Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
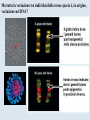
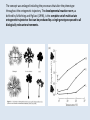
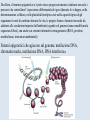
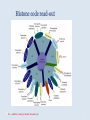
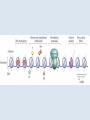
Epigenetica, un secondo codice genetico C. Soave La parola “development- sviluppo” è una metafora: suppone lo svolgersi di un qualcosa che è già tutto presente. Si considera lo sviluppo in termini dell’azione di geni. I geni nello zigote determinano lo stato finale dell’organismo, mentre l’ambiente nel quale lo sviluppo avviene è l’insieme delle condizioni che permettono ai geni di esprimere se stessi, analogamente ad una pellicola fotografica impressionata che produce l’immagine che è immanente in essa quando è posta in contatto con l’agente chimico necessario per lo sviluppo. Nel 700 divampava la polemica tra preformisti ed epigenetici. I preformisti sostenevano che l’organismo adulto era contenuto in miniatura nelle cellule germinali. Al contrario, per la teoria dell’epigenesi l’organismo si forma durante l’embriogenesi. Nel 20° secolo ha vinto il preformismo. Una sola specie umana, e non siamo tutti uguali: ci sono differenze morfologiche (più o meno chiare) fra individui Ma quanto siamo diversi? Quante basi del DNA lo sono? (totale genoma: 3,2 x 109 basi) Due cellule dello stesso individuo 0/1000 Due gemelli identici 0/1000 Due di noi a caso 1/1000 Uno di noi e uno scimpanzè 10-40/1000 Uno di noi e una banana 650/1000 Le differenze genetiche fra individui nella nostra specie sono le più basse di tutti i primati Albero evolutivo del cromosoma X: Kaessmann et al. (2001). Ma tutta la variazione tra individui della stessa specie è, in origine, variazione nel DNA? Phenotypic accomodation, plasticity polifenismo L’ontogenesi di un organismo è la conseguenza dell’interazione tra i suoi geni, la sequenza temporale delle situazioni ambientali che attraversa durante la sua vita, le interazioni casuali tra le molecole all’interno delle sue cellule: sono tutte queste interazioni che devono essere incorporate nel processo ontogenetico: genetic accomodation o capacitation. Un po’ di storia L’effetto Baldwin Nel 1896, Mark Baldwin, uno psicologo americano specializzato nello studio dello sviluppo della mente nell’infanzia, scrisse un articolo, pubblicato su American Naturalist, dal titolo “A new factor in evolution”. Il nuovo fattore, passato alla storia con il nome di “effetto Baldwin”, prendeva in considerazione la ben nota capacità degli organismi viventi di adattarsi, durante la loro vita, a nuove condizioni ambientali, a volte anche molto avverse. Questa capacità, sosteneva Baldwin, permette agli organismi di sopravvivere e riprodursi nel nuovo ambiente, generando figli capaci anch’essi di adattarsi e di sopravvivere nelle nuove condizioni fino a che, col passare delle generazioni, si selezionano quei varianti (mutanti) che rendono stabilmente incorporati nel programma genetico di sviluppo quei caratteri inizialmente emersi a seguito dell’adattamento Lo stesso concetto, sostenuto nello stesso anno da Lloyd Morgan e Osborn (finiti però nel dimenticatoio della storia scientifica), è efficacemente sintetizzato dalla West-Eberhard con l’aforisma “ phenotype first, genotype follows”. Si badi bene però non nel senso lamarckiano dell’ereditarietà dei caratteri acquisiti (Baldwin, Lloyd Morgan e Osborn erano fortemente antilamarckiani), perché quegli adattamenti fenotipici che permettono agli organismi di sopravvivere generazione dopo generazione non sono ereditati, ma spianano la via per quelle mutazioni genetiche che rendono stabilmente espressi gli adattamenti fenotipici: il costo del processo originale dell’adattamento è successivamente ridotto dall’instaurarsi del cambiamento genetico che permette l’evoluzione darwiniana. Perchè le idee di Baldwin non furono accettate dai Darwinisti Nella ipotesi darwiniana il cambiamento fenotipico può avere conseguenze evolutive (cioè può essere trasmesso) solo se la sua espressione è associata con una preesistente variabilità genetica: l’effetto Baldwin invece può operare senza questa variabilità preesistente. Johannsen: “The genotype-conception [of heredity] is an ‘‘ahistoric’’ view of the reactions of living beings.... I suggest that it is useful to emphasise this‘‘radical’’ ahistoric genotype-conception of heredity in its strict antagonism to the phenotypeview” (Johannsen 1911, The genotype conception of heredity.) Simpson “the idea [di Baldwin] was theoretically consistent, that is not inconsistent with the Modern Synthesis but the phenomenon could be proven to occur” Il ”sapore lamarchiano” delle idee di Baldwin contrastava con la dottrina di Auguste Weismann secondo la quale la trasmissione ereditaria avviene solo attraverso le cellule germinali ( Weismann, A.:1893, The germ-plasm: a theory of heredity) Weismann comunque pensava che le condizioni esterne fossero essenziali per produrre variazioni nelle cellule germinali e sosteneva “the inheritance of acquired germ-plasm variations” ed il meccanismo era descritto come una “definitely directed variation” . The modern synthesis Princeton Conference on Genetics, Palaeontology and Evolution January 1947 Assumptions of the Modern Synthesis (1940-1990 view) 1. Heredity occurs through the transmission of germ-line genes. Hereditary variation is equated with variation in DNA base sequence. There are no inherited non-DNA variations that cannot be explained in terms of DNA variations. 2. Hereditary variation is the consequence of (i) the combinations of preexisting alleles that are generated by the sexual processes; and (ii) new variations (mutations) that are the result of accidental changes in DNA. Hereditary variation is not affected by the developmental history of the individual. There is no “soft inheritance”. 3. Heritable variations have small effects, and evolution is typically gradual. Through the selection of individuals with phenotypes that make them slightly more adapted to their environment than others individuals are, some alleles become more numerous in the population. With a few exceptions, macroevolution is continuous with microevolution, and does not require any extra considerations beyond those operating during microevolution. Assumptions of the Modern Synthesis 4. The ultimate unit of selection is the gene. Although genes interact and the interactions are often non-linear, the additive fitness-effects of single genes drive evolution by natural selection. The geneticdevelopmental network and the phenotype it generates is not heritable and cannot be a unit of evolution. 5. Morphological evolutionary innovations, are the results of cumulative gene mutations. Generic physical-chemical processes are merely a constraint specifying the boundaries of physically possible forms. Physically-based plasticity is not a primary generative cause of morphological and physiological innovations. It is merely a boundary condition. The targets of selection are individuals that are, at all times, welldefined entities. Although conspecifics in groups interact and may coevolve with each other as well as with their symbionts and parasites, group selection or community selection is rare. The community is rarely considered as a target of selection. 6. 7. Evolution occurs through modifications from a common ancestor, and is based on vertical descent. Horizontal gene transfer has minor significance – it does not alter the basic branching structure of phylogenetic divergence. The main pattern of evolutionary divergence is, at all times and for all taxa, tree-like, not web-like. Baldwin’s ideas however were not completely lost and, from time to time, they were taken up in the past century. The concept of plasticity, introduced by Baldwin, was taken up by Woltereck (1909) with the concept of “reaction norm”: when an individual phenotype varies as a continuous function of some environmental variable, its reaction norm is the full set of phenotypic responses to that variable, usually expressed as a curve on a graph of phenotype value versus values of the environmental variable. The concept was enlarged including the processes that alter the phenotype throughout the ontogenetic trajectory. The developmental reaction norm, as defined by Schlichting and Pigliucci (1998), is the complete set of multivariate ontogenetic trajectories that can be produced by a single genotype exposed to all biologically relevant environments. Waddington: la genetic assimilation e l’epigenetic landscape He began with a stock of flies that produced a novel phenotype in low frequency when subjected to heat shock at an early larval stage. Selection for this heat shock-induced phenotype resulted in a much larger fraction of the population expressing the phenocopy, indicating that the plasticity of the trait was heritable. The most surprising observation came out after several generations of selection when the novel phenotype was expressed in the absence of the heat shock! Genetic assimilation is “a process by which a phenotype character, which initially is produced only in response to some environmental influences, becomes, through a process of selection, taken over by the genotype, so that it is found even in the absence of the environmental influence, which had at first been necessary” (Waddington, 1961. The genetic assimilation. Adv. Genet. 10 257-290. Waddington (an embryologist and a geneticist) introduced also the term epigenetics to refer to the study of the ‘‘causal mechanisms’’ by which ‘‘the genes of the genotype bring about phenotypic effects” during development, that is epigenetics is the conduit connecting the genotype to the phenotype during the entire life cycle. Furthermore, the Waddington concept of epigenetics invoked a causal role for the environment in developmental processes, and in the induction of alternative phenotypes (the so called epigenetic landscape, that is the “ensemble” of the possible phenotypes that a genotype can produce when exposed to different environments) Nanney (Epigenetic control systems. 1958 Proc. Natl. Acad. Sci.) introduce una distinzione concettuale tra ciò che mantiene la specificità dell’informazione, cioè il meccanismo di replicazione dell’informazione e i meccanismi ausiliari con diverso principio operativo, implicati nel determinare quali sono le specificità che devono essere espresse in ogni cellula. Individua quindi un sistema genetico primario (DNA e meccanismi di replicazione) e un sistema epigenetico deputato al controllo dell’espressione del primo. Prove dell’esistenza di un sistema epigenetico indipendente dal sistema genetico primario sono: -Cells with the Same Genetic Material May Manifest Different Phenotypes. -The Genetic Potentialities of a Cell Are Expressed in Integrated Patterns.-In the sequential induction of enzymes, the expression of one specificity may involve the eventual expression of a series of specificities. -Particular Patterns of Expression Can Be Specifically Induced. -Epigenetic Systems Show a Wide Range of Stability Characteristics. The view that differentiated cells in vertebrates do not "de-differentiate" suggests that some epigenetic systems have great stability. -Epigenetic Devices May Be Localized in the Nucleus or in the cytoplasm. Da allora, il termine epigenetica si è però esteso progressivamente a indicare non solo i processi che controllano l’espressione differenziale dei geni (durante lo sviluppo, nella determinazione cellulare, nella plasticità fenotipica cioè nella capacità tipica degli organismi viventi di cambiare durante la vita, le proprie forme e funzioni in modo da adattarsi alle condizioni imposte dall'ambiente) agendo sul genoma (senza modificare la sequenza di basi), ma anche sui sistemi informativi extragenomici (RNA, proteine, metabolismo, interazioni ambientali). Sistemi epigenetici che agiscono sul genoma: metilazione DNA, chromatin marks, metilazione RNA, RNA interference DNA methylation • • DNA methylation is mediated by DNA methyltransferases that target cytosines in all contexts (CG, CHG and asymmetric CHH; where H represents A, T, or C) DNA methylation occurs preferentially at repeat and transposon sequences, and DNA methylation particularly in 5’ control regions is generally associated with transcriptional repression and/or silencing of genes. MET1/VIM1 mCG DDM1 ATPase CMT DRM2 mCHG mCHH 22 Histone Code: the second code Strahl and Allis, 2000 Nature 403 41-45 Histone tails are modified in different ways and these modifications are associated with different cellular functions, because modifying enzyme necleosomes remodeling complexes and downstream effectors complexes read each others marks on the histone tails. 11/03/2016 Histone code 23 Histone code read-out 11/03/M20u1n6shiet al. (2009) J. Genet. Genomics 36 25