Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
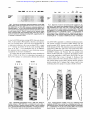
From www.bloodjournal.org by guest on November 7, 2014. For personal use only. 1994 84: 3205-3211 Molecular mapping of the Cromer blood group Cra and Tca epitopes of decay accelerating factor: toward the use of recombinant antigens in immunohematology MJ Telen, N Rao, M Udani, ES Thompson, RM Kaufman and DM Lublin Updated information and services can be found at: http://www.bloodjournal.org/content/84/9/3205.full.html Articles on similar topics can be found in the following Blood collections Information about reproducing this article in parts or in its entirety may be found online at: http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requests Information about ordering reprints may be found online at: http://www.bloodjournal.org/site/misc/rights.xhtml#reprints Information about subscriptions and ASH membership may be found online at: http://www.bloodjournal.org/site/subscriptions/index.xhtml Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036. Copyright 2011 by The American Society of Hematology; all rights reserved. From www.bloodjournal.org by guest on November 7, 2014. For personal use only. Molecular Mapping of the Cromer Blood Group Cr" and Tc" Epitopes of Decay Accelerating Factor: Toward the Use of Recombinant Antigens in Immunohematology By Marilyn J. Telen, Neeraja Rao, Manisha Udani, E. Scott Thompson, Richard M. Kaufman, and Douglas M. Lublin Cromer blood groupantigensresideon the complement regulatory protein decayacceleratingfactor(DAF,CD55). This glycosyl-phosphatidylinositol-anchored glycoprotein is widely dim-buted, especially among cell types in contact with plasma. Numerous Cromer blood group antigens have been defined using alloantibodies induced by transfusion or pregnancy. However,few pairs ofantithetical antigens have been described in this system, presumably because of the rarity of the low-frequency alleles. Analysis of polymerase chain reaction-amplified genomic DNA showed that the Pro substitution in short Cr(a-) phenotype has a Ala'= consensus repeat 4 (SCR4)of DAF, and the Tc(a-b+) phenotype has a Arg" --t Leu substitution in SCRl of DAF. The locations ofCr' and Tc' epitopes were confirmed by analysis of Chinese hamster ovary cell transfectants expressing a Cr(a-) allele-specifictransfectantandachimeric protein containing only SCRl ofDAF, respectively. Overall, these studies further show the usefulness of an approach based on recombinant proteins in mapping blood group antigen epitopes and identifyingblood group antibodies. 6 1994 by The American Society of Hematology. T The aim of this study was to determine the genetic and biochemical bases of the Cr(a-) and Tc(a-) phenotypes and to map the C f and Tc" epitopes using recombinant proteins. -+ HE CROMER BLOOD GROUP system comprises at least 10 individual antigens.' Of these, most are highfrequency antigens identified when a rare individual lacking the antigen produced an antibody after exposure to blood products expressing that antigen; only three low-incidence antigens have been identified by human sera. The antigens of the Cromer blood group system reside on decay accelerating factor (DAF, CD%), a glycosyl-phosphatidylinositol-anchored protein with complement regulatory activity.' The high-frequency D$ antigen was the first Cromer antigen to be deciphered at the molecular level, and the Dr(a-) phenotype is caused by a single amino acid substitution, Ser'65 Leu, in The C f and Tc" antigens are the two highincidence Cromer antigens to which alloantibodies are most frequently identified, but they have not been previously characterized at the molecular level. Unlike the Dr(a-) phenotype, the Cr(a-) and Tc(a-) phenotypes have not been associated with weakened expression of other Cromer antigens. The Cr(a-) phenotype was the first described and remains the most common polymorphism in this blood group sysThe Cr(a-) phenotype occurs in less than 1 in 1,000 individuals, and in the United States, nearly all Cr(a-) individuals are African-Americans. Although rare, human antiC f has been associated with significantly accelerated destruction of transfused red blood cells (RBCs).' Its possible role in hemolytic disease of the newborn is unclear; no case of hemolytic disease of the newborn has been reported to have been caused by anti-Cf', and one of the propositi included in this study delivered several children without complication. No antibody to the product of the Cr(a-) DAF allele has been described. The Tc(a-) phenotype is also rare. First recognized after the identification of two women who made anti-Tc" after pregnancy,6 the Tca antigen occurs in greater than 99% of all populations studied. In African-Americans, absence of Tc" is associated with expression of the Tcb allele, whereas in whites, the Tc(a-) phenotype is associated with expression of Tc". The incidence of the Tcballele in African-Amencans is *5%.' Little documentation of the clinical significance of anti-Tc" exists; although in vitro these antibodies give a positive mononuclear phagocyte assay,' hemolytic transfusion reactions and hemolytic disease of the newborn caused by anti-Tc" have not been described. Likewise, antiTcb and anti-Tc" are of unclear clinical importance. Blood, Vol 84, No 9 (November l), 1994: pp 3205-3211 MATERIALS AND METHODS Blood cells and sera. Blood from Cr(a-), Tc(a-b+) and normal Cr(a+) Tc(a+) donors was collected using tripotassium EDTA as anticoagulant. Erythrocytes (E) were then stored in acid citrate dextrose at 4°C for up to 1week and were washed inphosphate-buffered saline (PBS, pH 7.4) before use. Samples of whole blood collected from three unrelated Cr(a-) donors were kindly contributed by John Judd (University of Michigan, Ann Arbor) and by Rebecca Bullock (American Red CrossCarolinas Region, Durham, NC). Samples from the Tc(a-b+) donor were provided by M. Yacor and M. Reid (New York Blood Center, New York, NY), and anti-Tc"was provided by John Moulds (Gamma Biologicals, Houston, TX). Additional antisera and Cromer-variant RBCs from the Duke University Immunohematology Laboratory (an AABB-certified reference laboratory) were also used for confirmation of cell phenotype and antibody specificity. Cell typings and antibody identification were performed using standard agglutination testing. The following murine monoclonal antibodies (MoAbs) were used: M75 and M160 antimembrane cofactor protein (MCP)*; 1H4, 15B10, BRIC 230, and 3.3-136 anti-DAF?." Polyclonal anti-DAF was also used in some experiments? DNA isolation and analysis. To isolate leukocyte DNA, samples From the Department of Medicine, Division of Hematology/Oncology, Duke University Medical Center, Durham, NC; and the Departments of Pathology and Medicine, Division of Laboratory Medicine, Washington University, St Louis, MO. Submitted February 28, 1994; accepted June 29, 1994. Supported by Grants No. HW3572, HZ44042 (to M.J.T.), and A115322 (to D.M.L) fromthe National Institutes of Health (NIH), and a grant from the National Blood Foundation (to D.M.L.). M.J.T. is the recipient of Research Career Development Award No. HLO2233 lfrom NIH). Address reprint requests to Marilyn J. Telen, MD, Box 3387, Duke University Medical Center, Durham, NC 27710. The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advettisement" in accordance with 18 U.S.C.section 1734 solely to indicate this fact. 6 I994 by The American Society of Hematology. 0006-4971/94/8409-W.00/0 3205 From www.bloodjournal.org by guest on November 7, 2014. For personal use only. TELEN ET AL 3206 Table 1. Oligonucleotide Primers Used for Amplification of DAF Exons Exon SCRl SCRZ SCR3A SCRBB SCR4 SIT-A SIT-B, Sn-C Hydrophobic Primer Pair* T (anneal) 5"CCTTCAGTTCTGCmTGTCTCCC 5"CTATCAATTACTAGTCACTCCAAAGG 5"GGGTTAlTAGGGTCCAGATAA 5"GAGTTCTAGCATGAATGAAGGMGGG 5"CAAACAGCClTATATCACTC 1,200 5'-AACAATCTCACmAAG 5"AGAlTGATGTACCAGGTGGC 1,200 5"GAGTACTCAGCCTCACAATCTGAG 5"GCATCTmGTTGGTAATGCTG 5"CAACCCACATATAGACCGAGGG 5'-ACAGAGCAAGCAATGGC 5"ATlTGGGGlTGTTFCATGAA 5"CAGTGACTAATGGTCTC 2,100 5"GGAATATGGATTGTATAT 5"CCl"TGGCCAACCCCAAATTAACTG 5'-ACATTCCTAACACATClT Product Size (bp) 45°C 300 55°C 300 45°C 55°C 55°C 350 55°C 800 45°C 45°C 400 For eachprimer pair, the topprimer is in the coding direction, and the bottom primer is inthe noncoding direction. of anticoagulated whole blood were centrifuged and the buffy coat aspirated into a second tube. After several washes in PBS, cells were lysed in 10 mmollL TRIS-HCl (pH 8.0), 0.1 m o m EDTA, 0.5% sodium dodecyl sulfate (SDS) with 100 p@mLproteinase K (overnight at 42") and extracted with phenol. DNA was then precipitated from the aqueous phase with ethanol and stored in water. Analysis of the coding exons of DAF was performed as previously described,' using the primers listed in Table 1. Briefly, each exon was amplified by polymerase chain reaction (PCR) using Taq polymerase (Perkin Elmer-Cetus, Emeryville, CA). PCR reaction conditions were as follows: denaturation at 94°C for 2 minutes followed by 35 cycles of denaturation at 94°C for 1 minute, annealing at the temperature indicated in Table 1 for 1 minute, and extension at 72°C for 1 minute for products 4300 bp and 3 minutes for products >SO0 bp. The resultant product was subcloned into a plasmid vector (pUC19 or pBluescript KS+) and transformed into DHScu. Double-stranded sequencing was performed using Sequenase (US Biochemical, Cleveland, OH) according to manufacturer's recommendations, using either M13 universal primers or primers used during PCR amplification. In each case, at least two independent clones from each of two separate PCR amplifications were sequenced to confirm any variation from wild type. Construction and expression of wild-type and variant DAF. DAFMCP chimeric cDNA was made byPCR." The nucleotides encoding the 5' untranslated region, signal peptide, and SCRl of DAF were amplified from 1 ng of DAF cDNA plasmid," and the fragment encoding SCR2 through the 3'-untranslated region of MCP was amplified from MCP cDNA plasmid.13 These two fragments were ligated together in reading-frame in the vector pBluescript KS+, and the construct was confirmed by DNA ~equencing.'~ This DAF/ MCP chimera comprised amino acids -34 to +61 of DAF (numbered such that amino acid + 1 is the amino terminus of the mature protein) joined to amino acids 63 to 350 of MCP. The DAFMCP chimeric cDNA was subcloned into the pBSRaEN mammalian expression vector" and transfected into Chinese hamster ovary (CHO) cells byuseof the reagent Lipofectamine (GIBCO, Grand Island, NY). Specifically, 10 pg ofthe pBSRaEN vector containing the DAFMCP chimeric cDNA was mixed with 70 pg Lipofectamine in 3 mL Optimem (GIBCO) and then added to a subconfluent 10-cm plate of CH0 cells. After 4 hours incubation at 37T, 10 mL of complete medium containing fetal calf serum was added and the incubation continued. Approximately 24 hours later, 2-aminopurine (Sigma Chemical CO, St Louis, MO) was added to a final concentration of 10 mmol/L to increase expression of the transfected cDNA.'~After a further overnight incubation, the cells were solubilized for Western blotting. CH0 transfectant cell lines expressing a series of DAF deletion mutants, from which individual DAF domains have been removed, have been previously described? In addition, site-directed mutation of wild-type DAF to duplicate the nucleotide polymorphism found in the Cr(a-) donors' DNA was performed as previously described,' and mutated DAF was then transfected into CH0 cells, also as described. Cells transfected with vector only were used as a negative control in various experiments. Quantitation of DAF by radioimmunoassay. To ascertain the level of expression of DAF by Cr(a-) and Tc(a-) RBCs, antibody 3.3-136 wasused, along with other previously described MoAbs and P3 X 631Ag8 myeloma ascitic fluid as positive and negative control^.^' Briefly, RBCs collected and stored as described above were washed in PBS and resuspended at 4 X 10x/mL(as determined by an ELT8 automated cell counter, Ortho Diagnostics, Raritan, NJ). After cells were incubated with saturating amounts of antibody and washed, saturating amounts of 1251-labeledF(ab')* sheep antibody to mouse IgG (Amersham, Arlington Heights, E)were added to detect binding of first antibody. Cell-bound and unbound label were separated by centrifugation through phthalate oils, and bound label was counted in a gamma counter. Western blor analysis. RBC ghosts were prepared, solubilized under nonreducing conditions, and proteins separated by SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose as previously described." Lysates of stably transfected cell lines were made by adding 0.5 to 0.75 mL PBS, 1% Nonidet-P40 (Sigma), 0.2% phenylmethylsulfonyl fluoride (PMSF) to a 25-cm' tissue-culture flask containing a 75% to 100% confluentcell growth, previouslywashed several times withPBS. The lysate wasthen transferred to a microcentrifuge tube, centrifuged to pellet unsolubilized nuclear material, and stored at -80°C. Lysates weremixed with2x SDS gel loading buffer, andrun on similar gels. Immunoblotting with human antibodies was accomplished by first partially purifying the antibody by adsorption and elution onto RBCs as described." Most immunoblots were developed using an alkaline phosphatase-anti-IgG conjugate and the BCIPNBT chromogenic subAlternatively, Western blots were strate (Promega, Madison, W)." developed using the ECL chemiluminescent system (Amersham). Dot blot analysis. Ten-microliter lysate aliquots were applied to nitrocellulose and allowed to bind for 2 hours at 22°C. The membrane was then washed in TRIS-buffered saline and 0.05% Tween 20 (TBST) and blocked in TBST and 10% milk powder for at least 30 minutes at 22°C or overnight at 4°C. Membranes were then incubated with dilutions of human anti-CP, human anti-Tc",or rabbit anti-DAF sera made in TBST for at least 1 hour, washed in TBST, and then incubated with theappropriate alkaline phosphatase-linked anti-IgG. Detection of antibody binding was accomplished using a chromogenic substrate as described above for Western blots. RESULTS Expression of DAF by Cr(a-) and Tc(a-b+) erythroa vacytes. When tested by erythrocyte agglutination with riety of Cromerand anti-DAF antibodies, the Dr(a-) and Inab phenotypes demonstrate reduced or absent DAF expression of DAF, respectively. This is confirmed by radioimmun~assay?,~ The Cr(a-) and Tc(a-) phenotypes exhibit normal erythrocyte agglutination when tested with other Cromer blood group antibodies. We used radioimmunoassayquanto titate DAF expression on these cells. Erythrocytes from two Cr(a-) donorsandone Tc(a-b+) donorweretestedby From www.bloodjournal.org by guest on November 7, 2014. For personal use only. CPANDTc" EPITOPES OFDAF 3207 radioimmunoassay using MoAb 3.3-136, as well as other MoAbs. Binding of MoAb 3.3-136 to Cr(a-) and Tc(a-b+) E was not different from that of Cr(a+)Tc(a+) control cells (Table 2). In some experiments, other anti-DAF MoAbs reacted more strongly with Tc(a-b+) RBCs than with normal RBCs (data not shown). Thus, this limited sample suggests that there is no significant decrease in DAF expression in the Cr(a-) and Tc(a-b+) phenotypes, unlike that seen in Dr(a-) individuals.' Analysis of Cr(a-) and Tc(a-b+) RBCs by Western blotting (Fig I ) also showed thatthe migration of DAF expressed by Cr(a-) and Tc(a-b+) RBCs was comparable with wildtype DAF in SDS-PAGE, confirming that there was likely to be only a minor alteration of the DAF molecule in the Cr(a-) and Tc(a-b+) phenotypes. Mapping ofthe CY epifope. To map the location of the Cr' epitope on the DAF molecule, human anti-Cr' was tested for reactivity with C H 0 transfectants expressing various deleted forms of DAF. In both dot blots (Fig 2) and Western blots (data not shown), anti-Cl" reacted with all transfectants except those missing the DNA encoding SCR4 and those expressing vector DNA only. We then analyzed all exons encoding SCRs 1 through 4 using genomic DNA from one Cr(a-) individual. In the initial studies of the five exons encoding SCR domains, a single point mutation was found. This mutation was a G + C change in the 15th nucleotide of the exon encoding SCR4 (Fig 3); this change encodes an Ala'y3 + Pro substitution. Analysis of this exon in the DNA from two other Cr(a-) donors also showed this single point mutation. No other polymorphisms were identified in the DAF gene of any of the Cr(a-) donors. To confirm the molecular basis of the Cr(a-) phenotype, we created a DAF mutant cDNA with this single nucleotide substitution and expressed it in C H 0 cells. When anti-Cr' from several donors was tested against this Cr(a-)-type allele-specific transfectant and a previously reported Dr(a-) allele-specific tranfectant in dot blots (Fig 4) and Western blots (data not shown), anticl" failed to react withthe Cr(a-) allele-specific transfectant, whereas other anti-DAF antibodies reacted strongly with the Cr(a-) transfectant (data Table 2. Expression of DAF by Cr(a-) and Tc(a-b+) RBCs Phenotype Experiment no 1 Cr(a-) 1 Cr(a-) 2 Cr(a+) l Cr(a+) 2 Cr(a+ ) 3 Cr(a+) 4 Experiment no 2 Tc(a-b+) Tc(a+) 1 Tc(a+) 2 Tc(a+) 3 Tc(a+) 4 Tc(a+) 5 Specific Cpm Bound Using MoAb 3.3-136 Anti-DAF 2,514 2,175 2,022 2,451 3,008 2,364 A kD antid NHS anti-DAF NHS antiDAF 46 69 30 - 2 1 -4NI Clfa-) lnab antiTca B kD 97 69 - 30 - Fig 1. Western blot analysis of Crla-1 and Tcla-b+) erythrocyte membranes. The antibody used t o identify immunoreactive protein is indicatedabove each panel, while theRBC phenotype ofcells used t o prepare membrane proteins is shown beloweach lane. In A, neither Cda-) nor lnab ICromer-null) erythrocyte membrane proteins contained DAF reactive with human anti-Cr'. Normal human serum (NHS) was nonreactive with human erythrocytemembrane proteins. However, monoclonal anti-DAF reacted with normal (NI) Cr(a+) as well as Crla-) 70-kD DAF and showed that both are equally well expressed and migrate similarly in SDS-PAGE. Similarly, in B, antiTc" failed t o react with Tcla-) erythrocyte membrane proteins, but identified DAF in normal membranes; however, monoclonal anti-DAF reacted strongly with normalTc(a+) as well as Tc(a-b+) DAF, both of which show similarapparent molecular weights of 70 kD. not shown). Thus, these results confirm that the single G C mutation in the exon encoding SCR4 was responsible for the Cr(a-) phenotype. Mapping o f f h e Tc" epifope. Two examples of anti-Tc" were similarly tested by dot blot and Western blot for reactivity with transfected C H 0 cell lysates containing wild-type DAF or DAF deletion mutants. However, both antisera failed + 4,221 4.876 5,019 4.742 4.278 4,646 From www.bloodjournal.org by guest on November 7, 2014. For personal use only. TELEN ET AL 3208 Cr DAP V00 -ticr. -3 (a-) (a-) -4 DI RBC antiera Fig 2. Dot blot of transfectants expressing deletion mutant DAF cDNAs. RBCs of CH0cells transfected with vector only (Vec), normal DAF EDNA (DAF), or DAF cDNA constructs containing deletions of individual SCRs (-1, -2, etc) or the serinelthreonine-rich region (-SIT) were used t o prepare detergent extracts were then testedreactivfor ity with human anti-Cr'. Anti-Cr' reacted with all extracts except those containing CH0 cells transfected with vector only or with a construct lacking thesequence encoding SCR4, suggesting that the C? epitope is containedin SCR4. to react with DAF proteins missing SCRl (data not shown). Thus. we proceeded to PCR amplify, subclone, and sequence the exon encoding SCR I . This work led to identification of :I single point mutation in the exon encoding SCRI; a single G -t T change at position 55 in the exon encoding SCRl leads to an ArgIX Leu substitution (Fig 5). In addition, this mutation leads to creation of a new restriction enzyme recognition site for Stu I (AGGCCT). To confirm that the single nucleotide change identified in SCRl of DAF of the Tc(a-b+) phenotype corresponds to the TcVTc" epitope, we constructed a chimera that contains -+ mom1 T Q C Cr(a-) A T Q C ~ Fig 4. Reactivity of anti-Cr' with allele-specific transfectants. Human antiCr' was reacted with detergent extracts ofRBCs and CH0 cells transfected with vector only (Vec), normal DAF cDNA (DAF), cDNA constructs from whichsequence encoding SCR3 or SCR4 had been deleted (-3, -41, or cDNA constructs containing the G -C change in SCR4 found in Cr(a-) individuals or the C T polymorphism of SCR3 found in Dr(a-) individuals. Anti-Cr' reacted with all cells expressing DAF or DAF variants except those either missing SCR4 or containing theG C mutation in SCR4 found in Cr(a- j individuals. - - only DAF SCR 1 appended to a different membrane protein. The specific construct consisted of MCP lacking only the amino-terminal SCRl domain, whichwas replaced by the SCRl domain of DAF. This chimeric protein was transiently expressed in C H 0 cells, and lysates were analyzed by SDSPAGE and Western blot. To confirm that this protein was a DAFIMCP chimera, the Western blot was probedwith MoAbs to MCP and D A F as expected, the chimeric protein reacted with antibodies to MCP and to DAF SCRI, but not to DAF SCR3 (Fig 6, lanes 1 through 3). Probing of the chimeric protein blot with a human anti-Tc" eluate or a control normal human serum eluate showed that the chimeric protein contains the Tc" epitope (Fig 6, lanes 4 and 5 ) . thus confirming that this epitope is located in SCRl of DAF. Tc(a+) ATGC /H i\ C A T T T T T T G T T ns C A C C :/ " A A E\ A ;-". m I Fig 3. Nucleotide polymorphism of Cr(a-) DAF gene. Partial sequence analysis of DAF SCR4 from a normal Cr(a+) and a Cr(a-) individual is shown. Sequence analysis of SCR4 in three unrelated Cr(a-) propositi showed a single nucleotideG +C base-pair substitution. This mutation leads t o a proline-for-alanine substitution. Tc(a-) ATGC G A A A A C C A T T* C C G G A T G C C G G A A A A G G G G Fig 5. Partial sequence of SCRl of Tc(a-b+l individual. Partial sequences of SCRl from a normal Tc(a+) individual and a Tc(a-b+) individual are shown, showing a single point mutation comprising a G T substitution (T*) in the exon encoding SCR1. This mutation leads t o an arg leu substitutionin the mature protein. - - From www.bloodjournal.org by guest on November 7, 2014. For personal use only. CP AND Tc" EPITOPES OF 3209 DAF 105- I 2 3 4 5 Fig 6. Western blot ofDAF/MCP chimeric protein. A chimeric protein consistingof SCRl of DAF appended t o MCP (see Materials and Methods for details of construct) was expressed transiently in CH0 cells. A cell lysate was separated by SDS-PAGE and transferred t o nitrocellulose. Lanes containing identicallysates of transfected CH0 cells were probedwith theantibodies indicatedabove each lane and developed with a chemiluminescence technique. Antibodies: lane 1, mixture of M75and M160 murine monoclonalanti-MCP; lane 2, mixture of 15810 and BRIC230 murine monoclonalanti-DAF (specific for SCRl); lane 3, 1H4 murine monoclonal anti-DAF (specific for SCRB); lane 4, human anti-Tc' (eluted from antigen-positive RBCsl; lane 5, normal humanserum, similarly eluted. Human anti-Tc" was reactive with the chimeric protein containing only the first SCR of DAF and was not reactive withMCP (data not shown). DISCUSSION The C? and Tc" antigens, like other Cromer antigens, reside on DAF, a glycosyl-phosphatidylinositol-linked complement regulatory protein.' DAF (Fig 7) contains four consensus repeat regions at its N-terminus.I2Each SCR domain contains two internal disulfide bonds that presumably influence the conformation of the molecule. Between these repeat domains and the anchor lies a serine/threonine-rich domain that is highly 0-glycosylated. This study has identified the genetic and biochemical basis for the Cr(a-) and Tc(a-b+) phenotypes, in which an altered form of DAF is expressed. Unlike the Dr(a-) phenotype, in which expression ofan altered DAF molecule is associated with reduced DAF expression, the mutations responsible for the Cr(a-) and Tc(a-b+) phenotypes appear to produce no diminution of membrane surface DAF. Nevertheless, in each case, the molecule is sufficiently altered that Cr(a-) and Tc(a-b+) individuals produce antibody to the wild-type DAF molecule when exposed via transfusions or fetal/maternal hemorrhage. All antibodies in the Cromer system have similar characteristics in that they fail to react with RBCs pretreated with reducing reagents or with chymotrypsin, whereas all antibodies react with trypsin-treated cells.' The localization of the C? and Tc" epitopes to SCR domains (Fig 7) is consistent with existence of disulfide bonds in these regions and the known trypsin insensitivity of DAF function, which resides in SCRs 2 through 4.' Although the mutation responsible for the Cr(a-) phenotype is close to the SCR3-SCR4 junction, it is clear from the deletion mutation studies that only SCR4 is necessary for expression of the C? antigen. The combination of a consistent single base-pair substitution in three unrelated Cr(a-) individuals, along with evidence that DNA with a similar mutation produced by sitedirected mutagenesis encodes Cr(a-) DAF in transfected cells, provides a firm identification of the genetic and biochemical basis of this phenotype. Our studies using a single example of the Tc(a-b+) phenotype indicate that the Tc" epitope resides within SCRI. This conclusion is most firmly supported by results with the DAFNCP chimeric protein, which expressed only the SCRl domain of DAF and likewise expressed the Tc" antigen. Lack of reactivity of two anti-Tc' sera onlywith transfectants expressing DAF cDNA fromwhich SCRI-encoding sequence had been deleted also supports this conclusion. The SCRl location of Tc" is likewise supported by the finding of a single pointmutation in thatregion in an individual with the Tc(a-b+) phenotype. However, the unavailability DAF (Cromer) r Consensus Repeats I 0 - Dr(a-): Serls5 l ? Cr(a-): Ala193- 0-Glycosylation Region Leu Pro CHO-O- -0-CH0 CHO-O- -0-CH0 -0-CH0 -0-CH0 r I L Fig 7. Mutations responsible for rare Cromer blood group phenotypes are shown. The DAF protein contains four SCRs, a heavily 0glycosylated region, and a glycosyl-phosphatidylinositol anchor. All the mutations thus far found to be thebases of rare Cromer blood group phenotypesoccur in the SCRs. From www.bloodjournal.org by guest on November 7, 2014. For personal use only. 3210 TELEN ET AL of other individuals with this phenotype has not allowed confirmation of a similar mutation in unrelated Tc(a-b+) individuals. Thus far, the D f , Tc&, and Cr" epitopes reside in the SCR domains of DAF, and variant phenotypes all result from a single base-pair substitution (Fig 7). A single base-pair substitution that creates a stop codon in SCRl has been found in the original propositus of the Inab Cromer-null phenotype.lgWhen blood-group antigen polymorphisms are reviewed overall, it is clear that several, but byno means all of them, arise from single amino acid substitutions in the parent peptide. However, veryfew molecules other than DAF have been studied in this detail. The M and N antigens of glycophorin A correspond to paired polymorphisms of amino acids l and 5 (sed and gly', M; leu' and gIu5, N).*' However, recent work suggests that individual antisera may be relatively more dependent on the presence of a single amino acid and may require presence of neighboring oligosaccharides as we11.z',22However, the S and S antigens of glycophorin B result from a single amino acid substitution (metz9v th?')?' and the rare Webb phenotype of glycophorin C also results from a point mutation, although the resultant single amino acid substitution results in loss of an N-linked oligosa~charide.~~-*~ Nevertheless, it is clear that the immune system is capable of extremely fine discrimination. In the cases of the Cr(a-) and Tc(a-b+) phenotypes, no change in N-linked or 0-linked glycosylation would be anticipated from the amino acid substitutions found. Identification of unusual antibodies such as anti-Cr"or anti-Tc" have traditionally relied on the availability of appropriate nonreactive, phenotypically characterized RBCs as reagents. However, many hospital-based blood banks and transfusion services do not have such rare reagents available. Thus, patients with such antibodies may face considerable delay before transfusion, evenwhen the rare blood type would be available for transfusion (usually from a centralized depository of phenotypically rare blood). Thus, one advantage of identifying the molecular basis for blood-group antigens is the prospect of using genetically engineered reagents to identify antibodies with unusual specificities." Nonhuman cells expressing recombinant blood-group antigen-bearing proteins could also be useful in complex cases where many alloantibodies are present; in such cases, expression of limited numbers of antigens would be helpful in determining the individual antigen specificities present. In the case of the Cromer blood group, testing of an alloantiserum versus the C H 0 transfectant expressing human DAF would give anunequivocal answer to whether any antibodies to high-frequency Cromer antigens are present, regardless of the presence of additional antibodies such as Rh. The ability to use both allele-specific transfectants and transfectants containing cDNA with deleted domains or chimeric cDNA constructs in assays using human blood group antisera and solubilized cell preparations bodes well for efforts to create non-RBC-dependent methods of analyzing bloodgroup antisera. ACKNOWLEDGMENT The authors gratefully acknowledge the technical assistance of Nicole Anderson and Julie Lapp. REFERENCES l. Storry JR: Serology and genetics of the Cromer blood group system, in Moulds JM, Laird-Fryer B (eds): Blood Groups: Chido/ Rodgers, KnopslMcCoyNork and Cromer. Bethesda, MD, American Association of Blood Banks, 1992, p 31 2. Telen MJ, Hall SE, Green AM, Moulds JJ, Rosse WF: Identification of human blood group antigens on decay-accelerating factor (DAF) and an erythrocyte phenotype negative for DAF. J Exp Med 167:1993, 1987 3. Lublin DM, Thompson ES, Green AM, Levene C, Telen MJ: Dr(a-) polymorphism of decay accelerating factor: Biochemical, functional and molecular characterization and production of allelespecific transfectants. J Clin Invest 87:1945. 1991 4. McCormick EE, Francis BJ, Gelb AB: A new antibody apparently defining an allele of Go": 18th Annual Meeting Program, American Association of Blood Banks, Chicago, IL, 1965, p 59 5. Stroup M, McCreary J: CP, another high frequency blood group factor. Transfusion 15:522, 1975 (abstr) 6. Laird-Fryer B, Dukes C, Walker EM Jr, Glassman A, Moulds JJ: Tc": A new high incidence blood group factor. Transfusion 20:631, 1980 (abstr) 7. Lacey PA, Block UT, Laird-Fryer BJ, Moulds JJ, Bryant LR, Giandelone JA, Linnemeyer DR: Anti-Tcb, an antibody that defines a red cell antigen antithetical to Tc". Transfusion 25:373, 1985 8. Seya T, Hara T, Matsumoto M, Akedo H: Quantitative analysis of membrane cofactor protein (MCP) of complement. High expression of MCP of human leukemia cell lines, which is regulated during cell differentiation. J Immunol 145:238, 1990 9. Coyne KE, Hall SE, Thompson ES, Arce MA, Kinoshita T, Fujita T, Anstee DA, Rosse WF, Lublin DM: Mapping of epitopes, glycosylation sites, and complement regulatory domains in human decay accelerating factor. J Immunol 149:2906, 1992 10. Bobolis KA, Moulds JJ, Telen MJ: Isolation of the JMH antigen on a novel phosphatidylinositol-linked humanmembrane protein. Blood 79:1574, 1992 1 1. Saiki RK, Gelfand DH, Stoffel S, Scharf SJ, Higushi R, Horn GT, Mullis KB, Erlich HA: Primer-directed enzymatic amplification ofDNAwith a thermostable DNA polymerase. Science 239:487, 1988 12.Medof ME, Lublin DM, Holers VM, Ayers DJ, Getty RR, Leykam JF, Atkinson JP, Tykocinski ML: Cloning and characterization ofcDNAs encoding the complete sequence of decay accelerating factor of human complement. h o c NatlAcadSci USA 84:2007. 1987 13. Lublin DM, Liszewski MK, Post TW, Arce MA, Le Beau MM, Rebentisch MB, Lemons RS, Seya T, Atkinson JP: Molecular cloning and chromosomal localization of human membrane cofactor potein (MCP). Evidence for inclusion in the multigene family of complement-regulatory proteins. J Exp Med 168:181, 1988 14. Tabor S, Richardson CC: DNA sequence analysis with a modified bacteriophage T7 DNA polymerase. Proc Natl Acad Sci USA 844767, 1987 15. Shenoy-Scaria AM, Kwong J, Fujita T, Olszowy MW, Shaw AS, Lublin DM: Signal transduction through decay-accelerating factor. Interaction of glycosyl-phosphatidylinositol anchor and protein tyrosine kinases ~ 5 6 ' 'and ~ ~69"".J Immunol 149:3535, 1992 16. Kaufman RJ, Muflha P: Translational control mediated by eucaryotic initiation factor-2 is restricted to specific mRNAs i n transfected cells. Molecular Cell Biology 7:1568, 1987 17. Chow FL, Telen MJ, Rosse WF: The acetylcholinesterase defect in paroxysmal nocturnal hemoglobinuria: Evidence that the enzyme is absent from the cell membrane. Blood 66:940, 1985 From www.bloodjournal.org by guest on November 7, 2014. For personal use only. Cr" AND Tc" EPITOPES OF DAF 18. Telen MJ, Paker TJ, Haynes B F Human erythrocyte antigens 11. The In(Lu) gene regulates expression of an antigen on an 80kilodalton protein of human erythrocytes. Blood 64599, 1984 19. Lublin DM, Mallinson G, Poole J, Reid M E , Thompson ES, Ferdman BR, Telen MJ, Anstee DJ, and Tanner MJA. Molecular basis of reduced or absent expression of decay accelerating factor in Cromer blood group phenotypes. Blood 84:1276, 1994 20. Dahr W, Uhlenbruck G, Janssen E, Allen FW: Different Nterminal amino acids in the M,N glycoprotein from MM and NN erythrocytes. Hum Genet 35:335, 1977 21. Blackall DP, Ugorski M, Pahlsson P, Spitalnik SL: Determination of the fine specificity of antigen-antibody interactions using mutants of recombinant glycophorin A. Transfusion 32:24S, 1992 (abstr, suppl) 22. Judd WJ, Issitt PD, Pavone BG, Anderson J, Aminoff D: Antibodies that define NANA-independent MN-system antigens. Transfusion 19:12, 1979 23. Dahr W, Beyreuther K, Steinbach H, Gielen W, Kruger J: 321 1 Structure of the Ss blood group antigens. 11. A methionine/threonine polymorphism within the N-terminal sequence of Ss glycoprotein. Hoppe Seyler Z Physiol Chem 3612395, 1980 24. Reid ME, Shaw MA, Rowe G, Anstee DJ, Tanner MJA: Abnormal minor human erythrocyte membrane sialoglycoprotein p in association with the rare blood-group antigen Webb (Wb). Biochem J 232:289, 1985 25. Telen MJ, Le Van Kim C, Guizzo ML, Cartron J-P, Colin Y: Erythrocyte Webb-type glycophorin C variant lacks N-glycosylation due to an asparagine to serine substitution. Am J Hemato1375 1, 1991 26. Chang S, Meid ME, Conboy J, Kan W, Mohandas N: Molecular characterization of erythrocyte glycophorin C variants. Blood 77:644, 1991 27. Telen MJ: New and evolving techniques for antigen and antibody identification, in Nance ST, (ed): Alloimmunity: 1993 and Beyond. Bethesda, MD, American Association of Blood Banks, 1993, p 117