Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
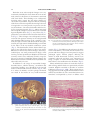
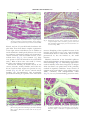
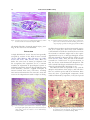
J. Comp. Path. 2008, Vol. 139, 24e33 Available online at www.sciencedirect.com www.elsevier.com/locate/jcpa Histopathology of the Alarm Reaction in Small Odontocetes D. F. Cowan*,†,‡,x and B. E. Curryk Department of Pathology, University of Texas Medical Branch, Galveston, TX, † The Marine Biomedical Institute, University of Texas Medical Branch, Galveston, TX, ‡ Department of Marine Biology, Texas A&M University at Galveston, TX, x Texas Marine Mammal Stranding Network, Galveston, TX and k Southwest Fisheries Science Center, 8604 La Jolla Shores Drive, La Jolla, CA, USA * Summary Pathological changes in the organs and tissues of beach-stranded, net-caught or captive small odontocete cetaceans (whales and dolphins) are reported. These changes include contraction band necrosis of cardiac and smooth muscles, smooth muscle spasm, ischemic injury to the intestinal mucosa (especially the mucosa of the small intestine) and acute tubular necrosis (ATN) of the proximal tubules of the nephron. Spastic contraction of terminal bronchiolar muscular sphincters was also observed. The changes are consistent with multisystemic injury caused by massive release of endogenous catecholamines or vasospasm leading to ischemic injury, followed by reperfusion and reperfusion injury. The histopathological findings suggest that the reflex response of an odontocete to any major perceived threat (the ‘‘alarm reaction’’) is to activate the physiological adaptations to diving or escape to an extreme or pathological level, resulting, if greatly prolonged, in widespread ischemic injury to tissues. These observations may explain why these species die abruptly from handling or transportation and why the mortality of highly stressed beach-stranded animals is very high. Ó 2008 Elsevier Ltd. All rights reserved. Keywords: alarm reaction; cetacean; histopathology; odontocete Introduction Free-ranging odontocete cetaceans (whales and dolphins) survive in a very demanding environment. They are creatures of the airewater interface and, as air-breathers, are in constant danger of drowning from the moment of birth. They are exposed to predation from sharks and other sea creatures, and a large proportion bear scars, often very large, from shark encounters and intra-species aggression. Even so, cetaceans have made a very successful adaptation and are found in all oceans. Hardy as they may be in that environment, most species of small odontocete whales and dolphins are very difficult to keep in captivity and are adversely affected by handling. Some beached individuals are reported to have died after simply being picked up by would-be rescuers, Correspondence to: D.F. Cowan (e-mail: [email protected]). 0021-9975/$ - see front matter doi:10.1016/j.jcpa.2007.11.009 and many dolphins have died without overt injury as a result of net encirclement in pelagic fishing operations (Cowan and Walker, 1979). Such deaths occurring in the absence of recognized injury have been attributed to ‘‘stress’’. At least some of these animals had myocardial lesions that have been attributed to catecholamine injury (Cowan and Walker, 1979; Turnbull and Cowan, 1998). Cowan et al. (1986) found numerous small myocardial scars in dolphins stranded on beaches in southern California, but could not attribute them to inflammation or coronary artery disease. Further research is required to provide a definition of the hypothetical stress injury in odontocetes. This report describes the findings of necropsy examinations of small odontocetes of many species obtained in diverse circumstances. Histopathological findings are interpreted in light of the highly specialized adaptations of cetaceans to diving and life in the marine environment. Cetaceans are protected under Ó 2008 Elsevier Ltd. All rights reserved. 25 Alarm Reaction in Odontocetes the laws of most countries, and so critical experiments to replicate lesions under controlled conditions are unlikely to ever be performed. As a result, interpretation of these findings is based on parallel studies of humans or laboratory animals. Materials and Methods Small odontocetes (n ¼ 407), primarily toothed whales and dolphins of 20 different species were collected over a period of 40 years. These animals were of both sexes and ranged in age from neonatal to 49 years as determined by examination of dentinal growth layer groups (Hohn et al., 1989). Species of dolphins and small whales examined included Atlantic bottlenose dolphin (Tursiops truncates), coastal Pacific bottlenose dolphin (Tursiops aduncus), Fraser’s dolphin (Lagenodelphis hosei), Pacific white-sided dolphin (Lagenorhynchus obliquidens), melon-headed whale (Peponocephala electra), spinner dolphin (Stenella longirostris), pantropical spotted dolphin (Stenella attenuata), striped dolphin (Stenella coeruleoalba), rough-toothed dolphin (Steno bredanensis), North Atlantic or long-finned pilot whale (Globicephala melas), short-finned pilot whale (Globicephala macrorhynchus), common dolphin (Delphinus delphis), pygmy sperm whale (Kogia breviceps), dwarf sperm whale (Kogia sima), killer whale (Orcinus orca), northern right whale dolphin (Lissodelphis borealis), Risso’s dolphin (Grampus griseus), Chinese humpback dolphin (Sousa chinensis), Baird’s beaked whale (Mesoplodon europaeus) and Dall’s porpoise (Phocoenoides dalli). Collection areas included the Trinity Bay, Newfoundland whale fishery (Cowan, 1966), eastern tropical Pacific (ETP) purse-seine tuna fishery (Cowan and Walker, 1979; Cowan and Curry, 2002), the beaches of Los Angeles and Orange counties, California (Cowan et al., 1986). Animals were also obtained from the Gulf coast of Texas, collected by the Texas Marine Mammal Stranding Network (TMMSN). These latter animals (n ¼ 129) were collected between 1990 and 2006 and were examined by one author (DFC). The interval between death and necropsy examination ranged from 20 min to 20 h. Two separate studies of by-catch in the ETP were done, one in 1979, which included 68 animals, and the second in 2000e2001 (n ¼ 56 animals). By-caught animals were examined typically within 1e2 h of death. Further contributions were received from several marine aquaria and a variety of individuals, and from the Cape Cod (Massachusetts) Stranding Network. All animals were collected according to the laws of the various countries current at the time and at the present time in the United States, under permit from the National Marine Fisheries service. No animals were killed for the purposes of this study. Autolysis is always a consideration in animals examined several hours or more after death. Autolysis was evaluated by initial assessment of the animal. Evidence of epidermal slippage, gas formation in tissues and other evidence of bacterial action including colour change and malodour were causes for rejection from this study. Autolysis was also assessed microscopically by evaluation of postmortem bacterial overgrowth, slough of vascular endothelium and pancreatic autodigestion. While only a few animals were considered perfectly preserved, all included in this study had only minor degrees of autolysis. All hearts taken from animals in the TMMSN (n ¼ 129) and the second ETP study (n ¼ 56) were serially sliced in the transverse plane. The entire heart was inspected and, in the absence of any specific grossly recognizable abnormality, a standard slice was selected for systematic sampling. The slice selected was the one that represented the level of the tip of the left anterior papillary muscle, an easily recognized standard landmark. Heart sections included the anterior left ventricle, lateral left ventricle, posterior left ventricle, anterior right ventricle, lateral right ventricle, posterior right ventricle, and septum from the standard slice, and a sample each of left and right atria. In addition, any grossly observed cardiac lesion was also sampled. Hearts from all other sources (n ¼ 222) were sampled randomly, mainly from the left ventricle, and from recognized lesions. All tissues were fixed in 10% neutral-buffered formalin at ambient temperature and processed into paraffin wax. Sections (5 mm) were stained with haematoxylin and eosin (HE) or haematoxylin, phloxine and safranin (HPS) trichrome. Results A variety of naturally occurring disease was recognized in these animals. Examples of these included parasitism of the brain and other organs (including complications such as gastrointestinal perforation), infection, lung disease, and either human-related trauma such as boat strikes or entanglement in fishing gear, or natural trauma such as caused by interspecific or intra-specific aggression. Because no differential correlations were found, except to note the great variation in these causes, naturally occurring disease will not be discussed further. Clark et al. (2006) reported adrenal changes in Atlantic bottlenose dolphins from the TMMSN collections that were deemed to be suffering from chronic stress. 26 D.F. Cowan and B.E. Curry Particular acute microscopical changes were very consistently identified in tissues from this series of animals, whatever the disease or circumstance associated with their death. These findings were widespread, involving many organs, but the major changes involved the myocardium. Frequent blotchy mottling of the heart with areas of pallor alternating with areas of congestion were interpreted to represent circulatory disturbance. Occasional subendocardial necrosis was recognized grossly (Fig. 1). Almost universal microscopical changes in myocardial tissue included hyperacidophilic fibres (Fig. 2), wavy fibres (Fig. 3), perinuclear vacuolation and contraction band necrosis (Fig. 4). These changes were widespread, but they occurred with greatest intensity in the subepicardial and subendocardial regions of the ventricles (especially the right ventricle) and affected both atria, particularly the right atrium. Similar findings were found in the fibres of the myocardial conduction system (Fig. 5). The distribution of these lesions did not appear to vary with species; however, there were species differences in the severity of these changes. Acute myocardial injury was most pronounced in Kogia, while scarring was most prominent in Delphinus. Extramural coronary arteries were uniformly normal, except for the rare occurrence of small, flat, fibrous plaques. Inflammation consistent with infectious myocarditis was observed in only one case. In addition to the range of myocardial changes described above, cellular disarray, vacuolation and contraction banding were identified in the smooth muscle of intramural coronary arteries (Fig. 6) and, in a subset of animals, eccentric plaque formation was noted in the intima of very small intramural Fig. 1. Heart of a melon-headed whale showing subendocardial pallor associated with severe ischemic injury. The right ventricle is extensively affected, but the lesion is most obvious as the yellowish subendocardial zone of the left ventricle. Fig. 2. Left ventricular myocardium from a spotted dolphin. The dark fibres are dense and hyalinized. This injury more often involves individual fibres than fascicles. Arrows indicate representative hyalinized fibres. HPS. arteries (Fig. 7). In addition to these features, hyalinization and disarray of arterial smooth muscle were present and these changes were interpreted to suggest prolonged spasm. Fibrous plaques affecting the intima of small intramural coronary arteries were commonly observed and these were interpreted as being a sequela to arterial thrombosis. One animal had evidence of recanalization of a thrombus (Fig. 8). Many animals had evidence of ventricular myocardial scarring, and this lesion was particularly noted in common dolphins stranded on beaches in southern California (Cowan et al., 1986). These regions of scarring appeared as small, superficial indentations and as stellate myocardial scars. The ventricular lesions were sometimes accompanied by focal or diffuse atrial Fig. 3. Left ventricular myocardium from a spotted dolphin. The dark staining, stretched, compressed wavy fibres also have condensed nuclei. HPS. Alarm Reaction in Odontocetes Fig. 4. Right ventricular myocardium from a striped dolphin showing prominent contraction banding. Arrows indicate representative bands which may be narrow or broad. HPS. 27 Fig. 6. Intramural coronary artery within the left ventricle of a striped dolphin. There is disarray, spastic contraction and vacuolation of mural myocytes in the vessel in the centre of the image. HPS. fibrosis, and one 27-year-old female bottlenose dolphin from Texas had almost complete replacement of the right atrium with fibrous scar in addition to atrial thrombosis. Small patchy fibrous scars in the myocardium (ventricular or atrial), of a size and in locations consistent with myocardial necrosis, as described above (Fig. 9), were common (e.g. these were present in 20 of 56 animals in the second ETP study). Most of these scars were small (1e2 mm), but some were evident on gross inspection. Contraction banding of smooth muscle of the viscera (intestine, urinary bladder) and of the media of vessels of many organs was present in every animal (Fig. 10). In the viscera, smooth muscle banding was circumferential, and occasionally produced thickening of segments of the muscularis mucosa. Sloughing of the superficial mucosa of the intestine was present in every case, even in animals examined less than 1 h after death. This was most pronounced in, but not limited to, the small intestine. Marked contraction of the bronchial sphincter muscles and pulmonary air entrapment was an almost universal change (Fig. 11). This change appeared to result in inability of the elastic lung tissue to collapse when the thorax was opened and, in life, probably resulted in impaired ventilation. Acute necrosis of segments of the renal tubules (acute tubular necrosis; ATN), commonly with infraglomerular reflux (Fig. 12), was very common finding, present in nearly Fig. 5. Right ventricular subendocardium from a bottlenose dolphin showing contraction banding in prominent Purkinje fibres (arrows). Bands are broader than in the myocardium. HPS. Fig. 7. Intramural coronary artery within the left ventricle of a common dolphin. There is a subocclusive eccentric fibromuscular plaque, attributed to organized thrombosis. HPS. 28 D.F. Cowan and B.E. Curry Fig. 8. Vascular webs (arrows) in a small intramural artery within the left ventricle of a bottlenose dolphin. HPS. all animals. Residue of smooth muscle injury (scarring) in the viscera was not identified. Discussion A single definition of ‘‘stress’’ has not been generally accepted by workers in the field of stress research (Levine, 1985; Moberg, 1987; Chrousos et al., 1988; Levine and Ursin, 1991; Fowler, 1995). Here, we define the term stress as demand for adaptation with the recognition that organisms are in a constant condition of adjustment of physiological systems to maintain homeostasis. Stress is then specified in terms of the demand (‘‘stressor’’) applied. Examples of stressors include heat stress, cold stress, crowding stress and social interaction. In cetaceans, stress also occurs in the adaptations needed to adjust to diving, Fig. 9. Left ventricular myocardium from a bottlenose dolphin showing irregular patchy fibrous scars and scattered hyalinized fibres. Note the absence of an inflammatory infiltrate. HPS. Fig. 10. Small intestinal muscularis propria from a spotted dolphin. There is marked contraction banding, seen here as ‘‘tiger striping’’ typically involving groups of fibres. HPS. for flight from predators or from intraspecific aggression, or to catch prey. Stress may occur whenever a wild animal is restrained or enclosed, however loose the restraint or enclosure might seem to the captor. Apprehension may be a mild psychological stressor that may intensify to become anxiety, fright or even terror (Fowler, 1995). Thus, the issue is not whether cetaceans are ‘‘under stress’’ in a given situation, as they are always under demand for adaptation. The issue is whether the degree and duration of stress experienced is physiologically damaging or not. The effects of stress are expected to vary with the adaptive mechanisms of the subject, and will therefore vary with the species and its environment. Since stress may have a psychological component, which could be influenced by experience, it may be expected Fig. 11. Lung from a spotted dolphin showing spasm of bronchiolar sphincters with air-trapping. S, sphincter muscle; L, bronchial lumen; C, bronchial cartilage. HPS. Alarm Reaction in Odontocetes Fig. 12. Acute renal tubular necrosis in a bottlenose dolphin. There is reflux of the detached proximal tubular epithelium into Bowman’s space of the glomerulus. G, glomerulus; TE, refluxed tubular epithelium. Arrows indicate Bowman’s capsule. HPS. to vary among individuals of the same species. Stress is not imposed, but is an expression of a response to a challenge, or stressor. Unfamiliar or extreme challenges may evoke unusual or extreme responses. That is, damaging stress might result from inappropriate or extreme adaptive responses. Syncope, for example, a transient dysfunction of the autonomic nervous system, has no particular adaptive value, but it does occur in some people under stress. In an extreme situation, some animals undergo an extreme, fatal cholinergic bradycardia (Fowler, 1995). All stressors do not elicit the same physiological or behavioural responses in all animal species. There are different types and degrees of stress, and these factors can influence an individual’s response to stress. Acute stress is sudden, whereas chronic stress is marked by long duration or frequent occurrence. An acute stressor may elicit an adaptive physiological response, but chronic stress may be pathogenic, resulting in tissue changes (Clark et al., 2006). The effects of both can be cumulative over time (Chrousos and Gold, 1992). Cetaceans exhibit the basic mammalian response to stress (Curry, 1999). Chronic stress or repeated acute stress can have maladaptive effects on immune responses, reproductive function and growth (Moberg, 1987, 1991; Rivier and Rivest, 1991; Chrousos, 1992; Chrousos and Gold, 1992; Chrousos, 1995; McEwen et al., 1997). Well-documented physiological responses to stressors in odontocetes include adrenocortical responses and effects on thyroid hormone balance. Elevated blood cortisol levels have been observed in odontocetes subjected to stressors such as capture, handling and restraint, although the elevations 29 appear to be modest in comparison with those known for other mammals experiencing similar stressors (Thomson and Geraci, 1986; St Aubin and Geraci, 1990). In contrast, aldosterone, which does not typically characterize the adrenal response to stress in terrestrial mammals, is greatly increased in cetaceans (and pinnipeds) subjected to adrenocortical stimulation (Thomson and Geraci, 1986; St Aubin and Geraci, 1990). This may be an adaptation reflecting the need to enhance water and sodium reabsorption during stress (St Aubin and Geraci, 1990; St Aubin et al., 1996). Clark et al. (2006) have demonstrated changes in the adrenal medulla of bottlenose dolphins under chronic stress. Thyroid hormone balance in odontocetes also appears to be sensitive to stress (Ridgway and Patton, 1971; St Aubin and Geraci, 1988, 1992; Orlov et al., 1988; St Aubin et al., 1996). Cowan and Walker (1979) in a study of by-catch from the ETP fishery concluded that several dolphins apparently died of massive cardiac reaction to stress, finding cardiac lesions consistent with those produced in laboratory animals injected with catecholamine and humans thought to have died of stress cardiomyopathy (Cebelin and Hirsch, 1980) The physiological response of marine mammals to exercise is quite different from the mammalian exercise response. When terrestrial mammals, who have unlimited access to atmospheric oxygen, undertake exercise, ventilation rate and cardiac output increase and peripheral vasodilation increases skeletal muscle perfusion and the dissipation of heat through the skin or lungs by panting. In contrast, diving marine mammals breath-hold, the heart rate slows and peripheral vessels constrict, the degree of bradycardia and peripheral vasoconstriction reflecting the level of exertion. Peripheral vasoconstriction maintains central arterial pressure despite decreased cardiac output by reducing flow to all organs and tissues except the brain, and all organs and tissues, including the heart, kidneys and splanchnic organs, experience a reduction in convective oxygen delivery resulting from both hypoxic hypoxia and ischemic hypoxia (Butler and Jones, 1997; Davis and Kanatous, 1999; Kanatous et al., 1999). An odontocete in free dive (voluntary dive) undergoes certain physiological adjustments reflecting exercise, including reflexive apnoea, with voluntary override, minimal cardiovascular adjustments, and a general maintenance of aerobic metabolism (Butler and Jones, 1997). Blood flow is reduced to the gut and kidneys, but maintained in the heart, brain and, to a degree, exercising muscles. The animal is able to surface and dive repeatedly (foraging dive pattern), as there is little lactic acid build-up. These features have together been termed the ‘‘dive response’’. An 30 D.F. Cowan and B.E. Curry odontocete in an involuntary dive situation undergoes a somewhat different set of adjustments, which have been termed the ‘‘dive reflex’’, but which may better be termed an ‘‘alarm reaction’’. These adjustments include not only reflexive apnoea, but also decreased heart rate (diving bradycardia), reduction of cardiac output and vasoconstriction with markedly decreased perfusion of gut, liver, kidneys and skeletal muscle and a substantial increase in production of lactic acid in these tissues, which is reflected in marked rise in blood levels on surfacing (Butler, 1982; Ridgway, 1986). The clear implication of the distinctive reactions to voluntary and involuntary diving is that the odontocete is responding to the environment as it is perceived; the triggering of the alarm reaction is a reaction to a situation interpreted by the animal as a dire threat, and the response involves a marked autonomic reaction. Since the major threats to an aquatic, air breathing mammal are drowning and predation, the alarm reaction is an accentuation of the physiological dive and escape responses. In responding to a perceived threat, odontocetes will mobilize for fight or flight, according to the established concept of stress. In terrestrial mammals, flight means acceleration of muscular activity, elevation of blood pressure, tachycardia and hyperventilation. For a diving mammal, however, mobilization for flight means breath-holding and re-directing the flow of blood away from non-vital to vital oxygendependent organs (i.e. the brain and heart). This is physiological and non-injurious, provided the changes are coherent and not extreme or overly protracted. Our hypothesis is that in the instance of a novel threat perceived as extreme, the smooth coordination of the cardiovascular adjustments may break down and a massive release of adrenergic hormone from the adrenal medulla occurs, a phenomenon well recognized in man and other mammals (Eliot et al., 1977). In this instance, a ‘‘sympathetic storm’’ occurs, with spasm of small intramural coronary arteries and myocardial ischemia, as occurs in human drowning (Lunt and Rose, 1987) This ischemia may be associated with arrhythmia and death of the animal. It may result in patchy death of myocytes followed by scarring in survivors, or it may occur with no evidence of residual injury. Muscle cells can respond to injury in limited fashion, so specific changes may have any of several different causes. ‘‘Wavy fibre’’ refers to a change in the appearance of a myocardial fibre in which it becomes thin and attenuated, dense or hyalinized, and assumes an angulated or ‘‘wavy’’ accordion-pleated appearance as if it is longer than the space in which it is contained, and is longitudinally compressed until it buckles. Wavy fibres are a characteristic sign of acute myocardial ischemia (Bouchardy and Majno, 1971, 1974; Eichbaum, 1975). Waving is thought to occur when a fibre becomes ischemic or suffers acute adrenergic injury with impairment of electrogenesis; that is, the fibre, while living, becomes unable to contract. Stretching forces in diastole or stretching due to bulging of the myocardial wall stretches or attenuates the fibres, which because unable to contract and become longitudinally compressed as surrounding muscle fibres or connective tissue elements shorten. The overextended fibre is unable to contract to a normal length relative to the fibres or tissue around it (Eichbaum, 1975). Ischemic wavy fibres are not necessarily necrotic, although they may well become so. Contraction banding is a manifestation of disruptive spastic contraction of contractile elements in a myocyte and in conduction fibres, producing a characteristic coarse cross-banding of the affected fibre (Hansen, 1995). The lesions of the myocardium described in the present report are all recognized to occur in man, monkeys, rats and pigs. All are attributable to catecholamine injury (Reichenbach and Benditt, 1970; Cebelin and Hirsch, 1980; Turnbull and Cowan, 1998), or to ischemia (Mukherjee et al., 1982; Muntz et al., 1984). Indeed, the final effector in ischemia may be the local release of catecholamines from the nerves in the heart (Mukherjee et al., 1982). The lesions of the intramural coronary vessels suggest spasm, which can produce ischemic injury with or without necrosis (Maseri and Chierchia, 1982), or by reperfusion, that is, interruption of blood flow followed after an interval by re-establishment of flow (Hansen, 1995). The patchy discoloration of the hearts observed on gross examination strongly suggests alternating areas of congestion and ischemia, which may be explained by irregular perfusion. Healing of injured striated muscle, including myocardium, may occur without scarring provided that the fine structure of the tissue, including basement membranes, is intact and if nuclei survive the injury. Therefore, the absence of recognizable scarring does not preclude prior myocardial necrosis. Scars indicate injury in foci too large to be amenable to restoration (Carlson, 1973). It is worth emphasizing that myocardial injury related to ‘‘stress’’ is not limited to odontocetes, but can occur in many species, under both stringent artificial laboratory conditions and under natural conditions (Van Vleet and Ferrans, 1986). Almost all animals in this study had lesions of acute renal tubular necrosis. This lesion is most commonly caused by prolonged renal ischemia (Kaufman et al., 1991; Kashgarian, 1998). Infraglomerular tubular reflux is the movement or intrusion of detached 31 Alarm Reaction in Odontocetes proximal tubular epithelium into Bowman’s space, around the renal glomerulus. This change is considered a very sensitive indicator of tubular epithelial damage (Waugh et al., 1964; Kashgarian, 1998). ATN, as seen in odontocetes in the ETP studies, is consistent with the histological observations of tissues from mammals suffering from capture myopathy (Curry, 1999). The distal airways of cetaceans are different from those of terrestrial animals in that the walls are made rigid with cartilage bars to the mouth of the alveolar duct, and the airways are equipped with a series of muscular sphincters. Many of the odontocetes in the present study showed expansion or overexpansion of alveolar spaces, when the normal elasticity of the lung should have caused collapse on opening the chest cavities. This was attributed to a readily observable spasm of the sphincter muscles of the distal airways. These smooth muscle structures also showed overcontraction, very similar to the overcontraction of the small intramural coronary arteries. We are inclined to attribute this highly stereotyped multisystem pattern of smooth muscle spasm of arterial media, bronchial sphincters and visceral smooth muscle to a single cause, massive protracted autonomic discharge, a ‘‘sympathetic storm’’ that is accompanied by high levels of circulating catecholamines. This discharge occurs as the major manifestation of an extreme stress response in odontocetes. Capture myopathy, a condition resulting from muscle exertion associated with capture and restraint of wildlife, is characterized by a variable and lengthy list of clinical signs, including ataxia, paralysis, myoglobinuria and acute muscle degeneration (Harthoorn and Young, 1974; Bartsch et al., 1977; Chalmers and Barrett, 1977; Basson and Hofmeyr, 1978; Hulland, 1985). Capture myopathy can be induced by a combination of many stressors (e.g. terror, chase, capture or restraint), and is associated with exhaustion of the normal physiological reserves that provide energy for escape. ATN and severe glomerular damage have been identified in many animals dying with capture myopathy, and are considered to be the result of renal hypoxia caused by catecholamine activity related to shock or the sympathetic storm described above (Wallace et al., 1987; Spraker, 1993; Williams and Thorne, 1996). Myoglobinuric casts that could result from myopathy were not found in the renal tubules of cetaceans in the present study. While the lesions described in odontocetes in the present study have certain features in common with capture myopathy (myocardial lesions, ATN), systematic muscle sampling was not part of the necropsy protocol, and we cannot assert that the alarm reaction in cetaceans is a form of capture myopathy at this time. The present study of many species of beachstranded, net-caught and captive odontocetes has revealed similar patterns of pathological change in organs and tissues. These changes are consistent with multi-systemic injury caused by massive release of endogenous catecholamines (alarm reaction) or by vasospasm, including spasm of small cardiac arteries, with ischemia and reperfusion. This pattern of pathology includes contraction band necrosis of cardiac and smooth muscles, ischemic injury to the intestinal mucosa, acute renal tubular necrosis and distal bronchospasm. The pattern appears to result from a stereotypic stress response, independent of the nature of the provoking stimulus. It may explain the propensity of otherwise hardy animals to die in an otherwise non-damaging stressful situation. This pattern of injury is rooted in the physiological adaptations of cetaceans that are associated with a fully aquatic life. Histopathological findings suggest that the reflexive response of an odontocete to any major perceived threat, the alarm reaction, is to activate all of the physiological adaptations to diving or escape to an extreme or pathological level, resulting, if greatly prolonged, in widespread ischemic injury to tissues. These observations may explain why ‘‘sensitive’’ species die abruptly from handling or transportation, and why the mortality of highly stressed beach-stranded animals is very high. Acknowledgments Thanks to Graham Worthy, past Director of TMMSN, the volunteers of the Texas Marine Mammal Stranding Network for their enthusiastic participation, and many other contributing colleagues. This work was supported by grants from the National Oceanic and Atmospheric Administration (NOAA) through the National Sea Grant College Program (NA16RGO457-01) and the Environmental Protection Agency (EPA) Gulf of Mexico Program (MX822147-01-0). The views expressed herein are those of the authors and do not necessarily reflect the views of NOAA, the EPA or any of their subagencies. References Bartsch, R. C., McConnell, E. E., Imes, G. D. and Schmidt, J. M. (1977). A review of exertional rhabdomyolysis in wild and domestic animals and man. Veterinary Pathology, 14, 314e324. Basson, P. A. and Hofmeyr, J. M. (1978). Mortalities associated with wildlife capture operations. In: The Capture and Care of Wild Animals, E. Young, Ed., Human and Rousseau, Cape Town, pp. 151e160. 32 D.F. Cowan and B.E. Curry Bouchardy, B. and Majno, G. (1971). A new approach to the histologic diagnosis of myocardial infarcts. Cardiology, 56, 327e332. Bouchardy, B. and Majno, G. (1974). Histopathology of early myocardial infarcts. American Journal of Pathology, 74, 301e318. Butler, P. J. (1982). Respiratory and cardiovascular control during diving in birds and mammals. Journal of Experimental Biology, 100, 195e221. Butler, P. J. and Jones, D. R. (1997). Physiology of diving birds and mammals. Physiological Reviews, 77, 837e899. Carlson, B. M. (1973). The regeneration of skeletal muscle e a review. American Journal of Anatomy, 137, 119e149. Cebelin, M. S. and Hirsch, C. S. (1980). Human stress cardiomyopathy: myocardial lesions in victims of homicidal assaults without internal injuries. Human Pathology, 11, 123e132. Chalmers, G. A. and Barrett, W. (1977). Capture myopathy in pronghorns in Alberta, Canada. Journal of the American Veterinary Medical Association, 171, 918e923. Chrousos, G. P. (1992). Regulation and dysregulation of the hypothalamicepituitaryeadrenal axis. Neuroendocrinology, 21, 833e858. Chrousos, G. P. (1995). The hypothalamicepituitarye adrenal axis and immune-mediated inflammation. New England Journal of Medicine, 332, 1351e1362. Chrousos, G. P. and Gold, P. W. (1992). The concept of stress and stress system disorders. Journal of the American Medical Association, 267, 1244e1252. Chrousos, G. P., Loriaux, L. D. and Gold, P. W. (1988). The concept of stress and its historical development. In: Mechanisms of Physical and Emotional Stress, vol. 245, G. P. Ghrousos, L. D. Loriaux and P. W. Gold, Eds, Plenum Press, New York, pp. 3e7. Clark, L. S., Cowan, D. F. and Pfeiffer, D. C. (2006). Morphological changes in the Atlantic bottlenose dolphin (Tursiops truncatus) adrenal gland associated with chronic stress. Journal of Comparative Pathology, 135, 208e216. Cowan, D. F. (1966). Pathology of the pilot whale, Globicephala melaena. A comparative survey. Archives of Pathology, 82, 178e189. Cowan, D. F. and Curry, B. E. (2002). Histopathological Assessment of Dolphins Necropsied Onboard Vessels in the Eastern Tropical Pacific Tuna Fishery. Southwest Fisheries Science Center, National Marine Fisheries Service, NOAA, La Jolla, California. Administrative Report LJ-02-24c. Cowan, D. F. and Walker, W. A. (1979). Disease Factors in Stenella attenuata and Stenella longirostris Taken in the Eastern Tropical Pacific Yellowfin Tuna Purse Seine Fishery. Southwest Fisheries Science Center, National Marine Fisheries Service, NOAA, La Jolla California. Administrative Report No. LJ-79-32c. Cowan, D. F., Walker, W. A. and Brownell, R. L., Jr. (1986). Pathology of small cetaceans stranded along southern California beaches. In: Research on Dolphins, M. M. Bryden and R. Harrison, Eds, Oxford University Press, Oxford, pp. 323e367. Curry, B. E. (1999). Stress in mammals: the potential influence of fishery-induced stress on dolphins in the eastern tropical Pacific Ocean. NOAA Technical Memorandum. NOAA-TM-NMFS-SWFSC-260. Davis, R. W. and Kanatous, S. B. (1999). Convective oxygen transport and tissue oxygen consumption in Weddell seals during aerobic dives. Journal of Experimental Biology, 202, 1091e1113. Eichbaum, F. W. (1975). Wavy myocardial fibers in spontaneous and experimental adrenergic cardiopathies. Cardiology, 60, 358e365. Eliot, R. S., Clayton, F. C., Pieper, G. M. and Todd, G. L. (1977). Influence of environmental stress on pathogenesis of sudden cardiac death. Federation Proceedings, 36, 1719e1724. Fowler, M. E. (1995). Restraint and Handling of Wild and Domestic Animals, 2nd Edit., Iowa State University Press, Ames. Hansen, P. R. (1995). Myocardial reperfusion injury: experimental evidence and clinical relevance. European Heart Journal, 16, 734e740. Harthoorn, A. M. and Young, E. (1974). A relationship between acidebase balance and capture myopathy in zebra (Equus burchelli) and an apparent therapy. Veterinary Record, 95, 337e342. Hohn, A. A., Scott, M. D., Welles, R. S., Sweeney, J. S. and Irvine, A. B. (1989). Growth layers in teeth from known age free-ranging bottlenose dolphins. Marine Mammal Science, 5, 315e342. Hulland, T. J. (1985). Muscles and tendons. In: Pathology of Domestic Animals, vol. 1, K. V. F. Jubb, P. C. Kennedy and N. Palmer, Eds, Academic Press, New York, pp. 139e199. Kanatous, S. B., Davis, R. W. and Cowan, D. F. (1999). High aerobic capacities in the skeletal muscles of seals, sea lions and fur seals: an adaptation to diving hypoxia? Journal of Applied Physiology, 86, 1247e1256. Kashgarian, M. (1998). Acute tubular necrosis and ischemic renal injury. In: Heptinstall’s Pathology of the Kidney, 5th Edit., J. C. Jennette, J. L. Olson, M. M. Schwartz and F. G. Silva, Eds, Lippincott-Raven, Philadelphia. p. 872. Kaufman, J., Dhakal, M., Patel, B. and Hamburger, R. (1991). Community acquired acute renal failure. American Journal of Kidney Diseases, 17, 191e198. Levine, S. (1985). A definition of stress? In: Animal Stress, G. P. Moberg, Ed., American Physiological Society, Bethesda, pp. 27e48. Levine, S. and Ursin, H. (1991). What is stress? In: Stress: Neurobiology and Neuroendocrinology, M. R. Brown, G. F. Koob and C. Rivier, Eds, Marcel Dekker, New York, pp. 3e22. Lunt, D. W. and Rose, A. G. (1987). Pathology of the human heart in drowning. Archives of Pathology and Laboratory Medicine, 111, 939e942. McEwen, B. S., Biron, C. A., Brunson, K. W., Bulloch, K., Chambers, W. H., Dhabhar, F. S., Goldfarb, R. H., Kitson, R. P., Miller, A. H., Spencer, R. L. and Weiss, J. M. (1997). The role of adrenocorticoids as modulators of immune function in health and disease: Alarm Reaction in Odontocetes neural, endocrine and immune interactions. Brain Research Reviews, 23, 79e133. Maseri, A. and Chierchia, S. (1982). Coronary artery spasm: demonstration, definition, diagnosis and consequences. Progress in Cardiovascular Diseases, 25, 169e192. Moberg, G. P. (1987). Problems in defining stress and distress in animals. Journal of the American Veterinary Medical Association, 191, 1207e1211. Moberg, G. P. (1991). How behavioral stress disrupts the endocrine control of reproduction in domestic animals. Journal of Dairy Science, 74, 304e311. Mukherjee, A., Bush, L. R., McCoy, K. E., Duke, R. J., Hagler, H., Buja, L. M. and Willerson, J. T. (1982). Relationship between beta adrenergic receptor numbers and physiological responses during experimental canine myocardial ischemia. Circulation Research, 50, 735e741. Muntz, K. H., Boulas, H. K., Willerson, J. F. and Buja, L. M. (1984). Redistribution of catecholamines in the ischemic zone of the dog heart. American Journal of Pathology, 114, 64e78. Orlov, M. V., Mukhlya, A. M. and Kulikov, N. A. (1988). Hormonal indices in the bottle-nosed dolphin in the norm and in the dynamics of experimental stress. Zhurnal Évolyutsionnoi Biokhimii I Fiziologii, 24, 431e436. Reichenbach, D. D. and Benditt, E. P. (1970). Catecholamines and cardiomyopathy: the pathogenesis and potential importance of myofibrillar degeneration. Human Pathology, 1, 125e150. Ridgway, S. H. (1986). Diving by cetaceans. In: Diving in Animals and Man, A. O. Brubakk, J. W. Kanwisher and G. Sundness, Eds, The Royal Norwegian Society of Science and Letters, Trondheim, Norway, pp. 33e62. Ridgway, S. H. and Patton, G. S. (1971). Dolphin thyroid: some anatomical and physiological findings. Zeitschrift fur Vergleichende Physiologie, 71, 129e141. Rivier, C. and Rivest, S. (1991). Effect of stress on the activity of the hypothalamicepituitaryegonadal axis: peripheral and central mechanisms. Biology of Reproduction, 45, 523e532. St Aubin, D. J. and Geraci, J. R. (1988). Capture and handling stress suppresses circulating levels of thyroxine (T4) and triiodothyronine (T3) in beluga whales Delphinapterus leucas. Physiological Zoology, 61, 170e175. St Aubin, D. J. and Geraci, J. R. (1990). Adrenal responsiveness to stimulation by adrenocorticotropic hormone 33 (ACTH) in captive beluga whales, Delphinapterus leucas. In: Advances in Research on the Beluga Whale, Delphinapterus leucas, vol. 224, T. G. Smith, D. J. St Aubin and J. R. Geraci, Eds, Canadian Journal of Fisheries and Aquatic Sciences, 149e157. St Aubin, D. J. and Geraci, J. R. (1992). Thyroid hormone balance in beluga whales, Delphinapterus leucas: dynamics after capture and influence of thyrotopin. Canadian Journal of Veterinary Research, 56, 1e5. St Aubin, D. J., Ridgway, S. H., Wells, R. S. and Rhinehart, H. (1996). Dolphin thyroid and adrenal hormones: circulating levels in wild and semi-domesticated Tursiops truncatus, and influence of sex, age, and season. Marine Mammal Science, 12, 1e13. Spraker, T. R. (1993). Stress and capture myopathy in Artiodactylids. In: Zoo and Wild Animal Medicine, M. E. Fowler, Ed., W.B. Saunders, Philadelphia, pp. 481e488. Thomson, C. A. and Geraci, J. R. (1986). Cortisol, aldosterone, and leucocytes in the stress response of bottlenose dolphins, Tursiops truncatus. Canadian Journal of Fisheries and Aquatic Sciences, 43, 1010e1016. Turnbull, B. S. and Cowan, D. F. (1998). Myocardial contraction band necrosis in stranded cetaceans. Journal of Comparative Pathology, 118, 317e327. Van Vleet, J. F. and Ferrans, J. V. (1986). Myocardial diseases of animals. American Journal of Pathology, 124, 97e178. Wallace, R. S., Bush, M. and Montali, R. J. (1987). Deaths from exertional myopathy at the National Zoological Park from 1975 to 1985. Journal of Wildlife Diseases, 23, 454e462. Waugh, D., Schleiter, W. and James, A. (1964). Infraglomerular epithelial reflux: an early lesion of acute renal failure. Archives of Pathology, 77, 93e96. Williams, E. S. and Thorne, E. T. (1996). Exertional myopathy (capture myopathy). In: Noninfectious Diseases of Wildlife, 2nd Edit., A. Fairbrother, L. N. Locke and G. L. Hoff, Eds, Iowa State University Press, Ames, pp. 181e193. April 12th, 2007 ½ Received, Accepted, November 22nd, 2007