyst Epidermis Fig. 5a: Part of the epidermis showing cnidoblasts. Page 31 of 115 Cnidocil Operculum Nematocyst Cnidoblast Coiled thread Nucleus 5b Barbules Barb Operculum Nematocyst Everted thread Muscular fibers 5c Fig. 5b: Undischarged nematocyst. Fig. 5c. Discharged nematocyst. Page 32 of 115 Penetrant Volvent Glutinant Undischarged Nematocysts Discharged Nematocysts Fig.6a: Types of nematocysts Page 33 of 115 Penetrant nematocyst Enlarged view of the tail region of this larva Penetrant Volvent Fig. 6b. An insect larva paralyzed by penetrant nematocysts and volvents. Page 34 of 115 Circular canal Ectoderm Gastroderm Mesogloea Interstitial cells Tentacular base Outer nerve ring 7a Inner nerve ring Statocyst Sensory cells with processes Otolith Lithocyte with a nucleus 7b Fluid filled vesicle Fig. 7a: Statocyst at the base of an adradial canal in L.S. 7b. Structure of a single statocyst in a section. Page 35 of 115 8 Ciliated ectoderm Coelenteron Endoderm Tentacles Polyp Annuli Substratum 9 Fig.8: Planula Larva shown in a section. Fig. 9. Hydrula larva Page 36 of 115 Female medusa (2n) Obelia colony (2n) Male medusa (2n) Ovum (n) Sperm (n) Zygote (2n) 2-cell stage Hydrula larva (2n) Life cycle of Obelia Blastula (2n) Young polyp Gastrula forms by delamination Longitudinal section (L.S.) Ciliated Planula Fig. 10. Life cycle of Obelia Page 37 of 115 IV. Aurelia aurita (Jelly-fish) 1. Introduction: The class scyphozoa is represented primarily by the medusoid form, which is more dominant phase in their life history whereas the polypoid stage is restricted only to the larval stage which is known as scyphistoma. Aurelia aurita is a typical example of Class - Scyphozoa (which means ‘cup like animals’) and is commonly known as jelly fish or sea jelly. There are around 200 species of jelly fishes and all of them are found in marine water. The name jelly fish or sea jelly is assigned to Aurelia aurita because its body is transparent due to jelly like massive mesogloea present between epidermis and endodermis. It does not have any resemblance to fish (which is a highly evolved vertebrate) except that it is a water living animal. There is also no relation between two as both of them have entirely different structures. 2. Habit and Habitat: Aurelia aurita is marine water living animal and is the inhabitant of coastal waters in warm and temperate seas. It may remain solitary (live alone) or in groups floating quietly by rhythmic contractions. They are thrown by sea currents on the sea shore during storms. 3. Structure: Aurelia is a medusoid form; however it differs from the medusa of Obelia in many respects (Table 1). For instance oral arms while present in Aurelia, are absent in a typical hydrozoan medusa. Sensory organs of Obelia are more advanced (rhopalium) than statocyst of hydrozoan medusa. Aurelia has a transparent body and has a radial symmetry (Fig. 1a). It has an inverted cup like body which is gelatinous in nature. Size may vary from 7 cm. to about 60 cm. however; some are quite large and may be 2 meter across. It has slightly convex exumbrellar surface and concave subumbrellar surface and its body can easily be identified by the presence of four horse-shoe shaped gonads in the centre of the subumbrellar surface a little away from the manubrium. The body parts can be shown with reference to particular radii of the bell or cup. A. Oral arms and Mouth: A very short manubrium is present in the centre of the subumbrellar surface which bears a mouth at its tip (Fig. 1b). There are four frilled and delicate oral arms arising from the four angles of the mouth. The four arms are almost as long as the radius of the bell and their broad bases are united to surround the mouth. Each oral arm bears longitudinal, ciliated grooves leading into the mouth. The thin edges of the grooves are convoluted, and bear numerous short tentacles (fimbriae) with batteries of nematocysts, but the base is thick. Cross section of an arm is ‘V’ shaped. B. Radial canals: The corners of the mouth and the four radial canals arising from them are per-radial in position (Fig. 1b and Fig. 2). Midway between four per-radial canals are 4 inter-radial canals. Per-radial and inter-radial canals are branched and rebranched before joining the circular canal. Further, there is one ad-radial canal present in between Page 38 of 115 each per-radial and inter-radial canal giving rise to total 8 ad-radial canals. The body is roughly circular in outline interrupted with eight notches placed at the 4 per-radial and 4 inter-radial positions. Each notch hosts a sensory organ called tentaculocyst or rhopalia protected by a pair of small lobes called marginal lappets. C. Velarium: There is a thin, delicate, flexible, marginal flap called velarium which is a continuation of the sub-umbrellar surface and bears tentacles and 8 sensory organs called rhopalia all along the margin. It has gastrodermal canals running through it. It is different from the velum of the medusa of Obelia because velum is only a narrow fold of subumbrellar ectoderm and there are no canals running through it in contrast to velarium. D. Sub-genital pits: Sub-genital pits are four in number and lies just beneath the gonads and are inter-radial in their location. These are small rounded apertures which lead into a small shallow cavity. They do not have any special function in relation to gonads. E. Gonads: There are four gonads. Each gonad is present just above the sub genital pits at the inter-radial position. They are horseshoe-shaped structures and are visible through jelly like body. 4. Histological structure: The arrangement of body wall layers and cell types are similar to that of Obelia or Hydra. A. Epidermis: The exumbrellar and subumbrellar surfaces, the tentacles, velarium, oral arms and manubrium are covered externally by epidermis (Fig. 3). The gullet and the sub-genital pits are also lined by invaginated epidermis. Exumbrellar surface bears epithelial cells while sub-umbrellar surface has epithelio-muscular cells in addition to nerve, sensory, gland and stinging cells (cnidoblasts). Epithelio-muscular cells of the subumbrellar surface are striated while muscles of the tentacles, manubrium and the oral arms are simple longitudinal muscles. B. Mesogloea: It is sandwiched between epidermis and gastrodermis. It is highly gelatinous and forms a very thick sheet on the exumbrellar surface. It contains branching fibers and amoeboid cells derived from the epidermis. C. Gastrodermis: The gastrovascular cavity (stomach), radial canals originating from stomach and their extensions into the rhopalia or tentaculocysts, circular canal at the periphery etc. are all lined internally by the gastrodermis. Gastrodermis consists of flagellated endothelial cells, gland cells and nerve cells. 5. Nervous System: Nervous system of Aurelia is more advanced than that of medusa of Obelia. It consists of (A) main nerve plexus present on the subumbrellar surface, and (B) a diffuse nerve net present both on exumbrellar and subumbrellar surfaces. A. Main nerve plexus: Main nerve plexus constitutes a rapid conducting system of stimulus through bipolar cells. It is present below the ectoderm on the subumbrellar surface and helps in coordination of muscular activity during locomotion. It is also extended in the oral arms, manubrium, tentacles and rhopalia. It is made up of bipolar nerve cells and nerve fibers arising from them forming a continuous net work. These nerve cells form special thickening at the base of tentacles present at the per-radial and inter-radial positions. This ganglion like concentrations of nerve cells coordinates with the rhopalia present near them. Sensory impulses received by the sense organs are Page 39 of 115 conveyed to the muscle fibers and mesogloea to control the movements of the jelly fish by them. B. The diffuse nerve net: The diffuse nerve net constitutes a slow conducting system of multipolar neurons. It is present in ectoderm of both exumbrellar and subumbrellar surfaces. The nerve cells are smaller with multipolar nerve fibers and help in recognition of local stimulus and animal can respond accordingly. 6. Sense organs: Aurelia aurita freely swims in the water showing periodic movements from surface waters to deeper waters and back again. They also form temporary breeding aggregations. At times they are drifted upside down with the water currents. To maintain themselves in the right orientation, they have developed special sensory organs like rhopalium for maintaining equilibrium, ocelli-to respond to light and olfactory pits- sensitive to smell. All these structures coordinate with each other to give a proper response. A. Rhopalia (Tentaculocysts): They are modified tentacles and hence called tentaculocysts. There are eight characteristic sensory organs called rhopalia. Out of which four are located at per-radial and four are at inter-radial position. Each rhopalia is located between a notches formed by a pair of specialized tentacles called marginal lappets and is protected over by hood like process of bell margin (Fig. 4a, 4b). Each rhopalium consists of a hollow club shaped projection enclosing an extension of circular canal lined by gastrodermis. At the apical end of gastrodermal cells, a spherical mass of cells called statolith is present (Fig. 4b). Its function is just like the statocyst of the Obelia. Statolith is made up of calcium sulphate and calcium phosphate particles. There is a region of ciliated sensory cells (adoral olfactory pit explained a little later) in the epithelial layer just beneath the tentaculocyst. If the animal is tilted, the tentaculocyst containing statocyst press against the ciliated sensory cells present below, with the result a stimulus is generated in the associated nerve cells of main nerve plexus present on the subumbrellar surface. Tentaculocyst makes the animal aware of its displaced orientation and coordinates the muscular activity and the animal comes to its original position. So rhopalium act in coordination with the sensory cells present nearby and maintains the equilibrium of the jelly fish by controlling the rhythmic action of the swimming bell. B. Ocelli: Two types of ocelli are present in Aurelia. One is pigment spot ocellus and the other is pigment cup ocellus (Fig. 4b). a. Pigment spot ocellus is exposed on the outer side of the epidermal cells of the tentaculocyst. It consists of photoreceptor cells packed with light sensitive pigment and is connected with the nerve net present underneath. b. Pigment cup ocellus is situated on the inner side of the tentaculocyst in association with the statocyst. It is in the form of an inverted cup made by pigmented and sensory gastrodermal cells. The pigment cup ocellus is connected with the underneath nerve net. Both these ocelli (Pigment spot ocellus and Pigment cup ocellus) are simply light receptors which help the animal to come to the surface or go down in the water according to the availability of light. Both these ocelli are non image forming light receptors. C. Olfactory pits: Olfactory pits are the regions lined with sensory cells present in the epidermis. There are two olfactory pits, outer or aboral olfactory pit is present on the Page 40 of 115 exumbrellar surface at the base of the hood, and the inner or adoral olfactory pit is present below the tentaculocyst. 7. Locomotion: Locomotion in Aurelia is brought about by the sudden rhythmic contractions of the muscles and gelatinous mesogloea. Mesogloea functions as a skeletal system relaxing the muscles after their contraction. When the muscle fibers of the jelly fish contract, the volume of the fluid under the bell decreases, with the result water is forcibly expelled from under the bell and the animal is propelled in the opposite direction. Due to contraction of muscles, the gelatinous mesogloea is displaced. When the muscles relax, mesogloea comes back to its original form and the volume of water enclosed by the swimming bell increases and the jellyfish is pushed downward. 8. Food and feeding: Aurelia is carnivorous and its food consists of planktons, eggs, and larval stages of many marine invertebrates. Sometimes they also derive nutrition from zooxanthellae (unicellular algae) that live in symbiotic relationship with them. Feeding in Aurelia may takes place (i) by using oral arms and tentacles: as the jelly fish swims and the prey comes in contact with the tentacles or oral arms, both these structures contract and bring the prey near the manubrium and then taken within the mouth for further digestion or (ii) they may behave as suspension feeders by trapping planktons with the help of mucus on the subumbrellar surface. The mucus laden food is then swept towards the bell margin, from where it carried by flagellated grooves within the oral arms towards the mouth and then to the stomach. 9. Digestive system: Digestive system here is better known as a system of fluid-filled gastrovascular canals. Gastrovascular canals help in the circulation of oxygen and carbon dioxide (vascular part), as well as in the circulation of food (gastro part) and hence called gastrovascular system. Gastrovascular system consists of mouth, gullet, stomach, gastrogenital canal, gastric pouches, adradial canals, circular canal, per-radial and inter-radial canals, exhalent canal, gastro-oral canal, basal groove of oral tentacle. Nematocysts present on the oral arms or tentacles paralyze the food particles and carry into the mouth. Food is then carried to four gastric pouches bearing gastric filaments (Fig. 2 and 3). Gastric filaments bearing nematocysts paralyze or kill and digest the prey with the help of digestive enzymes they secrete. Partly digested food is then conveyed to the stomach. Gland cells present within the gastrodermal cells release enzymes for extracellular digestion while intracellular digestion takes place by phagocytosis of the food particles within the gastrodermal cells. Digested food is then circulated through radial canals by the ciliated gastrodermal cells, while undigested food is thrown out of the mouth by exhalent current of water. The path of food circulation is explained in the water circulation. 10. Water circulation: Circulation of water plays an important role in the distribution of food and oxygen in Aurelia. Inhalent current of water enters the mouth, passes through the narrow gullet into the stomach and then to gastric pouches. Each gastric pouch contains an opening to help circulation of the water. From gastric pouches, water enters through the unbranched ad-radial canals and reaches the circular canal (Fig. 2). The exhalent water current returns by the branched inter-radial and per-radial canals back to gastric pouches. Per-radial canals bring water current to gastro-oral canal and then to the stomach, while inter-radial canals bring water to exhalent canal, to gastro-oral canal and Page 41 of 115 then to stomach. From the stomach water goes out through basal groove tract in the oral arms. 11. Respiration and Excretion: The highly branched gastrovascular cavity and constant circulation of water brings in oxygen along with food while carbon dioxide and excretory wastes (ammonia) are carried away from the body either by outgoing current of water or diffuses out through general surface of the body. 12. Reproduction: Asexual mode of reproduction is completely absent in Aurelia. Sexual reproduction takes place by sex cells produced in the gonads. Sexes are separate i.e. sperms and ova are produced by different individuals. Sexes cannot be differentiated externally (no sexual dimorphism). There are four horse-shoe shaped gonads in each sex, which are present at the per-radial position (Fig. 1b). Gonads develop within gastrodermal tissue unlike Obelia where gonads develop from the ectodermal tissue. Gonads are closely associated with the gastric pouches and their (gonads) concavities face towards the stomach. Mature sperms from male Aurelia are released from gonads into the stomach and passed out through mouth by outgoing current of water. While a mature ovum is discharged from the ovary, get fertilized by incoming sperm inside the gastric pouch (internal fertilization). 13. Life Cycle: After fertilization, zygote reaches the oral groove through water current. Oral groove acts as the temporary brood chamber as further development takes place there (Fig. 9). By repeated divisions, zygote develops into a ball of cells called morula. Morula changes into a single layered blastula with a cavity inside called blastocoel. Blastula undergoes a process of invagination and a two layered gastrula is formed. Gastrula has an outer ectoderm and inner endoderm. The gastroderm lines a central cavity called coelenteron having a small opening to outside called blastopore which is not completely closed. It is in contrast to Obelia where gastrulation takes place by delamination. Gastrula elongates, its outer ectoderm develops cilia and blastopore gets closed. Larva at this stage is named as planula larva (Fig. 5a, 5b). These larvae can be seen lodged on the oral arms of the female Aurelia. Planula larva: Once Planula larva is fully developed, it escapes out of the oral arms and freely swims in the water with the help of cilia. Planula larva cannot feed on its own as it does not have any mouth. Therefore, after swimming for sometimes it settles down and attaches itself to substrate, loses its cilia and a mouth is being developed at the opposite end of attachment. Soon tentacles are developed around the mouth. Its attached end develops into a basal disc. In this way planula larva is being transformed into a small Hydra like polypoid larval stage called hydratuba or young scyphistoma larva (Fig.6a). B. Scyphistoma larva: It is the only polypoid stage in the life cycle of Aurelia. It is attached to the substratum. Scyphistoma larva is sessile (fixed) like Obelia colony. It has no statocyst and ocelli like adult Aurelia. Its attached end elongates and narrows down to a basal disc. Its free end develops a squarish mouth and its edges elongate to form manubrium. Firstly, four per-radial tentacles bud out around mouth. Subsequently, other four inter-radial and then eight ad-radial tentacles are developed. So, larva has developed sixteen tentacles around its mouth (Fig. 7). It starts feeding on its own. Transverse section of a scyphistoma larva shows the presence of a diploblastic body wall (Fig. 6b). The gastrodermis lining the coelenteron is raised into four per-radial gastric pouches Page 42 of 115 alternating with four gastric ridges or taenioles. These gastric pouches and gastric ridges partially divide the gastrovascular cavity. Each gastric ridge bears rudimentary gonads, septal muscles and a sub-umbrellar funnel in the centre. All four gastric ridges bear small gastric filaments hanging towards the gastrovascular cavity. The gastric ridges are present only in the larval stage of Aurelia and are absent in the adult. Presence of gastric ridges is one of the characteristic features of the scyphozoans and these are considered as preliminary evolution of mesenteries present in anthozoans. As scyphistoma larva feeds on its own, it grows in size. It either undergoes a process of strobilation or it may multiply asexually by process of budding and can form new hydratuba which separates from the parent attaches to the substratum and grows into another scyphistoma. C. Strobilation: Scyphistoma undergoes a process of strobilation. During strobilation, larva stops feeding. Strobilation is a process during which the body of scyphistoma subdivides transversely forming number of saucer shaped structures piled one above the other. At this stage it is known as a strobila. Each saucer shaped structure is known as ephyra larva. Ephyrae break away from the strobila one by one as they grew up and start swimming freely in the water. First ephyra larva which breaks from the parent bears the original tentacles of the scyphistoma, which falls later on. When many ephyrae are produced from single scyphistoma, it is called polydisc strobilation. Polydisc strobilation takes place when food is in plenty and the temperature is low. When the food is in scarcity and the temperature is high, only one ephyra is produced from the scyphistoma larva, it is called monodisc strobilation. D. Ephyra Larva: It is a young, medusoid form larva of Aurelia. It has an exumbrellar and subumbrellar surface. Its body has eight bifid arms; out of which four are per-radial and four are inter-radial (Fig. 8a and 8b). Each arm is notched at the terminal end to form a pair of marginal lappets. In between the marginal lappets, there is a short tentacle which later on develops into rhopalium or tentaculocyst. All oral arms are free from each other at the periphery and joined in the form of a disc like structure towards the centre. At this stage, 8 rayed ephyra acquires a star like structure. In between each arm a rudimentary bud of ad-radial canal can be seen. The ephyra contains a small segment of stomach of scyphistoma with gastric ridges. On the exumbrellar surface, stomach is closed, while on the subumbrellar side it is open. Its edges grow out to form a short manubrium bearing a squarish mouth at its terminal end. Cavity of the stomach grows into eight arms forming eight radial canals. Gastric ridges are replaced by full fledged gastric filaments. E. Metamorphosis: The ephyra freely swims in the water feeding on microorganisms. As it grows, mesogloea thickens enormously, so that two gastrodermal layers come together forming a solid lamella between the gastrodermal radial canals. The ad-radial buds grow rapidly and fill the gaps between per-radial and inter-radial arms. All sixteen radial canals are fully formed and all of them finally open in the circular canal which is developed at the periphery of the bell. Now, star shaped, 8 rayed ephyra becomes circular and saucer-shaped like the adult medusa (Fig. 9). Further, four oral arms around mouth and numerous marginal tentacles appear at the periphery of the medusa. A young medusa changes its physical appearance altogether and becomes an adult Aurelia. Page 43 of 115 14. Alternation of Generation: Aurelia reproduces sexually whereas the hydratuba and scyphistoma reproduce asexually. Aurelia is medusoid in nature and the hydratuba and scyphistoma are polypoid forms as they resemble a typical polyp structure. Therefore, it can be said that the medusoid phase alternates with the polypoid phase. But, Aurelia does not present a true case of alternation of generation, because here the medusoid phase develops as a result of metamorphosis of an ephyra which is developed as one of the several segments of scyphistoma. The life cycle represents metamorphosis complicated by asexual multiplication in the larval condition. Table 1. Comparison of Aurelia and medusa of Obelia colony. S. No. Aurelia ( a medusoid form) Medusa of Obelia colony 1. 2. Saucer shaped with exumbrellar and subumbrellar surfaces. Bell margin is notched into 8 lobes bearing 8 rhopalia present at the per-radial and inter radial positions. 3. Rhopalium endodermal Bell shaped but grows from blastostyle. Bell margin is smooth and bears 8 sense organs (statocysts) present at the base of the ad radial tentacles. Statocyst ectodermal 4. 4 unbranched radial canals are present. No ocellus present. 6. 8 branched and 8 unbranched radial canals are present. Pigment cup ocellus and pigment spot ocelli are present. Velarium is present 7. 4 oral arms are present around the mouth. Oral arms absent. 8. Absent 10. Gastric ridges bearing gastric filaments present. 4 gonads present internally at the base of gastric pouches 4 Sub genital pits are present. 12. Fertilization internal External fertilization 13. Gastrulation by invagination. By delamination 14. Planula → Scyphistoma→Ephyra→adult Aurelia Planula→Hydrula→ Obelia colony 5. 9. Page 44 of 115 Velum is present 4 gonads hang externally from the radial canals. Absent. Exumbrellar surface Gonad Rhopalium Marginal Tentacles Oral arms Subumbrellar surface Fig. 1a. External features of Aurelia as seen from the exumbrellar surface. Page 45 of 115 Tentacles Velarium Gastric filaments Gonad Branched Interradial canal Sub-genital pit Mouth Branched perradial canal Unbranched adradial canal Gastric pouch Rhopalium Fig.1b Subumbrellar view of Aurelia. Page 46 of 115 Oral arms Mouth region Gastro-genital duct Gastro-oral canal Gastric pouch Exhalent canal Gonad Per-radial canal Ad-radial canal Inter-radial canal Circulatory canal Location of Rhopalia Fig. 2. Diagrammatic representation of the gastrovascular system in Aurelia. A part of subumbrellar view is enlarged. Arrows represent the circulation of water current. Page 47 of 115 Exumbrellar surface Gastric filament Stomach Gonad Inter-radial canal Mesogloea Ectoderm Gastroderm Circular canal Hood Sub-genital pit Radial canal Gullet Mouth Subumbrellar surface Fig. 3. Diagrammatic vertical section of Aurelia showing gonads and gastric filaments. 4a Page 48 of 115 Hood Olfactory pit Statocyst Tentacles Marginal lappets Outer olfactory pit Gastrodermis Hood Pigment spot ocellus Extension of circular canal Pigment cup ocellus Mesogloea Statoliths Amoebocyte Nerve cells Epidermis Ciliated sensory cells Inner olfactory pit 4b Figs. 4a. Location of rhopalium in relation to tentacles and marginal lappets and 4b. Vertical section of rhopalium. Page 49 of 115 Blastopore not fully closed Ciliated ectoderm Mesogloea Gastroderm Enteron Ciliated ectoderm 5a 5b Fig. 5a. Planula larva. 5b.Vertical section of planula larva. Page 50 of 115 Tentacles Septal funnel pit Mouth Bud Basal disc 6a Septal muscles Gastrodermal cavity Epidermis Mesogloea Gonad Gastrodermis Gastric pouch Gastric filaments Subumbrellar funnel Gastric ridge 6b Fig. 6a. A young scyphistoma larva. 6b. Transverse section of a scyphistoma larva showing gastric ridges and other structures. Page 51 of 115 Tentacles Mouth Mature Ephyrae Young ephyrae produced by transverse fission Stalk Basal disc Fig. 7. Strobilating scyphistoma larva. Page 52 of 115 Exumbrellar surface Arms Mouth Subumbrellar surface 8a Primary Lappets Buds of adradial canal Mouth Gastric ridges Per-radial canal Rhopalium Inter-radial canal 8b Fig. 8a. Ephyra larva (lateral view). 8b. Ephyra larva (subumbrellar view) Page 53 of 115 Adult male Adult female Aurelia in a section Gonad Ephyra larva Fertilization Oral arm Sperm enters female medusa Life cycle of Aurelia Zygote 2-celled stage Blastula Gastrula forms by invagination Ciliated planula larva Scyphistoma larva Young strobila Planula larva (A section) Fig. 9. Life cycle of Aurelia aurita. Page 54 of 115 V Polymorphism 1. Introduction: The presence of polymorphism in cnidarians is one of their characteristic features. It is defined as the occurrence of structurally and functionally different types of individuals within the same organism during its life cycle. A species that exhibits this phenomenon is called polymorphic. Polymorphism is predominantly exhibited by the different animals of class- hydrozoa. Hydroid colonies which bear two types of zooids are known as dimorphic, while colonies which bear more than two types of zooids are called polymorphic colonies. There are different types of zooids which have evolved in different animals according to their habit and habitat and are discussed in this section. 2. Class- Hydrozoa: Phenomenon of polymorphism is exhibited generally by the animals of class hydrozoa but a few animals belonging to class anthozoa like Penaatula (sea pen) also show dimorphism. Degree of polymorphism can be explained from the following examples. A. Order- Hydroida: a. Hydra: Hydra is the fresh water animal in contrast to majority of cnidarians discussed below which are marine water animals. It remains singly and is attached to the weeds by its base with the mouth hanging down. Hydra is the simplest type of cnidarians having only polyp form as it has no trace of medusa either in adult form or in larval stage. As it exists only in one form and all the functions are performed by the polyp itself, it is considered as the monomorphic form. It has a cylindrical structure of which the lower end is attached to the substratum while other free end always hangs down and bears a small mouth located on the raised structure called manubrium. Mouth is surrounded by number of filiform (pointed) tentacles bearing batteries of nematocysts. Mouth leads into a central body cavity called coelenteron. It reproduces asexually by producing buds and sexually by producing sex organs developed from the ectoderm of the body (Fig. 1a, b) b. Obelia: It is an important example of the trimorphic colony. As already discussed earlier in detail, it has two basic forms of zooids, i.e. polyp and medusa. Polyp and medusa are fundamentally similar and can be derived from each other hypothetically. Although, both of them are different in structure and function (Fig. 2) the polyp form exists in 2 structurally and functionally different forms making the total number of zooids to 3 types. i. Polyp: It is sessile (fixed to the substratum) with a hydra like body attached to the main colony by narrower end. Its free end is wider and raised into hypostome that bears a mouth surrounded by tentacles. It faces upwards and carries the function of feeding the colony. Polyps are specialized for feeding and thus known as gastrozooids (Fig. 2). ii. Blastostyle: Blastostyle is club shaped and it arises as an extension of the main colony from the axils of the branches and produces medusa by process of budding. It is protected by a thin transparent perisarc called gonotheca having an opening called Page 55 of 115 gonopore. Blastostyle cannot feed on its own. Medusa when mature can come out of the gonopore to swim freely in the water. iii. Medusa: It is a free swimming zooid with an umbrella shaped body having exumbrellar and subumbrellar surfaces. It has a mouth facing downwards (present on the tubular growth called manubrium hanging down from subumbrellar surface) in contrast to polyp in which mouth faces upwards. Like hydranth, it can feed on its own for its survival until its function is over. It bears four gonads (testis or ovary) which produces either sperms or ova at the time of maturity and is responsible for sexual reproduction therefore also known as gonozooid. They normally die after reproducing the colony. Therefore, they not only help in sexual reproduction but also play an important role in dispersal of the colony. c. Bougainvillea: It is a colonial, fixed, dimorphic hydrozoan. It has creeping stolons called hydrorhiza attached to the substratum from where vertical branches arise. Each vertical branch produces polyps and medusae. Each polyp is present at the terminal end of the branch and has a conical manubrium bearing a mouth, surrounded by two circlets of tentacles. A thick, chitinous, perisarc covers hydrorhiza and vertical branches arising from it. There is no hydrotheca as is present in the Obelia. Inside the perisarc is present coenosarc which is made up of ectoderm, mesogloea and gastroderm. Small rounded buds arise from the coenosarc of the branches (Figs. 3a, b). Each bud develops into a medusa. Developed medusa has a bell like structure bearing gonads for the sexual reproduction. It breaks from the stalk at the time of maturity and swims freely. Medusae do not arise from blastostyles as they arise in Obelia. d. Tubularia: It may occur singly or in colonies fixed to the substratum in marine water. It is dimorphic as it bears two types of zooids- gastrozooid and gonophores present very close to each other. Gastrozooid is flower like, has two circlets of tentacles. Oral tentacles surrounding mouth are smaller while tentacles present near the base are longer. Reproductive zooids are called gonophores as they are degenerate medusae which hang down in the form of clusters from the base of the gastrozooid (Fig. 4). They bear both male and female medusae, so colony is dioecious. The medusae are never set free. Fertilization takes place within female gonophore. Zygote develops into planula larva which remains fixed (sessile) in the gonophore which further develops into another larval stage called actinula larva which gets free from the gonophore and freely swims in the water before it settles to metamorphose into adult Tubularia. So, Tubularia has free swimming actinula larva in contrast to many other coelenterates where planula larva is a free swimming larva. e. Hydractinia: It is a small, colonial, marine and highly polymorphic animal present in shallow waters on rocks, gastropodan shells inhabited by hermit crabs etc. When Hydractinia is found attached to the gastropodan shell occupied by hermit crab, it does not have a symbiotic relationship with the crab as it is capable of managing its food on its own but it gets an extra advantage of being mobile along with the hermit crab. Hermit crab also gets an advantage of being protected as predators will not harm it because of the nematocysts loaded on the tentacles of the Hydractinia zooids. There is an epizoic (external benefit) relationship between both of them. Hydractinia has five different types of zooids which are directly attached to the hydrorhiza which forms a brownish mat over the substratum (Fig. 5). Page 56 of 115 i. Gastrozooids: These are feeding zooids. They have a large size with a mouth and one circlet of long, capitate (knobbed) tentacles at the base of conical manubrium. ii. Gonozooids: These are reproductive zooids which are without mouth and tentacles. They are smaller than gastrozooids. They are degenerate medusa and bear a group of sac like gonophores containing eggs or sperms. They have a short, rounded hypostome which bears knob-like projections bearing nematocysts. iii. Spiral dactylozooids: These zooids are without mouth and tentacles. As their name suggests they are spirally coiled, more elongated and bear numerous nematocysts at their terminal ends. They provide protection to the colony by having stinging cells (nematocysts). iv. Tentaculozooids: These are long blunt ended zooids which are present at the periphery of the colony. They are devoid of mouth and tentacles but have clubbed knobs bearing nematocysts. v. Skeletozooids: Many thin, long, spine like structures are also present which provides support and thus forms the skeleton of the colony. f. Vellela: It has an elliptical, umbrella like body resembling a typical medusa bearing a vertical crest like structure called sail on the dorsal surface. There is a pneumatophore on the dorsal side having chitinous, concentric chambers containing air and opening through fine pores on the dorsal side. Around the margin, it bears simple tentacles called dactylozooids (protective). A big single gastrozooid with a big mouth is present in the centre on the sub-umbrellar surface (Fig. 6). Gastrozooid is surrounded by numerous gonozooids, each having a mouth at its tip. Gonozooids bud off free swimming medusae at the time of maturity. There are ectodermal canals originating from the sub-umbrellar surface and opening into the pneumatophore chambers. Gastrozooids and gonozooids open inside the gastrodermal canals, which further open into the gastrodermal cavity present inside the body. It is photosymbiotic and harbor symbiotic algae in their polyps or medusa. g. Porpita: Its structure resembles Vellela, except a sail. It has a disc like body enclosing a chambered chitinous shell containing air in it (Figs. 7a, b and c). It has a large central gastrozooid, surrounded by 3-4 circlets of gonozooids which are further surrounded by dactylozooids bearing nematocysts, arranged at the periphery of the disc. Medusae are budded off from gonozooids. It is also photosymbiotic like Vellela. B. Modifications of polyp: Polyps structurally get modified into different types of zooids according to the requirement of an individual, which are described below: i. Gastrozooids: These are feeding zooids and resemble the structure of polyp without usual tentacles. They are tubular, elongated, with a mouth facing towards the bottom of the colony. From the base of the polyp arise one or two long, contractile, hollow tentacles which bear lateral contractile knobbed branches (tentilla) at regular intervals. These knobs are the batteries of nematocysts (Fig. 8a and b). Page 57 of 115 ii. Dactylozooids: These are protective zooids. They may be also called as feelers or palpons. Structurally, they look like gastrozooids but are blind structures without any mouth. They may have a long tentacle arising from the base but unlike tentacles of gastrozooid, do not bear branches (Fig. 8a, 8b). iii. Gonozooids: They may resemble gastrozooids having mouth but are without tentacles (Fig. 8a, 8b) and bear medusa. In other the gonozooids may form stalked branches bearing grape like structures called gonophores. Sometimes tentacles like dactylozooids are attached to them which are called gonopalpons. iv. Pneumatophore: It is a hydrostatic apparatus present in siphonophores. It is gas filled chamber that appear to be a highly modified polyps (although previously considered as derived from medusae). The pneumatophore is the first zooid formed from the planula larva in those animals that possess it. It helps in keeping the body in an upright condition while floating. It is without mesogloea but the umbrella cavity contains an air chamber called a pneumatocyst (Fig. 8a). Cells lining the pneumatocyst secrete the gases or may expel out of it through one or more small openings called stigmata. Thus pneumatophore is a balloon like structure or a hydrostatic chamber containing air. It is present in those forms which have a long stem and helps in keeping the body in an upright position (Fig. 9, 10a and 11a). When pneumatophore is filled with air, the colony becomes lighter and floats at the surface of the water, but when the gas is expelled out of the pneumatophore, colony sinks down. C. Modifications of medusa: Medusoid zooids are being modified into much different type of structures which help the colony to swim (Fig. 8a and b). These may be of following types : i. Nectocalyx (nectocalyces-plural): These are small swimming bells, resembling medusa in having radial canals, circular canal and velum with a deeply concave muscular subumbrellar surface. They do not bear mouth, tentacles and sense organs, probably because their main function is to make the colony lighter, so that these colonies can swim freely and easily (Fig. 8a). They move the colony through water by their contractions. ii. Nectophore: It is another kind of modified medusa which is present in Diphyes. It is a large conical swimming bell having a groove into which whole colony can be retracted inside (Fig. 10a). It helps in swimming of the colony as Diphyes does not have any separate pneumatophore for swimming. iii. Hydrophyllum or bract: Hydrophyllum as its name suggests, is a small leaf like, protective structures modified from umbrellas of medusa but having a reduced gastrovascular cavity and have a cleft on one side. In many forms, these are much bigger in size. Bract on the other hand, is a small leaflet and is much smaller than the hydrophyllum. Bracts are attached to each cormidium (Fig. 10b, 11b). They partially cover the cormidium and help them in protection and in floating. Page 58 of 115 All above mentioned different type of zooids are originated as buds from the main axis of the stem and are lined by ectoderm and endoderm. Their cavities communicate with the cavity of the stem. D. Order- Siphonophora: This order exhibit highest degree of polymorphism. It includes complex, floating, polymorphic colonies produced by budding from an originally medusoid or polypoid form. Here, Polyps and medusae are modified into different zooids which remain attached to the same colony and perform specific functions. They do not possess any sensory organ found in other floating organisms like Aurelia and medusa of Obelia. The characteristic feature of this order is the presence of swimming bells, nectocalyces or a pneumatophore (an air filled balloon like structure) meant for the floating of the colony. The structure of a generalized siphonophore exhibits different types of modifications of polyp and medusa (Fig. 8a). Examples of few animals are discussed below to explain polymorphism. a. Physalia: It is commonly called as Portuguese man-of- war because it can suddenly appears and disappears from the surface of water by increasing or decreasing the gas content within its pneumatophore. It possesses a large pneumatophore (a modified polyp lying horizontally) on the dorsal side filled with air to float in the water. There is a small opening at the front end of the pneumatophore which leads into a chamber called pneumatocyst (Fig. 9), the air secreting cells are present at the base of the pneumatocyst. The main component of the gas within the float is around 90 percent carbon monoxide and rest is oxygen. The float has a crest formed by a fold of the trunk on the dorsal side. The pneumatocyst extends into the crest and is divided by a number of transverse septa into air chambers. A group of zooids arise from the ventral side of the float in a multiple series. Each group of zooid arising from the same stalk is called a cormidium (pluralcormidia). Each cormidium bears short and long dactylozooids (without mouth), gastrozooids (with mouth), and clustered gonozooids. Long dactylozooids sometimes may reach a length of 6 feet and have especially large stinging cells (nematocysts) that can paralyze the prey immediately and may cause burning sensation, followed by fatal injury even to humans. b. Diphyes: It consists of a long contractile hollow stem bearing at the apex two opposed nectocalyces or swimming bells without manubrium, but with four radial canals, a circular canal, and velum. At regular intervals a series cormidia present. At the point where the stem joins two nectocalyces, there is a deep groove called the hydroecium, into which the contractile stem with its cormidia can be retracted. In the jelly of the upper most nectocalyx is a space lined by large vacuolated cells, and sometimes containing an oil drop. This is a dilatation of the upper end of the central canal of the stem and is called the stomatocyst. The buds of the cormidia are always formed at the upper end of the stem, so that the oldest cormidium is the lowest. Cormidia are separated by equal sized internodes, thus called endoxiform (Fig. 10a and 10b). Each cormidium consists of two medusoid individuals-the one of these is a sterile and the other is a fertile medusoid form. The sterile medusoid consists of a bract or hydrophyllum, a siphon (trumpet shaped polyp), and a tentacle, while the fertile one is a gonophore. The siphon is the manubrium of the sterile medusoid, which is displaced from its umbrella and has a trumpet-shaped mouth at its free end. The tentacle is the marginal tentacle of the medusoid which has shifted on to the base of the manubrium. The tentacle is tubular and is beset with a series of lateral tubular tentilla. The gonophore has a 4-radiate canal system and a velum but Page 59 of 115 without tentacles and mouth. manubrium. The sex cells originate from the ectoderm of its c. Helistemma: It shows bilateral symmetry in contrast to other coelenterates which show radial symmetry. It is a colonial animal showing a higher degree of polymorphism. It has a long stem bearing a single float (pneumatophore) without a terminal opening of the neumatocyst (an air chamber) on its dorsal side. A number of swimming bells (nectocalyces) are arranged on the both sides of the stem (Fig. 11a and 11b). Each bell has four radial canals, circular canal, a velum and sometimes ocelli. This part of the stem helps in floating of the colony and thus called nectosome. Below the swimming bells, rest of the colony, bearing a number of cormidia arranged in groups, is called siphonosome. Each cormidium consists of one siphon (gastrozooid) with tentacle, mouthless dactylozooid with a tentacle, several bracts (hydrophyllia), two types of gonozooids (male and female) and a palpon like structure which has a terminal opening (anal like) which help in excreting fluid and wastes. The batteries of nematocysts are present on the tentilla attached to the tentacles. Female gonozooid produces only one egg which after fertilization gives rise to a zygote, which develops into planula larva. One end of the planula larva develops a pneumatophore by invagination while the other end forms a polyp which elongates and develops into an elongated colony by asexual budding. 3. Class Anthozoa: Polymorphism is generally absent from anthozoans but one example is given below which shows dimorphism. a. Pennatula (Sea Pen): Its structure is like a quill feather. It has a lower peduncle and a distal rachis (stem). They are found in the warmer coastal waters mainly where bottoms are soft, so that their peduncle can easily be buried in the sea bottoms. There is a horny axial skeleton in the stem and calcareous spicules in the mesogloea. It is anchored in the sand by its peduncle. It is dimorphic and bears two types of zooids (Fig. 12a). From the both sides of the rachis arise lateral branches bearing feeding zooids called autozooids or anthocodia (Fig. 12b). These zooids are joined at their bases in one plane and are attached to the main rachis. They are feeding zooids. On the back side of the rachis are present, small polyps called siphonozooids which have no tentacles and helps in the circulation of water in the colony. From the account given above it is clear that coelenterates exhibit polymorphism and that polymorphism has evolved mainly in the colonial animals and that too in the hydrozoans except few anthozoans. The importance of polymorphism is reflected to have division of labor among different forms and help in the survival and dispersal of the colonial animals. In addition it helps in avoiding overcrowding of species at a particular area. Page 60 of 115 Twig Pedal disc a. Tentacles Mouth Testis Body Bud Ovaries b. Fig. 1a. Hydra hanging down from the twig. 1b. Hydra. Page 61 of 115 Blastostyle Medusa budding from blastostyle Polyp Fig. 2. An Obelia colony showing trimorphism (presence of three types of zooids). Page 62 of 115 Hydranth/ Polyp a Contracted hydranth Medusa buds Exumbrellar surface Oral tentacle Fig. 3a. Bougainvillea colony (a part of the colony). Sense organ Radial canals Marginal tentacles . b. Single medusa of Bougainvillea Page 63 of 115 Gastrozooid Oral tentacles Aboral tentacles Stomach region Gonophores Stalk Fig. 4. Tubularia hydranth. Page 64 of 115 Nematocysts Gastrozooid Tentaculozooid Nematocysts Spiral dactylozooid Young dactylozooid Female gonozooid Male gonozooid Substratum Dactylozooid Fig. 5. Hydractinia showing different types of zooids. Page 65 of 115 Sail or crest Pneumatophore with Concentric air chambers Edge Gastrovascular cavity Tracheal canal Gonozooids with medusae Dactylozooids Mouth Gastrozooid Fig. 6: Diagrammatic section of a Vellela showing different types of zooids. 7a. 7b. Page 66 of 115 Pneumatophore Gonozooids Dactylozooids Dactylozooids Gastrozooid 7c. Chambered Pneumatophore Gastrozooid Medusa Gonozooid Dactylozooids Fig. 7a. Porpita, dorsal view, b. subumbrellar view. c. A section of Porpita. Page 67 of 115 Pneumatophore Ectoderm Nectocalyces Endoderm Hydrophyllium Enteron Gonophore Tentacle Dactylozooid Tentilla Tentacle of dactylozooid Gastrozooid Fig. 8a. A generalized diagram of a siphonophore. Page 68 of 115 Battery of nematocysts Gastrozooid Dactylozooid Tentacle Basal tentacle Nematocyst knobs Tentilla Gonophore Gonopalpon Gonozooids Testis Ovary Male gonophore Female gonophore Fig. 8b: Modifications of polyp and medusa. Page 69 of 115 Sail Gas chamber (Nematocyst) Gastrodermal cavity Short dactylozooid Cormidia Gonozooids Gastrozooid Long dactylozooid Mouth A single cormidium Fig. 9: Part of a Physalia showing the structure of a cormidium. Page 70 of 115 Cavity of swimming bell Swimming bells (Nectophores) Coenosarc Group of zooids (Cormidium) Bract Medusa Gastrozooid Tentacles 10 a. 10 b. Fig.10a: Diagrammatic view of Diphyes colony. Fig. 10b. Enlarged view of a Cormidium (group of zooids). Page 71 of 115 Pneumatophore Swimming bells or Nectocalyces Nectosome Tentacle Coenosarc Tentilla Siphonosome Gastrozooid Gonozooids Hydrophyllum (Bract) Dactylozooid Unbranched tentacle a b (a cormidium) Fig. 11a: A colony of Helistemma. 11b. A cormidium. Page 72 of 115 Anthocodia / Autozooids Pinnulae bearing anthocodia Rachis Anthocodia Pinnulae Peduncle Rachis Siphonozooids 12a 12b Fig. 12a. Pennatula colony. 12b. A part of the Pennatula showing two types of zooids. Page 73 of 115 VI Mesenteries 1. Introduction: Mesenteries are present only in class anthozoa. All anthozoans are polypoid animals and live in marine water. Mesenteries are septa which grow from the gastrodermis of the column body wall and join the pharynx in the upper half of the body. They thus divide the coelenteron into many chambers up to the pharyngeal region. Before discussing the variations of mesenteries in different animals, it is of great importance to study the basic structure and mesenteries of a typical anthozoan- the Metridium. 2. Structure of Metridium (Sea Anemone) explaining Mesenteries: The structure of Sea Anemone (Metridium) is represented here as it is a typical example of the class anthozoa. Longitudinal section of sea anemone (Fig. 1) shows the positioning of mesenteries and other associated structures present within the body. It has a polyp like structure which can be divided into three regions, oral end, column and basal disc. Oral end bears a mouth surrounded by a circlet of tentacles. Tentacles are hollow and the gastrovascular cavity is extended into them. Mouth is a slit like opening guarded by circular muscles. It leads into a pharynx which is lined from inside by an ectoderm, mesogloea and gastroderm. There are two ciliated grooves called siphonoglyphs which allow circulation of water within gastrovascular cavity even if mouth is closed. Pharynx opens into the gastrovascular cavity or the coelenteron. Column represents the main body of the animal. Basal disc is used for the attachment of the animal to the substratum. The gastrovascular cavity is divided by paired mesenteries which according to their size and position are differentiated into primary, secondary and tertiary mesenteries. A. Primary mesenteries or complete mesenteries: These are those paired mesenteries that are connected to the body wall on one side and to the stomodaeum (pharynx/siphonoglyph) on the other side. These completely divide the gastrovascular cavity into separate chambers. This is shown in a transverse section of Metridium through stomodaeum (Fig. 2a). Below stomodaeum, primary mesenteries hang in the coelenteron (Fig. 2b). Each mesentery is lined on both sides by the gastrodermis with a thin sheet of mesogloea in between as these are the extensions of the gastrodermis towards the pharynx. The part of the gastrovascular cavity or coelenteron enclosed between a pair of mesentery is called endocoel while within the adjacent pairs is called exocoel. The mesenteries of each pair bear longitudinally arranged parietal and retractor muscles. There is a specific arrangement of retractor muscles on the mesenteries. Retractor muscles are present on one side of each mesentery and are visible as bulging present on the mesenteries. These retractors of adjacent mesenteries either face towards each other (facing endocoel thus called endocoelic) or do not face each other (facing exocoel thus exocelic). Generally, mesenteries attached to the siphonoglyph have retractors facing away from each other and thus are called directives. B. Secondary mesenteries or incomplete mesenteries: These are those mesenteries that are connected to the body wall and do not reach to the stomodaeum (Fig. 2b), they hang freely in the coelenteron or gastrovascular cavity. They may be secondary or Page 74 of 115 tertiary mesenteries. Six pairs of secondary mesenteries are present between the primary mesenteries and do not reach the gullet, thus hang halfway from the body wall into the gastrovascular cavity. While, 12 pairs of tertiary mesenteries are much smaller and are present in exocoels present between the primary and secondary mesenteries. Secondary and tertiary mesenteries are formed later than the primary mesenteries during development. So in Metridium there are total 24 pairs of mesenteries including 6 primaries, 6 secondaries and 12 tertiaries. The free ends of incomplete mesenteries are trilobed in cross section (Fig. 3a), having two lateral lobes and one central lobe (Fig. 3b). Lateral lobes are lined by ciliated endothelial lining and called ciliated lobes while central lobe bears lot of nematocysts and glandular cells producing enzymes and mucous, thus called cnido-glandular lobe. Lower down in the coelenteron, ciliated lobes disappear and only cnidoglandular lobe remains and forms the absorptive part of the gastrodermis and bears batteries of nematocysts to paralyze the prey (Fig. 3c). In some sea anemones, the ends of the mesenteries are drawn into thread like structures called acontia, containing mucus cells and cnidoblasts. These acontia can come out of the small openings called cinclides present in the column body wall (Fig. 1). Acontia provides protection and help in the digestion of food as they bear number of nematocysts (stinging cells) and enzymatic cells within in their gastrodermal lining. Near the terminal ends of the incomplete mesenteries, gonads are present which bear sex cells for the reproduction. They are dioecious and sperms and ova are released in the water through mouth and fertilization takes place in the water. Zygote develops into a free swimming ciliated planula larva which attaches to the substratum and grows into an adult sea anemone. There is an extensive asexual reproduction by budding, or by splitting of the polyp into parts, where each part develops into a new polyp or by pedal laceration in which animal moves further and a part of the pedal disc is left behind. Detached pedal disc regenerates into another sea anemone. The number and arrangement of mesenteries is different in different animals of class anthozoa which is obvious by looking at the following examples: 3. Octocorallians: are polypoid animals which are colonial, bear eight pinnate feather like tentacles, 8 complete mesenteries, have one siphonoglyph and supporting skeleton is endoskeleton which is soft or horny, perforated by gastrodermal tubes that are continuous with the gastrovascular cavity of the polyp and are supported by calcareous spicules. 4. Hexacorallians: are polypoid animals which are colonial bear either six pairs of mesenteries or in the multiples of six. The number of mesenteries is generally equal to the number of tentacles. In hexacorallians, the six largest tentacles correspond to the six primary mesenteries and are developed first during the course of development. Later on more tentacles are added which corresponds to the secondary mesenteries and so on. 5. Examples of anthozoans showing different arrangement of mesenteries: a. Alcyonium: It has all the diagnostic features of octocorallians. The polyps (anthocodia) have 8 pinnate tentacles and 8 mesenteries (Fig. 4). The siphonoglyph is single and the side on which it is present is called ventral (no homology to the ventral surface of higher animals). A pair of mesenteries attached to the siphonoglyph have longitudinal muscles facing towards the endocoel ( also called sulcul mesenteries) while a pair of mesenteries present on the opposite side (i.e. dorsal side in respect to siphonoglyph) bear muscles facing away from each other(towards exocoel and are called Page 75 of 115 asulcal mesenteries). The dorsal mesenteries are often longer than the others, and are developed earlier in the bud but later in the egg. Rests of the four mesenteries are unpaired and bears muscles which are facing down (Fig. 5a and b). Below pharynx, six of the mesenteries hang freely in the coelenteron and bear unilobed ends while asulcal pair of mesenteries bear cilia and help in circulation of water within a polyp and also throughout the colony for the renewal of food and oxygen. Alcyonium has one of the simplest body forms and simplest type of arrangement of mesenteries found in the anthozoans. Similar type of arrangement of mesenteries is also present in the polyps of Tubipora. Many of the hexacorallians like sea anemone and stony corals pass through a larval stage (Edwardsia stage, Fig. 6) in their development in which they resemble a young alcyonarian polyp in having 8 mesenteries. b. Edwardsia: It has also total 8 mesenteries like Alcyonium. The Tentacles are more in number as compared to the number of mesenteries. There are two siphonoglyphs, so two pairs of mesenteries are directives and are attached to both the siphonoglyphs. Muscles present on the both pairs of directives do not face each other in contrast to Alcyonium where muscles face each other on one pair of directives (Fig. 6). Rests of the four mesenteries are unpaired and their muscles face downwards like Alcyonium. Edwardsia is an important animal as it resembles the octocorallian structure (have 8 mesenteries) and helps in tracing the origin of hexacorallians from octocorallians. c. Gonactinia: The arrangement of mesenteries of this anthozoan resembles the arrangement seen in Edwardsia. There is an addition of few mesenteries in this animal e.g. 4 new mesenteries (microsepta) facing 4 already existing unpaired mesenteries (seen in Edwardsia) have started growing (Fig. 7). Another difference is that two pairs of secondary mesenteries (incomplete) have also grown up. With the result this animal has total 8 pairs of mesenteries, out of which two pairs are secondary mesenteries, two pairs are directives and 4 pairs are under completion. d. Halcampoides: It is a primitive sea anemone having two siphonoglyphs. There are only 6 pairs (12) of primary mesenteries out of which two pairs are directives bearing exocoelic muscles (Fig. 8) and four pairs are bearing endocoelic muscles. These first formed paired and complete mesenteries are called protocnemes. Secondary and tertiary mesenteries were not developed at all in this animal. . e. Halcampa: It has six pairs of primary mesenteries like Halcampoides. In addition it has 6 pairs of small underdeveloped 6 pairs of secondary mesenteries called microsepta in the region of stomodaeum (Fig. 9). So, in this animal rudiments of secondary mesenteries are reported which are completely formed and are reported in the Adamsia. f. Adamsia: It has two siphonoglyphs. There are total six pairs of primary mesenteries out of which two pairs are called directives and the longitudinal muscles present on them do not face each other while longitudinal muscles present on rest of the four pairs of mesenteries face each other. 6 pairs of secondary mesenteries are fully developed each having endocoelic retractor muscles (Fig. 10). Even, 6 pairs of tertiary mesenteries are also reported in this animal. Now, if we compare the mesenteries of Adamsia with the Metridium, both of them have similar type of arrangement of mesenteries (Fig. 2a). g. Haloclava: It is decamerous as it has total 10 pairs of primary/complete mesenteries some of which are formed by the extensions of the micro-septa joining the stomodaeum Page 76 of 115 (Fig. 11). Both pairs of directives have muscles towards the exocoel while rest of the mesenteries has retractor muscles facing towards the endocoel. h. Zoanthus: The arrangement of mesenteries is quite different from others and bears either 6 or in the multiples of six. There are total 12 mesenteries out of which 6 are complete mesenteries and 6 are incomplete mesenteries. Out of 6 complete mesenteries, two directives are complete and two directives are incomplete (Fig. 12). Rest of the 4 pairs of mesenteries consists of one complete (macro-septa) and other incomplete (microsepta) mesenteries with endocoelic retractor muscles in each pair. Such an arrangement of mesenteries is called brachycnemous. i. Epizoanthus: It has total six pairs of mesenteries out of which, 3 pairs are complete mesenteries (macrosepta) i.e. the ventral directives, 4th and 5th mesenteries on each side starting from the dorsal directives (Fig. 13), while 3 pairs are incomplete mesenteries(1st, 2nd and 3rd pair). j. Cerianthus (a burrowing anemone): Sand living, elongate, solitary polyps. They secrete a tube of the discharged threads of ptychocysts (special cnidoblasts). Cerianthus has a single siphonoglyph and many complete, unpaired and coupled mesenteries. Their number is not fixed, as it grows, more and more mesenteries are added towards the dorsal growth zone (dorsal interseptal space) and not between already existing couples (Fig. 14). The two ventral directives attached to siphonoglyph are small. The mesenteries present on either side of the directives are larger and reaches to the aboral end. The mesenteries decrease in size towards the dorsal region where new mesenteries are added. The retractor muscles are absent in all the mesenteries. k. Antipathes (Black coral): It has six tentacles. Mesenteries may be 4, 6 or 10. There are 10 complete mesenteries having rudimentary retractor muscles (Fig. 15a). Another section shows presence of four directive mesenteries and other two lateral mesenteries which bear gonads for the sexual reproduction. In addition there are 4 secondary mesenteries (Fig. 15b). Mesenteries are without mesenteric muscles. Skeleton is branched, made up of scleroprotein (antipathin), consists of black ectodermal chitinous axis covered with thorns. It is found in deep waters. l. Peachia: It burrows in the sand when adult but its larval form live as parasites or commensal in the radial canals of scyphomedusae (Fig. 16). It has 6 pairs of complete mesenteries and 4 pairs of incomplete mesenteries. m. Metridium: As we have seen in fig.2a where it has 6 pairs of complete mesenteries, 6 pairs of secondary mesenteries and 12 pairs of mesenteries which are developed sequentially giving rise to total 24 or 12 pairs of mesenteries. 6. Formation of mesenteries: Formation of mesenteries have been reported in some larval forms of hexacorallians i.e. Actinaria. Mesenteries are not added at random but they are formed in a specific manner. Some of the stages have a particular arrangement of mesenteries which are found to be present in some adult anthozoans. This is evident from the arrangement of mesenteries discussed earlier in various anthozoans.. The formation of mesenteries is explained below in brief (Fig 17a to 17f). Step 1: Initially considering there is no mesentery within the coelenteron. First of all two mesenteries (1st pair mesenteries) are formed at right angles to the pharynx and divide the gastrovascular cavity into two chambers; one is dorsal and bigger (LC) while Page 77 of 115 the other is ventral and smaller chambers (SC) giving rise to bilateral symmetry (Fig. 17a). Step 2: 2nd pair mesenteries started appearing in the bigger chamber (LC) Fig. 17b. Step 3: 3rd pair of mesenteries begin to appear within the smaller chamber (SC) shown in Fig. 17c. Step 4: 4th pair of mesenteries are formed within the chamber enclosed by 2nd pair (LC) mesenteries, just opposite to the 3rd pair of mesenteries (Fig. 17d). This type of arrangement is found to be existed in Edwardsia (Fig. 6). Step 5: Two paired mesenteries grew between the 1st, 2nd and 1st, 3rd. Newly formed mesenteries remain incomplete and their longitudinal muscles face already existing 2nd and 1st pair mesenteries (Fig. 17e). In this way, 8 complete and 4 incomplete mesenteries are formed which are also found to be present within Gonactinia (Fig. 7) and with little modification in Zoanthus (total 12 complete or incomplete mesenteries (Fig. 12). Step 6: A pair of small mesenteries destined to be incomplete mesenteries start appearing within exocoels (Fig. 17f). In Peachia only four pairs of incomplete mesenteries are found to be present. Later on, more and more mesenteries are added which may be secondary or tertiary mesenteries as are already discussed. In the examples discussed above, 3rd and 4th pair of mesenteries became the two pairs of directives. It is very clear that secondary and tertiary mesenteries are developed only in the exocoels. The arrangement of longitudinal muscles is also very specific as the muscles present on the directives faced exocoelic chambers in majority of animals, in contrast to other mesenteries where muscles faced endocoelic. The gonads are developed in the mesenteries and the sex cells are lodged in the endoderm. Once the sex cells are mature, they are released in to the coelenteron. 7. Significance and function of mesenteries: The main function of the mesenteries is to increase the surface area. The free terminal ends of the mesenteries are very long and bear more number of enzymatic cells, mucous cells and cnidoblasts. Their increased surface area not only helps in killing the prey but also helps in extracellular and intracellular digestion. Retractor muscles present on the mesenteries also help in retraction and invagination of the oral disc and tentacles. If a sea anemone is over stimulated, it retracts its body fully and acontia are protruded out of the mouth or through cinclides in the column. Here, the nematocysts present on the acontia are used for the defense purpose and for killing the prey. Besides they also help in excretion and production of gonads for the sexual reproduction. The tentacles are also developed in a similar order to that of the developing mesenteries. First pair of tentacles appears with the appearance of the dorsal bigger chamber (LC) and is longer than rest of the tentacles which are added later on as the embryo grows into an adult. From the edges of the mouth arise siphonoglyphs which help in maintaining a continuous supply of water inside the gastrovascular cavity even if the mouth is closed. The number, type and arrangement of mesenteries determine the classification pattern of most of the anthozoans. In addition they also help in tracing the evolution of many anthozoans. Radial symmetry in anthozoans is also lost due to the presence of siphonoglyph and mesenteries. With the result, bilateral symmetry (when one Page 78 of 115 siphonoglyph is present), or biradial symmetry (when two siphonoglyphs are present) is attained. Bilateral symmetry or biradial symmetry is attained during embryonic development and persists into the adult which is another sign of gradual evolution of higher animals having bilateral or biradial symmetry. Page 79 of 115 Mouth Oral disc Tentacles Fossa Siphonoglyph Oral ostium Capitulum Parapet Sphincter Longitudinal retractor muscles Pharynx Marginal ostium Complete mesentery Mesenteric filament Transverse muscle Mesenterial filaments Gonads Gastrovascular cavity Acontium Limbus Cinclide Parietal muscle Epidermis Acontia Basal disc Fig. 1: Longitudinal section of Metridium showing mesenteries and other structures. Page 80 of 115 Exocoel Endocoel Paired secondary mesenteries Epidermis A pair of Primary mesenteries Gastrodermis Directive couple A pair of Tertiary mesenteries Pharynx Siphonoglyph Gonads Retractor muscles . Gastrodermis 2a Epidermis Exocoel Mesogloea Endocoel Mesenteric filaments Coelenteron 2b Fig.2a. Diagrammatic transverse section of Metridium through pharynx showing mesenteries. 2b. Diagrammatic transverse section of Metridium below pharynx showing hanging mesenteries. Page 81 of 115 Epidermis Mesogloea Gastrodermis Parietal muscles Retractor muscles Gonads Cnidoglandular lobe 3a. Fig. 3b Fig. 3c Mesogloea Ciliated cells Ciliated tract Gland cells A core of mesogloea Gland cells Nematocyst Nematocyst Cnidoglandular lobe Cnidoglandular lobe Fig.3a: T.S. of the body wall of sea anemone showing trilobed cnidoglandular lobe of a single mesentery. 3b. Magnified view of T. S. of a trilobed end of mesentery (upper part). 3c. Magnified view of T.S. of unilobed end of mesentery (lower part). Page 82 of 115 Pinnules Tentacles Mouth Opening of pinnule Siphonoglyph Mesenteries Coelenteron Solenia Soft skeleton due to gastrodermal tubes Gastrodermal tubes Axial skeleton . Fig. 4: Structure of an octocorallian polyp Page 83 of 115 Asulcal mesenteries or directives Asulcal mesenteries having ciliated ends Retractor muscles facing outwards Endocoel Coelenteron Exocoel Epidermis Mesogloea Pharynx Siphonoglyph Stomodaeum Sulcal mesenteries or directives With retractor muscles facing inwards 5a Gastrodermis Unilobed end of mesentery 5b Asulcal mesenteries or directives, Retractor muscles facing outwards Exocoel Secondary mesenteries Endocoel Primary mesenteries Microsepta Siphonoglyphs Directives retractor muscles facing outwards 6 7 Fig. 5a: T.S. Alcyonium through pharynx showing arrangement of mesenteries. 5b: T.S. Alcyonium below pharynx. Fig.6. Cross section through Edwardsia. Fig. 7. Cross section through Gonactinia. Page 84 of 115 A pair of Primary mesenteries Exocoelic retractor muscles Exocoel Endocoel Siphonoglyphs Endocoelic retractor muscles Fig. 8. Cross-section through pharynx of Halcampoides showing 6 pairs of complete mesenteries. Siphonoglyphs Secondary mesenteries Endocoel Exocoel A pair of Primary mesenteries Fig. 9. Cross section through pharynx of Halcampa showing 6 pairs of complete (primary mesenteries while 6 pairs of secondary mesenteries are beginning to appear. Page 85 of 115 Exocoel A pair of primary mesenteries Secondary mesenteries Endocoels Tertiary mesenteries Fig. 10. Cross section through pharynx of Adamsia. Retractor muscles Siphonoglyphs Endocoel Exocoels Fig. 11. Cross section through pharynx of Haloclava showing 10 pairs of primary mesenteries. Page 86 of 115 Incomplete Dorsal directives Complete mesenteries Incomplete mesenteries Complete ventral directives Fig. 12. Section of Zoanthus through pharynx showing 6 complete and 6 incomplete mesenteries. Incomplete Dorsal directives 1 1 2 2 Incomplete mesenteries 3 3 4 4 5 5 6 6 Complete ventral directives Fig. 13. Section of Epizoanthus through pharynx. Page 87 of 115 Complete mesenteries Ventral directives Siphonoglyph All Complete mesenteries Ventral exocoel, new mesenteries are added here. Youngest mesentery Fig. 14. Section through pharynx of Cerianthus. Siphonoglyph Incomplete mesentery Siphonoglyphs Poorly developed retractor muscles 15b 15a Complete mesenteries Fig. 15a. Antipathes section with 10 complete mesenteries. Fig.15b. Antipathes section showing 6 complete mesenteries and 4 incomplete mesenteries. Page 88 of 115 Secondary mesenteries Siphonoglyph Primary mesenteries Fig. 16. Section of Peachia through pharynx. Page 89 of 115 a b 2 LC 2 LC Dorsal Larger chamber (LC) LC LC 1 1 SC 1 1 Ventral Smaller SC chamber (SC) 4 c d 2 2 2 2 LC LC 1 LC LC 1 SC 1 BC LC SC 1 SC Present in Edwardsia 3 3 e f 4 4 2 2 5 5 Incomplete mesenteries 1 1 6 Present in Gonactinia and Zoanthus 6 3 6 6 Present in Peachia 3 Figs. 17a to 17f. Formation of mesenteries. They are numbered according to the order of their development. Page 90 of 115 VII Corals and coral reefs 1. Introduction: Corals are hard structures that are made up of calcium carbonate and are deposited into big stony formations by the anthozoans which are exclusively marine water living. All anthozoans are polypoid animals without any medusoid form at any stage of the development. The polyps produce sex cells, fertilization gives rise to formation of zygote developing into planula larva, which directly form new polyp. These anthozoans may remain singly or form big colonies and majority of them secrete hard calcareous structures around them starting from their basal disc. Anthozoans bear cilia on different regions of the body e.g. the cilia present around the mouth beat towards outer side and thus clean the oral end. Colonial and fixed polyps show a great degree of development by secreting enormous structures called coral reefs which play an important role in maintaining aquatic ecosystem. Coral reefs are submarine ecosystems which can be compared with the rain forests as they also host diverse flora and fauna. The reef building corals require warm shallow waters i.e. above 200C. They are therefore limited to the Indo-Pacific, Central Western Pacific, and the Caribbean regions north to Bermuda, while other corals live at moderate depths throughout the world. In addition to different types of the coral forming anthozoans, there are variety of other organisms belonging to different groups which also contribute significantly in the formation of big stony structures over the years called coral reefs. Different anthozoans and hydrozoans which contribute to the formation of coral reefs are briefly discussed below. 2. Class: Anthozoa: A majority of the coral forming animals belongs to class anthozoa and structurally resembles sea anemones except that they do not have siphonoglyph. A coral polyp resembles the basic structure of the sea anemone. A. Subclass: Octocorallia or Alcyonacea a. Structure of Octocorallian coral: Octocorallians are mostly represented by the Alcyonarians (soft corals) and gorgonians in many coral reefs. Besides this, the red coral (Tubipora) and Blue coral (Heliopora) having calcareous, massive skeleton are also coral forming octocorallians and contribute in the formation of coral reefs. Each Octocorallian bears 8 pinnate tentacles (tentacles have small side branches like a feather), and have 8 complete mesenteries on either side of the base of the tentacle. One siphonoglyph is present. Polyps are small and are joined to each other by the skeleton called coenenchyma which is secreted by the ameobocytes present within the mesogloea (Fig. 1a). Therefore, the skeleton of the octocorallians is internal in contrast to external skeleton present in the hexacorallia. The skeleton may consist of spicules of different sizes and shapes (Fig. 1b) or horny strands made up of calcium carbonate. Coenenchyma consists of thick mesogloea which is perforated by the gastrodermal tubes which are in continuity with the gastrovascular cavities of the polyp bodies. Besides these gastrodermal tubes there is a network of small ciliated interconnecting tubes called solenia which help in maintaining a current of water inside the colony. It is covered on the outer side by the epidermis which is continuous with the Page 91 of 115 epidermis of the polyp. Lower part of the polyp is present within the coenenchyma so that only upper part pf polyp is protruded out of it. They are commonly found in warm waters of Indo-Pacific Ocean. A few examples are given below to have an idea about different types of corals. i. Alcyonium (Dead man’s finger or soft coral): It is the soft coral and is one of the simplest body forms of the anthozoans. It bears only feeding polyps. The polyps are embedded in a fleshy mass called coenchyma from which only delicate oral ends of the polyps (anthocodia) protrude and can be withdrawn within the coenenchyma at the time of danger (Fig. 2). Polyps are located only on the free end, whereas the proximal part is sessile (fixed) is devoid of polyps. Skeleton is made up of separate calcareous spicules which form a mass of calcium carbonate after their death. ii. Heliopora (blue coral): It is bright blue in color and is found living along with the true corals on the coral reefs of the Indo-Pacific. They live in symbiotic relationship with zooxanthellae so are zooxanthellate. It is the only octocorallian in which the secreted spicules are not separate but form a massive calcareous skeleton perforated by numerous closely set cylindrical cavities (diverticula’s) closed below (Fig. 3) which increase the surface area. Skeleton is lobed and bears polyps which can withdraw themselves within the pores. Live polyps are always present on the surface of the skeleton as colony grows. The skeleton formed by one polyp is called a corallite while many polyps form a big coral structure which is known as corallum. The corallum not only have pores for the polyps but also has tubular cavities of intermediate size which sometimes are occupied by a chaetopod worm, (Leucodore genus). When worms are protruded out from their holes in search of food, polyps get retracted within their pores. iii. Tubipora (Organ pipe coral): Tubipora as its name suggests has long, horny, tubes joined by horizontal platform in which polyps are present. The base of the polyp grows out and form horizontal platform containing a tube inside called solenia from where new polyps arise (Fig. 4). Polyps lie parallel to each other and their internal skeleton is made up of fused spicules produced by the ameobocytes present within mesogloea. With the result large colonies of the coral are produced. Polyps are present in the tubes partly projecting above. Polyps are green, while skeleton or coral produced is dark red due to the deposition of iron salts and calcium carbonate. iv. Corallium (Red coral): It is a dimorphic colony with upright branches having a rigid axis supported by the delicate tissue of coenosarc from which the short polyps arise (Fig. 5). Polyps are of two types i.e. anthocodia and siphonozooids, which arise in perpendicular to the central axis lined by gastrodermal tubes (could be polyp coelenterons). Anthocodia are feeding zooids while siphonozooids bear gonads and help in sexual reproduction. Spicules are compacted with the calcareous cement like substance and produce hard axial skeleton. It is one of the precious red coral and found in the Mediterranean Sea and the Sea of Japan. It is used for making jewelry. v. Gorgonia (Sea fan): They form a large interwoven tree like structure with branches arising from the common basal plate. Branches arise only in one plane. They bear numerous slender polyps called anthocodia with 8 pinnate tentacles while siphonozooids are absent. All branches are interconnected by small cross connections to form a mesh like structure. Within the mesogloea there is a network of branching tubes called solenia which are extensions of the coelenteron (Fig. 6). The skeleton is made up of horny protein called gorgonin around which calcareous skeleton is secreted. They are expanded during the day than during the night. As gorgonians grow bigger, many small animals Page 92 of 115 and symbiotic algae grow and crawl on the surface and get shelter and food. The color of the gorgonians depends on the colour of the calcareous spicules and symbiotic zooxanthellae growing within the colony. B. Subclass: Hexacorallia (Zoantharia): a. Structure and formation of a Hexacorallian coral: Hexacorallians include stony corals which are also called true corals; therefore they are very important and contribute a lot in the formation of coral reefs and coral islands. These are cosmopolitan in their distribution in marine water and secrete compact calcareous exoskeleton. They have mesenteries and tentacles either 6 in number or in the multiples of six. Tentacles are simple. They may be solitary or colonial. Coral polyps are similar in structure to sea anemones but do not have siphonoglyph. The skeleton is secreted externally by the epidermis of lower half of the column as well as by the basal disc of the coral polyps (Fig. 7a). The cells secreting the hard skeleton are called calicoblasts which is composed of calcium carbonate. So, each polyp is fixed in a cup like structure called calice. Calice is that part of the coral which is in direct contact with the basal ectoderm of the polyp. The shape of the calice may be cups like, conical or saucer shaped. The flat base of the cup secreted by the polyp beneath it is called basal plate and surrounds the polyp base, while the side walls of the cup are called the theca. From the basal plate, sclerosepta arise vertically between the mesenteries (Fig. 7b). Sclerosepta are made up of rod like skeletal elements called trebaculae. As sclerosepta grows up, the lower part of the polyp is invaginated forming the blunt end ridges towards the gastrodermal cavity. These ridges alternate with the grooves enclosing the mesenteries. Sclerosepta help in fixing the polyp to the basal plate and also help protect them from predation. Mesenteries are also the site of digestion, excretion and gonad development. They have double layers of gastroderm enclosing a thin sheet of mesogloea as discussed earlier in the previous chapter of mesenteries. . The mesenteries are arranged in pairs. The space between each pair is called endocoel while the space present between adjacent pair of mesenteries is called exocoel. The lower epidermis secretes the skeleton which grows in the form of sclerosepta in between the endocoel in the beginning but more can be added even in the exocoel as the polyp grows. The inner ends of the sclerosepta are fused to form an irregular central skeletal mass called columella. In some corals, there is a special type of growth producing narrow upright pillars along the inner ends of the sclerosepta. These are called palli and are formed in a circle around columella. The skeleton secreted by a single polyp is called a corallite and is called a solitary coral (Fig. 8a and b). Solitary corals are constricted at regular intervals. In colonial corals, a single polyp grows asexually by producing buds laterally, or by vertical fission into two or more daughter polyps within the calice, and skeleton is secreted by each polyp. Thus polyps producing colonial corals are joined with each other laterally near the bases. These lateral connections have upper and lower layers of ectoderm and gastroderm with the extensions of the gastrovascular canals. In colonial corals, thousands of corallites are secreted by thousands of polyps growing together by budding or splitting. As all the polyps are joined together by horizontal partitions, the resulting combined hard skeleton produced by a colony together is called corallum. Live polyps are always present over and above the dead calcareous corallum secreted at the base by them. A few examples are given below: i. Fungia (Mushroom coral): It is a solitary discoid coral. Planula larva develops into a single stalked animal resembling a cup like coral. Adult animal has a single polyp with Page 93 of 115 many tentacles. Once it grows, it breaks itself from the stalk and free coral grows and is moved by the water currents. It can perform certain movements because of the presence of cilia and by inflating coelenteron with the water. The theca (cup) is confined to the lower surface. Sclerosepta as usual grow within the base of the polyp forming ridges and grooves. As sclerosepta grow, they are fused in the center producing columella. Many thin synapticulae grow horizontally in between each sclerosepta connecting adjacent septa and close the interseptal spaces. It secretes coral which is convex on the upper side and concave on the lower surface (Fig. 8a). ii. Madrepora (Horn coral): It is a colonial coral with many perforations present within the coral. Colony is branched like the horns with small polyps within cup like structures separated from each other by perforated coenosteum (The lower surface of the coenosarc secretes the skeleton between adjacent corallites, which is known as coenosteum). Terminal polyps bear 6 tentacles while lateral polyps bear 12 tentacles. The polyps are connected by the coenenchymal canals passing through the loosely secreted theca, (Fig.9) and as the corallites have many pores, are called perforate corals. iii. Astraea (Star coral) : The colony consists of numerous polygonal cups or theca. Each cup encloses a polyp and is very close to each other and shares their walls (Fig. 10). Polyps are connected by coenenchyma only over the upper edge of the theca. Coenenchyma is formed by calcification of the coenosarc and gives rise to individual corallite which lie near by. Skeleton produced by them is very hard, stony and therefore produce massive corals. There are no pores or perforations in the coral colony and thus called an imperforate coral. iv. Meandrina (Brain coral): Brain corals form extensive masses of hard structures. The surface of coral is marked by long, wavy ridges and grooves almost parallel to each other and resembles the grooves and ridges present on the surface of the human brain. Here many polyps are fused together so that they have a fused calice at the base, sharing a common fringe of tentacles, rows of mesenteries and sclerosepta. But the mouths of these joined polyps remain separate. v. Astrangia (white coral): It inhabits the waters of north Atlantic coasts. These corals secrete calcareous cups into which the polyps live and delicate polyps can withdraw inside the cups at the time of danger. vi. Antipathes (Black coral): It is a black coral. It is tree like with upright, plant like colonies (Fig. 11a and 11b). Polyps bear six tentacles, out of which two opposite ones are longer than others. Stem and branches have thorny appearance and have flexible axial skeleton within the polyps. Skeleton is composed of a black, non-collagenous horny material called antipathin. Polyps may have 6, 10 or 12 mesenteries and two siphonoglyphs, but without muscles. Because of the absence of muscles, these polyps cannot contract immediately like other anthozoans. 3. Class: Hydrozoa: Some animals of the class hydrozoa like Millepora also contribute towards the formation of coral reefs. i. Millepora (Fire coral or sting coral): It is a colonial animal and bears gastrozooids, dactylozooids and ampullae (which produce medusae). The calcareous skeleton secreted by zooids bears number of pores and is called coenosteum. Through these pores zooids protrude out. A gastrozooid is always surrounded by many dactylozooids. The dactylozooids are longer than gastrozooids and bear capitate tentacles bearing nematocyst which are highly irritating to humans. Some species have special rounded depressions called ampullae in which simple medusae are produced for the sexual reproduction. They Page 94 of 115 always live in symbiotic relationship with zooxanthellae and thus corals produced are brown in colour (Fig. 12a and 12b). They secrete enormous skeleton and thus make significant contribution in the development of coral reefs. 4. Coral reefs: Coral colonies grow in size either by budding of the new polyps or by vertical splitting of the body of the polyp. All the polyps are joined together at their bases and the skeleton is hardened by the growth of inter-trebaculae horizontal partitions. As a result live polyps move towards the top regions of the colony and are found growing there. The skeleton secreted by them also grows as each polyp help in the formation of corals. Scleractinian corals play a major role in the formation of corals reefs. Scleractinian are the stony corals which secrete enormous amount of hard external skeleton made up of calcium carbonate. As we have already discussed different types of coral forming animals, it is very clear that some are solitary corals and majority of them are colonial corals. Different types of colonial corals growing together along with other associated fauna form large heaps of calcareous mounds of lime stone called coral reefs, the upper surface of which is near the surface of the sea. Such large rigid structures called coral reefs are built up by many generations of coral polyps. The coral reefs provide a natural habitat for diverse organisms like, fishes, mollusks, chaetopods, arthropods and echinoderms which live together in a perfect harmony in a most complicated ecosystem. Dead remains and shells of these animals also contribute to the growing corals reefs and thus indirectly help in the formation of massive coral reefs. It takes millions of years for a full fledged coral reef system to establish itself. Majority of the coral reefs were formed during palaeozoic era. The madreporarian corals (modern corals) replaced them in the coenozoic era and form the majority of the existing reefs and islands present in the Indian and Pacific oceans. A. Development of coral reefs: The development of coral reefs takes place in four stages. First of all a single coral polyp colony grows. Later on this colony is joined by other colonies and this becomes a thicket which may consist of colonies of same species or different species. Thicket, thus created attracts other animals, like crustaceans, mollusks, echinoderms, fishes and barnacles etc., forming a different niche (Fig. 13a). The new ecosystem thus developed will always have wear and tear of the fauna and associated corals due to interaction among different animals. When so many different animals are staying together, some of them may be causing harm or may be benefiting others. They also create lot of debris and excretory wastes. Some animals may be utilizing the wastes, so they further attract more animals for the food and shelter. This leads to further growth of the substratum for coral growth and another stage is developed called coppice which may be several meters across which leads to increase in the substratum for the growth of the colonial animals. As a result there is more debris production thus increasing the number of coral inhabitants, with the result a bank is developed which is known as a coral reef. As we have already discussed that there are many important factors which determine the growth of corals, so obviously there will be a gradation in the type and frequency of the corals at different depths of the sea starting from the sea shore as each species occupies a specific habitat. Different regions of the coral reefs have different species, e.g. in some regions, gorgonians are predominantly present while in other regions scleractinian corals are present. For example the corals present at the inner reef flat, lagoon, outer reef flat, Page 95 of 115 reef edge, and seaward slope will be different as all these different zones of the coral reefs have different environmental conditions. This is called zonation of the corals, which varies from reef to reef depending upon the environmental factors (Fig. 13b). . B. Types and structure of Coral reefs There are three types of coral reefs: a. Fringing reefs b. Barrier reefs c. Atoll a. Fringing reefs: Fringing reefs are developed in shallow waters on or near the shores of the volcanic islands. These are the simplest kind of reefs. They extend from the sea shore towards the sea as a platform ranging from few meters to half a kilometer and then slopes down towards the bottom of the sea (Fig. 14). Fringing reefs consist of several zones that are characterized by their depth, the structure of the reef, and its plant and animal communities. If a section passing through the volcanic island and the fringing reefs is observed, it shows different regions which can be diagrammatically shown (Fig. 15). These regions differ from reef to reef and are not fixed in their occurrence. Reef edge or reef front: The edge of the reef where the coral growth is maximum and it is slowing down towards the sea in a steep slope is called seaward slope. It is belt like and broken here and there by water channels. It is covered by attached plants and animals. Seaward slope: Beyond the reef edge is a steep slope of the coral reef down to the sea bottom. Live corals are present on this seaward slope from 40-100 meters deep depending upon the penetration of light as many animals live in symbiotic relationship with algae. Reef flat: Reef flat can be inner reef flat and outer reef flat. Inner reef flat is the region of the coral reef present between the shore and the reef edge. It contains coral sand, detritus, dead remains of the coral colonies, and other animals. It may be 50 to 100 meters in breadth. It may take a form of a lake through which small boats can pass through. Outer reef flat is another flat which may or may not present between boulder zone and reef edge. The reef flat is generally submerged at high tide but it is exposed at low tides. Boulder zone: When water is flowing through the reef flats; stones, sand, pebbles, coral sand, detritus, dead remains of the coral colonies, and other animals etc get accumulated in an area called boulder zone and may be swept partly and are carried across the reef flat by the surfs created by water currents. Lagoon: It is the channel of water present between sea shore and reef edge of volcanic island and another reef present within the sea water. b. Barrier reefs. A volcanic island with the fringing reefs is surrounded with a big channel of water called lagoon (Fig. 16). Its depth may vary from 20 to 100 meters and even ships can pass through it. Lagoon is further surrounded by reefs called barrier reefs. As their name suggests, they act as a barrier for ships between sea shore and the main sea. Lagoon has fringing reefs towards the volcanic coast and barrier reefs on the other side of it. Sometimes both fringing reefs and barrier reefs may join each other at the bottom of the sea. There is a Great Barrier Reefs of Australia. It is not a single structure but is made up of many strings of separate reefs joined to each other at the bottom and thus forms a Page 96 of 115 very big structure which extends along the north eastern coast of Australia for over 2000 kilometers. It is separated from the main land by a lagoon which is around 15 to 250 kilometers wide and 70 meters deep. During high tide, big ships can sail over it without realizing that reefs are present nearby and may crash. The Great Barrier Reef is the contribution of all different kinds of coral growth over the years. c. Atoll: Atolls are coral reefs which are present within sea water hundreds or thousands of kilometers away from the nearest sea shore. There is no volcanic island present. It is more or less circular or horse shoe shaped reef enclosing a central lagoon which may be 40 or 50 miles across and 20 to 90 meters deep. It may be a complete or broken into many small reef islands separated from each other by water channels (Fig. 17). At some places reef is low so sea water simply covers it and reef is not visible. Sometimes a large atoll is formed by many small islets joined together along a line of reef. Thousands of such atolls are found in the South Pacific. It must be noted that reefs are not continuous rigid structures but they are broken up into many reefs and islands by water channels. Suvadiva is the largest atoll present in Maldives. Its circumference is about 195 kilometers and consists of around 102 separate little islets on its rim. Bikini Atoll has 2.87square miles land area with a lagoon area of 280 square miles. It was inhabited by the people but people moved to different places as it was selected by the United States for testing hydrogen and atomic bombs. Horse shoe shaped atoll of West Texas is 70 to 90 miles across and 1,000 meters thick. C. Theories explaining the formation of coral reefs: Many scientists tried to explain the formation of coral reefs and gave their views which are discussed below in brief. a. Darwin’s subsidence theory: Darwin believed that the reef began as fringing reefs on a sloping shore. Then the island subsides very slowly, so slowly that the reef grows upward at about the same rate, naturally the island becomes small, the channel between the reef and land widens and thus the fringing reef transforms into a barrier reef. Further subsidence of the land till it sinks completely out of site results in the formation of an atoll (Fig. 18). This is substantiated by the fact that all the known coral reefs were in regions where a sinking of the land was known to have taken place or where there were evidences that it had probably occurred. b. Stutchbury’s volcanic crator theory: According to Stutchbury, atolls of the pacific were built upon the lost volcanoes. The crator of the volcano over the years got widened by water currents and became the lagoon, while its raised edges of the land were grown over by the coral reefs and formed atolls. This theory did not get much acceptance as diverse shapes of the atolls with limited depths of lagoons and the great number of craters in a single archipelago was present. c. Samper Murray solution theory: This theory states that calcareous skeletons of animals and other sediments form big mounds on the floor of the oceans. Over the time, these deposits grow to certain heights and corals grow on them and reach the water surface. Barrier reef is formed by the luxurious growth of coral at the outer edge while atoll is formed by dissolution of the inner coral rock. d. Submerged bank theory: This theory states that both barrier reefs and atolls grew upon pre-existing flat surfaces. Extensive coral growth occurred on a flat surface which got submerged in the water with the passage of time. Exposed regions formed the barrier reefs, while the shape of the atolls is obtained by the action of prevailing water currents and winds. Page 97 of 115 Many boring experiments were performed in different coral reefs by many scientists to find out the age and evolution of different coral reefs but no single theory could explain the formation of coral reefs. Reef formation is not the result of one mechanism as many different geological processes are occurring in the nature which might have led to the formation of these heavy structures. 5. Significance of corals: i. They show a high degree of physiological integration, division of labor, and perfect coordination with other groups of animals staying together, mutually benefiting each other in getting shelter, protection and food. Thus they constitute an ecologically important aquatic ecosystem. ii. They help in studying the evolution of the animals as fossils of the animals are preserved in the coral reefs over the years. iii. By studying the lines of growth on the fossils of some mollusk help in knowing about the seasonal fluctuations as the thickness of the growth line varies from season to season. iv. A few stony corals because of the presence of minute pores are used in surgical procedures as human capillaries can easily pierce through equal sized pores which are helpful in interconnecting two bones with each other. These corals are being used in the surgery for bone grafts and jaw surgery etc. v. Horseshoe shaped Atoll is the largest limestone reservoir for the oil production in North America. vi. Many of the corals are precious stones which are used in making jewelry and have aesthetic value. vii. Coral reefs are very hard structures and are an important source of mortar, cement, lime etc as they contain enough amount of CaCO3, therefore their rocks can be used for making roads and houses etc. viii. Reefs are also a rich source for medical formulations, used to treat a wide range of diseases like asthma, heart diseases, and viral, fungal and bacterial infections. It has been reported in 2006 that Yellow coral (Isis hippuris) collected off the coast of Okinawa island of Japan has yielded a compound that can slow down and possibly prevent virus replication and also treat cancer. ix. Lastly, corals act as affective buffers against erosion and storms occurring in the sea thus help in preventing tsunami disaster. 6. Coral crisis: There are about 2500 living species of coral many of them are becoming extinct due to change in the environment and global warming which is declining their number. Coral growth is dependent upon many environmental factors as well as the inhabitants of the coral reefs. Therefore, their growth is determined by the seasonal changes like wave action of water, harm caused by boring organisms and predators feeding on the coral polyps. These changes may be due to natural factors and man made factors. Corals and coral reefs are extremely sensitive to slight changes in the reef environment and which may have detrimental effects on the health of entire coral colonies. There is a great loss to the species with the habitat loss and leads to the extinction of many species. Environmental factors change from season to season, therefore, coral formation and deposition of the calcareous skeleton also varies. Besides this, there are certain predators which feed on the coral forming animals, or form burrows inside the coral reefs and thus Page 98 of 115 cause harm to the corals and produce solid calcareous debris on which algae grows, thus sometimes preventing planulae larvae to form new colonies. A. Natural factors affecting coral growth: Few factors which have a great impact on the building of coral reefs are briefly given below. a. Temperature: Corals are found in the tropical seas where the temperature never falls below 200C at normal salinity. Therefore, they are absent from the region where temperature goes down due to cold ocean currents coming from Antarctic. A temperature of 370C favors the growth of bacteria and viruses within the coral reefs which damage the coral structures. b. Depth of the ocean: Corals are differently located at various depths of the ocean. As one goes down the ocean, temperature decreases and therefore they grow luxuriously at the depth of about 30 meters but are scarcely found at the depth of 90 meters and more. c. Availability of light and presence of algae: Light availability also plays an important role in the formation and growth of the corals. Their growth is comparatively more in shallow waters as compared to the regions where light penetration level is veryvery low. This vertical distribution is restricted to the hermatypic corals (corals having symbiotic algae i.e. dinoflagellates or zooxanthellae, growing in their endodermal cells) as they have a significant role in coral formation. These algae grow in the presence of sunlight, CO2, and water and release O2 and carbohydrates. The carbohydrates thus produced are utilized as nutrients by the coral polyps while CO2 released by them is consumed by the algae, thus mutually benefiting each other and getting rid of the excretory wastes in a natural way. Therefore, hermatypic corals are restricted to shallow warm waters where light penetration is sufficient for their growth. In contrast deep sea corals i.e. ahermatypic corals have no zooxanthellae, and can grow up to the depth of 8000 meters They can grow even at 00C but are found growing in plenty within 5-100C. They can live without light. Deep sea corals are solitary, which settles at the bottom, a few are colonial and dendriform. d. Sea storms: There are several natural disturbances which cause significant damage to coral reefs e.g. hurricanes, tsunamis and storms, which bring large and powerful water waves which break apart large corals and scatter them into fragments. After the storm, these slow growing corals might easily be overgrown by quicker growing algae. In addition, these storms generally bring heavy rain which increases runoff and sedimentation e.g. latest disaster, the earth quake and tsunami of December 2004 caused a great damage in the Andaman and Nicobar islands. Nearly 3,500people were reported dead and thousands were left homeless. Extensive and beautiful coral reefs because of which these islands were famous had a great damage. These reefs were hit because of the submergence, the increased turbidity and the physical damage caused by the debris thrown back and forth by the devastating waves. In the Andaman waters, huge coral reefs were permanently exposed above the high tide line destroying them within weeks. There is a not only reduction in the number of corals but other associated coral reef inhabitants like fish, shrimps, flat worms and hermit crabs were also affected. B. Man made factors: There are several factors which are dependent upon the humans which contribute in destroying or damaging the coral reefs. Page 99 of 115 a. Increase in human population and development: Increase in human population is not only responsible to the depletion of natural resources but also causes harm to the coral reefs. As population is growing, there is a need for the development of housing societies and factories, with the result the amount of freshwater runoff increases carrying large amounts of sewage, nutrients, pollutants like insecticides and fumigants from the farm lands into the sea water. Sewage treatment facilities greatly increase the nutrients and microorganisms while large power plants discharge extremely hot water along into the coastal waters and increase the water temperature which may lead to white band disease (WBD) or black band disease (BBD). Increase in turbidity decreases the amount of light reaching the corals which may cause bleaching. Increase in nutrients may enhance the growth of other boring animals and sponges which outgrow the corals for space on the coral reefs. With the increasing human population, there is an increase in the demand for the sea food, including reef fish, gastropods, bivalves etc. Due to over fishing, overgrowth of the algae may take place which will not allow the planula larvae to settle and so the coral reefs are not able to establish themselves. Sometimes, explosives and poisons are used by the fishermen for the easy catch of the fish, which further damage the corals in those areas. b. Aesthetic value: Corals are very colorful and are also used in for making jewelry and as show pieces for decorations. People either collect pieces of coral themselves or buy them from the shops. Commercially they select most healthy colonies and sell them at higher rate, thus making a great loss to the coral reefs. c. Sea traffic: Coral reefs are also damaged by the leakage of fuel oils by large tankers into the water. Sometimes large oil carrying vessels sink in the sea and oil spillage cause mass destruction of the floating gametes and thus can effect coral reproduction and development. d. Predators: Boring chaetopods, gastropods, bacteria, viruses and starfish etc. cause damage to the coral reefs. It has been reported that there is a large star fish i.e. Acanthaster planci which take out its stomach on the coral and digest the living tissue layer and cause drastic damage to the coral populations. C. Coral Bleaching: Coral bleaching is the whitening of coral colonies due to the loss of symbiotic zooxanthellae from the tissues of polyps. This loss exposes the white calcium carbonate skeletons of the coral colony. Coral bleaching may be caused by the expulsion of the algae from the polyps either due to shortage of nutrients or algae may produce toxins under stress which affect the polyps. Coral bleaching may also be caused due to coral diseases, excess shade, increased levels of ultraviolet radiation, sedimentation, pollution, salinity changes, and increased temperatures. Coral bleaching because of the warming has destroyed more than 90% of coral around Seychelles. D. Coral Diseases: There are certain diseases which are reported to cause harm to the coral reefs e.g. white band disease (WBD) and black band disease (BBD) which kill coral tissue by growing in the form of a band around the coral and make them colorless and lifeless. BBD is caused by cyanophyte Phormidium corallyticum, while WBD is believed to be caused by a bacteria pathogen not known yet. These diseases weaken the Page 100 of 115 corals and damage them. BBD has a higher rate of infection in warmer water so seasonal temperatures affect the spread of BBD. The exact methods by which these diseases are transmitted are unknown. There are a great number of threats to coral reefs, and most of the threats are related to humans directly or indirectly. Work must be done quickly to protect our threatened aquatic ecosystem. There is a great need to enforce strict rules and regulations to ensure that proper techniques of fishing are used without disturbing the coral life. Rules are of no use unless people are properly educated throughout the world so that they are willingly ready to protect natural resources and so the coral reefs. Recently, Japan has begun planting baby coral on a Pacific atoll, to save sinking islets. It is a multi-million-dollar project in which several plants of juvenile corals near the uninhabited islets will be planted. So, it is a beginning not only to protect the islets from disappearance but also to preserve the coral community. Page 101 of 115 Mouth Pinnate tentacle Opening of pinnate tentacles Siphonoglyph Solenia Epidermis Coelenteron Spicules Coenenchyma Soft coral Gastrodermal tubes 1a 1b Fig. 1a: Octocorallian polyp and coral structure. 1b: Different types of spicules found in mesogloea of octocorallian corals. Page 102 of 115 Expanded polyps (anthocodia) Polyps withdrawn within Enteron Stalk of the colony Base for the attachment Fig. 2: Alcyonium coral colony Polyps Pinnate tentacles Coenosteum Solenia Blind cylindrical cavities Fig. 3: Heliopora coral showing two polyps and coral skeleton in a section. Page 103 of 115 Expanded polyps Polyp openings Vertical tubes Horizontal platforms Fig. 4: Tubipora, a part of the colony. Fully expanded polyps Pores in which polyps are withdrawn Base of the colony Fig. 5: Corallium, a part of the colony. Page 104 of 115 Anthocodia Main Branches Mesh formed by cross connections between smaller branches bearing polyps called anthocodia. Central axis Stalk Basal disc Fig. 6: Gorgonia colony Page 105 of 115 Mouth Epidermis Tentacle Gastrodermis Stomodaeum Gastrodermal canal of another polyp which is being budded off Mesenteries Ridges Sclerosepta Corallite Groove Basal plate 7a Theca Primary mesenteries Retractor muscles Pharynx Secondary mesenteries Endocoelic Sclerosepta 7b Fig. 7a: L.S. of a Hexacorallian coral showing the growth of sclerosepta alternating with mesenteries. 7b: Diagrammatic section of a polyp showing formation of sclerosepta within endocoels. Page 106 of 115 Tentacles Mouth Coelenteron Mesenteries Sclerosepta growing inwards Basal plate Sclerosepta Columella 8a 8b Fig. 8a. Fungia (a solitary coral) 8b. Diagrammatic section of a solitary coral showing sclerosepta and mesenteries. Corallites Coenosarc Polyps Fig. 9: Madrepora (Acropora), a part of the colony. Page 107 of 115 Polyps Corallites Solid and stony coral Fig. 10: Diagrammatic view of Astraea Page 108 of 115 11a Tentacles Mouth Retracted polyps 11b Fig. 11a: Antipathes, a single expanded polyp. 11b. A few polyps in retracted form. 12a 12b Capitate tentacles Dactylozooids Medusa Dactylopore Gastrozooid Gastropore Pores Coenosarcal canals Basal disc Degenerating canals Ampulla with medusa Tabulae Fig. 12a: Millepora colony ( A hydrozoan coral). Fig. 12b: Millepora in section (magnified view). Page 109 of 115 A polyp colony Thicket Coppice Bank Dead corals and shells Heavy coral growth zone 13a Reef edge Reef slope Sea ward slope Coral reefs Island 13b Fig. 13a: Development of coral reefs 13b: Generalized diagram of formation of coral reefs. Page 110 of 115 Island Fringing reefs Barrier reefs Lagoon Sea water Fig. 14. Fringing reefs and Barrier reefs Boulder zone Fringing reefs Inner flat Outer flat Reef edge Seaward slope Coral reef Island Coral reef Fig. 15. Section showing Fringing reefs Page 111 of 115 Low tide High tide Fringing reefs Lagoon Barrier reefs Coral reef Island Coral reef Fig. 16: Section through Barrier Reefs Water channels Atoll coral reefs Sea water Coral reef Coral reef Fig. 17. Section through Atoll Page 112 of 115 Coral reefs growing up Growing corals Subsiding island Island Island Lagoon Coral reefs Cora reefs Island Fig. 18. Darwin’s theory to explain origin of coral reefs. Page 113 of 115 VIII Bibliography 1. W. D. Russell-Hunter (1968), A biology of lower Invertebrates, The Macmillan Company, Collier-Macmillan limited, London. 2. M. D. L.Srivastava (1964), A text book of Invertebrate Zoology, Central book depot, Allahabad. 3. Robert D. Barnes (1986) Invertebrate Zoology, 5th Edition, ISBN 0-03022907-3 Saunders College Publishing, Address editorial correspondence to: 210 West Washington Square, Philadelphia, PA 19105 4. Edward E. Ruppert, Richard S. Fox, Robert D. Barnes (2004), Invertebrate Zoology, a functional evolutionary approach, 7th edition.. Thomson Learning Academic Resource Center i-800-423-0563. ISBN0-03-02982-7 5. R.S.K. Barnes, P. Calow, P. J. W. Olive, D.W. Golding, J.I. Spicer (2001), The Invertebrates: a Synthesis, 3rd edition; Blackwell Science Ltd Osney Mead, Oxford OX2 0EL, 25 John Street, London WCIN2BS 6. N.P.O. Green, G.W. Stout, D.J. Taylor (1996), Biological Science, 2nd edition Edited by R. Soper, Cambridge University Press, 7. Adam Sedgwick (1990) A student’s text book of zoology, Vol. I The Protozoa, by professor J. S. Dunkerley, Manchester University, low price publication. 8. T. Jeffery Parker and William A. Haswell, (1962) A text book of zoology, 6th edition, vol. I Macmillan & Co.Ltd, New York, St. Martin Press. 9. Richard A. Boolootian and Karl A. Stiles (1976), College Zoology, 9th edition. Macmillan Publishing Co., Inc. New York, Collier Macmillan Publishers, London. 10. Jan A. Pechenik (2002), Biology of the Invertebrates, 4th edition. Tata McGraw-Hill Publishing Company Limited, New Delhi. 11. E. N. K. Clarkson(1986), Invertebrate Palaeonotology and evolution 2nd edition, Allen & Unwin(Publishers) Ltd, 40 Museum street, London WCIAILU, UK 12. Ashok Verma (2005), Invertebrates Protozoa to Echinodermata. Narosa publishing house, 22, Darya Ganj, Delhi Medical association Road, New Delhi 110002.Brown, B.E., Odgen, J.C. 1993. Coral Bleaching. Scientific American, 269:64-70. 13. Pechenik, J. A. 1991. Biology of the Invertebrates. Wm. C. Brown Publishers, Dubuque, IA, pp. 91-92. 14. Rowan, R. and D. A. Powers. 1991. A Molecular Genetic Classification of Zooxanthellae and the Evolution of Animal-Algal Symbioses. Science, Vol. 251:1348-1351. Page 114 of 115 15. Sebens, K.P., Johnson, A.S. 1991. Effects of Water Movement on Prey Capture and Distribution of Reef Corals. Hydrobiologia, 226:91-101. 16. Wilkinson, C. R. 1987. Inter ocean Differences in Size and Nutrition of Coral Reef Sponge Populations. Science, Vol. 236:1654-1657. IX Acknowledgements I would like to express my deep thanks towards Dr. Narender, NISCAIR, for the confidence he showed in me and gave me this opportunity to contribute in E-Books for the vast subject of Zoology. I am also indebted to Ms. Shailley Anand, who showed a great patience in drawing the figures included in this chapter. I earnestly thank my daughters Shalini and Deeksha who kept me updated on my computer skills. I would also like to appreciate the technical support provided by my brother Mr.Dinesh Sharma. I express my gratitude to Dr. Rajendra Prasad, Principal, Ramjas College, University of Delhi, for all the encouragement during the course of writing these chapters. My thanks are also due to my husband, Prof. Rup Lal for his continual support and motivation in all the challenges that I have faced. Page 115 of 115