Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
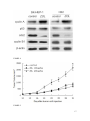
Zoledronic acid inhibits growth of hepatocellular carcinoma cells in vitro and in vivo 1 1 Quan LIU, 2Yonghui TAO, 1Ruizhen BAI, 1Shujian CHANG, 1*Dong HUA Oncology institute, The Fourth Affiliated Hospital of Soochow University, Wuxi 214062, China 2 Key Laboratory on Technology for Parasitic Disease Prevention and Control, Ministry of Health, Jiangsu Institute of Parasitic Diseases, Wuxi, Jiangsu, People's Republic of China. * Correspondence to: Dr. Dong Hua (email: [email protected] ), Oncology institute, The Fourth Affiliated Hospital of Suzhou University, Wuxi 214062, China Tel:86-510-88682109 The first two authors contributed equally to this work. 1 Abstract Background Growing preclinical evidence shows that zoledronic acid (ZOL) exhibits direct antitumor activity in various cancer cell lines. However, the cytotoxic effects of ZOL on human hepatocellular carcinoma (HCC) cells have not been established. In the present study, we investigated the effect of ZOL on HCC both in vitro and in vivo. Methods Cytotoxicity and cell cycles were assessed with Sulforhodamine B colorimetric assay and flow cytometry. Expression levels of cell cycle phase-linked proteins were examined. The effect of ZOL on HCC in vivo was explored based on H22- s.c. (subcutaneous injection) and H22- i.p. (intraperitoneal injection) mice model. Results ZOL inhibited the growth of SK-HEP-1 and H22 cells and induced S-phase arrest through downregulating cdc2 protein and upregulating cyclin A. It inhibited the growth of s.c tumors, and increased the survival of both H22- s.c. and H22- i.p. mice in vivo. Conclusion In conclusion, ZOL inhibits growth of HCC cells in vitro and in vivo. Additional studies are warranted for management of HCC based on ZOL. Keywords Zoledronic acid; Hepatocellular carcinoma; S phase arrest 2 BACKGROUND Hepatocellular carcinoma (HCC) is the third leading cause of cancer-related mortality worldwide, with an estimated one million new cases each year [1]. Most HCC cases are diagnosed in the late stage, and the median survival following diagnosis is only approximately 6 to 20 months [2]. Only 40% of HCC patients are eligible for potential curative treatments (resection, transplantation, or local ablation). 20% of the patients are eligible for chemoembolization, and for the remaining 40% patients with unresectble lesions and unsuitable for locoregional therapy, the systemic therapy is appropriate [3]. The effect of chemoembolization and systemic therapy relies heavily on the use of chemical drugs. Unfortunately, no specifically effective drug has been identified for the HCC patients to date. This situation necessitates discovery and development of new drugs to improve the therapeutic effect among HCC patients. Zoledronic acid (ZOL) is heterocyclic imidazole bisphosphonate a used third-generation as standard nitrogen-containing care for preventing skeletal-related events (SREs) in patients with bone metastases [4]. Recently, growing preclinical evidence shows that ZOL exhibits remarkable antitumor activity in several cancer cell lines [5-8] and animal models [9-12]. The antitumor ability has also been corroborated in clinical trials. In postmenopausal breast cancer patients, ZOL combined with standard adjuvant therapy reduces the risk of mortality by 29% (hazard ratio, 0.71; p=0.017) compared with the adjuvant therapy alone [13]. Consequently, ZOL may exert as a promising therapeutic effect in multiple cancers. However, the cytotoxic effects of 3 ZOL on HCC have not been established so far. In the present study, we investigated impact of ZOL on HCC both in vitro and in vivo. METHODS Cell culture The SK-HEP-1 cell line was obtained from the American Type Culture Collection (Manassas, Virginia, US). H22 cell lines were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). All cancer cell lines were maintained with DMEM medium. The medium was supplemented with 10% inactivated fetal bovine serum (Life Technologies, Carlsbad, California, US) and penicillin/streptomycin (200 μM/l). Cultures were kept in an incubator at 37.0 ℃ in a water-saturated atmosphere with 5% CO2. Sulforhodamine B colorimetric assay The viability of SK-HEP-1 and H22 cells was evaluated by Sulforhodamine B colorimetric assay (SRB) ( St. Louis, Missouri, US). 5000 cells/well were seeded into 96-well plates and left to adhere overnight. The cells were incubated for 48 h in the absence or presence of 0.5-350 μM ZOL(Novartis Pharma AG, Basel, Switzerland). Cell monolayers were fixed with 10% (wt/vol) trichloroacetic acid and stained for 30 min, and after that the excess dye was removed by washing repeatedly with 1% (vol/vol) acetic acid. The protein-bound dye was dissolved in 10 mM Tris base solution for optical density determination at 495 nm using a microplate reader. The half-maximal maximal inhibitory concentration (IC50) was determined using the nonlinear regression program CalcuSyn (Biosoft, Cambridge, UK). 4 Flow cytometry Cells were incubated with or without ZOL as indicated and harvested. The cells were fixed with 70% ethanol at 4 °C overnight. The fixed cells were washed with D-PBS and resuspended in phosphate-buffered saline containing 500 μg/ml RNase A and 50 μg/ml propidium iodide (Sigma-Aldrich, St. Louis, Missouri, US). DNA content was determined by flow cytometry using a Coulter EPICS XL-MCL cytometer (Coulter Co., Miami, Florida, USA) and the cell cycle phase distribution was analyzed by ModFit 3.0 software (Verity Software House, Topsham, Maine, USA). Western blotting Total proteins were extracted by lysing cells with a buffer containing 50 mM Tris–HCl, pH 7.4, 150 mM NaCl, 5 mM MgCl2, 0.5 mM EDTA, 0.1% SDS and protease inhibitor cocktail followed by ultracentrifugation at 13,000 rpm for 10 min. All fractions were assayed for protein contents according to the protocol [14]. Equal amounts of proteins were mixed with Laemmli sample buffer, resolved on SDS/PAGE (12%) and blotted onto PVDF membranes (Millipore, Billerica, Massachusetts). Immunodetection was carried out in 50 mM Tris–HCl, pH 7.4, 150 mM NaCl, 0.1% Tween 20, and 5% non-fat dry milk, followed by HRP-conjugated secondary antibodies at 1:20,000 dilution. Detection was performed using an ECL kit. H22 Solid tumor-bearing mouse model 5 In vivo experiments were carried out in compliance with Guidelines on the Humane Treatment of Lab Animals (Agreement No. [2006]398 ). H22 was maintained in DMEM medium. H22 cells (1x107 ) were inoculated subcutaneously (sc.) into the right flanks of 4-6-week- Female Kunming mice (SLAC Laboratory animal, Shanghai). 30 of the inoculated mice developed palpable tumors (60–90 mm3) by Day 10 after tumor cell injection. These mice were randomly assigned to various treatment groups (control, treated by ZOL 100μg/kg group, ZOL 400μg/kg group). ZOL was injected into the mouse peritoneal cavity 100μg/kg or 400μg/kg every other day for a total of four injections. PBS was used as a control. Tumors were measured using calipers. Tumor size was measured with calipers every 3 days and tumor volumes were calculated (tumor volume = 0.52 × length × width2). Mice were sacrificed when tumors reached 2,000 mm3. Ascites tumor-bearing mouse model Four- to six-week female Kunming mice were inoculated intraperitoneally (i.p.) For 1x107 H22 cell suspension on Day 0. After 3 days of tumor transplantation, ZOL was administrated to the peritoneal cavity of H22-bearing mice at a dose of 100 μg/kg body weight every other day for a total of four injections, and the survival days of mice were recorded. Statistical analysis Results were presented as means ± SD. Statistical significance was determined by one-way analysis of variance followed by the Student’s t test. Kaplan-Meier survival 6 curves were determined by the Log-rank test. A value of P<0.05 was considered statistically significant. RESULTS Antiproliferative activity assay SK-HEP-1 and H22 cell lines were treated with increasing concentrations of ZOL for 48 h. The growth inhibitory effects of cells were examined by SRB assay. As shown in Figure 1, after 48 h of ZOL treatment SK-HEP-1 and H22 showed significantly reduced growth in a dose-dependent manner. S phase arrest in HCC cells. To elucidate the effect of ZOL on the cell cycle progression, flow cytometry analysis of DNA content was performed on SK-HEP-1 and H22 cells (Figure 2). SK-HEP-1 and H22 cells were treated with different concentrations of ZOL. A clear cell cycle perturbation effect of ZOL was observed. At 48 h after the treatment, with increased concentrations of ZOL, the cells accumulated in the S phase, and gradually decreased in G1 and G2/M phases (Figure 2). We speculated that ZOL arrested SK-HEP-1 and H22 cells in S phase. Expression of phase markers and cell cycle regulators In order to understand whether ZOL induced an S-phase cell cycle arrest in SK-HEP-1 and H22 cell lines, we investigated the expression of p53, cyclin B1, cyclin A and cdc2 in SK-HEP-1 and H22 cell lines treated by ZOL. Cells were treated with 0.3% DMSO or 20 μM ZOL, and analyzed for protein expression levels after 48 h of treatment (Figure 3). 7 There was no significant change in the expression of Cyclin B1 and p53. Cdc2 was markedly decreased after treatment with ZOL, while cyclin A was upregulated. Zol inhibited tumor growth in vivo. To evaluate the antitumor activity of the ZOL in vivo, xenograft H22 liver cancer model was established in Kunming mice. Tumor volumes were recorded to assess the antitumor effect in these treatment groups. As shown in Figure 4, the tumors in the PBS group grew progressively during the course of the experiment. On the contrary, a lag was noticed in the growth of tumor in the ZOL group until Day 19. At the sixth week after administration, the average tumor volume of ZOL-treated mice was statistically smaller than that of untreated mice (P < 0.05). Survival of H22-s.c. and H22-i.p. mice To determine the effect of ZOL on survival, H22- s.c. and H22- i.p. mice were treated with 100 μg/kg body weight in the peritoneal cavity every other day for a total of four injections starting from 10 days and 3 days after inoculation of tumor cells (Figure 5). Among the H22- s.c. mice, the survival time in the ZOL-treated group was significantly longer than in the control group (P < 0.05), with five mice having a long survival and the survival of the dead mice over 45 days after tumor inoculation. Among the H22- i.p. mice, all animals (n = 10) in control group died within 10-25 days, whereas those in the ZOL-treated group died within 15–35 days, with two mice having a long survival. 8 DISCUSION In the present study, ZOL showed a significant inhibitory effect on the growth of SK-HEP-1 and H22 cells in a dose-dependent manner. To elucidate the effect of ZOL on the cell cycle progression, flow cytometry analysis of DNA content was performed on SK-HEP-1 and H22 cells. (Figure 2). A clear dose-dependence of the S phase arrest effect was noticed. This effect is in accordance with the previous studies on different types of tumor cells, all suggesting arrest of ZOL-treated cells in the S-phase [15-17]. ZOL may fail to induce apoptosis. As shown in Figure 2, in both hepatocellular carcinoma cell lines, ZOL treatment did not induce obvious apoptotic sub-G1 peak. We also found no evident characteristics of apoptosis (such as cell shrinkage or cytoplasmic blebs) in SK-HEP-1 cellular morphology after ZOL treatment (Appendix 1). Clearly, further experiments are warranted to verify this assumption. To further elucidate molecular mechanisms of the S phase arrest, we examined the cdc2, p53, cyclin A and cyclin B1 expression in SK-HEP-1 and H22 cell lines treated with or without ZOL. According to Figure 3, ZOL induced S-phase arrest probably through downregulating cdc2 and upregulating cyclin A. The direct antitumor effect of ZOL on solid tumors in animal models has been demonstrated in several studies [11, 18, 19], and was further confirmed in our study (Figure 4). The average tumor volume of ZOL-treated mice was statistically smaller than that of untreated mice. However, it showed no significant difference between the 100 μg /kg and 400 μg/kg groups. This may result from the plateau effect of 100 μg/kg and thus no additional effect can be achieved with increased doses. Moreover, a life 9 span-prolonging effect were found in ZOL-treated mice group compared to the control group in H22- s.c. and H22- i.p. mice models (Figure 5). This effect of prolonging survival was also observed in mice with osteosarcoma [12]. We hypothesize that ZOL significantly inhibits HCC growth, thus extending the survival time. Our results suggest that ZOL may be a promising drug for the treatment of HCC. For most cancer types, pharmacokinetics of ZOL is the biggest obstacle that prevents us from translating the preclinical antitumor activity into use in clinical settings. Research shows that a routine clinical dose of 4 mg ZOL achieves a peak plasma concentration of 0.75–2 μM before it is rapidly absorbed by bones [20]. A single 30–100 μg/kg injection in mice is equivalent to a single 4 mg human dose [21, 22]. Updated evidence has showed the cytotoxic effects of ZOL in different cancer cell lines at concentrations of 10 μM of ZOL or higher in vitro [23]. It means that the dose of ZOL should be elevated, otherwise the direct antitumor effect of ZOL is not noticable. Obviously, it is unsuitable for human body and may produce serious toxicity according to drug instructions. However, this challenge posed by pharmacokinetics of ZOL may be overcome by combined use with transarterial chemoembolization (TACE). TACE has become a standard care for HCC patients who are not suitable for surgical or ablative treatment if extrahepatic metastases and advanced liver disease are absent [24]. The rationale for treatment of HCC by TACE is based on the dual blood supply of the liver. The nutrition of liver parenchyma is predominantly supplied by portal veins, and the majority of the blood supply to HCC is derived from the hepatic artery. TACE is designed to eliminate 10 the tumor blood supply by particle embolization and directly infuse cytotoxic chemical into the branch of the hepatic artery that feeds the tumor. Transarterial delivery of cytotoxic drugs combined with embolization of arterial feeders are shown to reduce the maximum plasma concentration and increase the concentration of cytostatic agent in the tumor compared to systemic treatments [25]. Consequently, the hurdle of pharmacokinetics will be resolved if ZOL can be applied in TACE, at least this will be possible according to our findings. Abbreviations ZOL, Zoledronic acid; HCC, hepatocellular carcinoma; s.c., Subcutaneous injection; i.p., Intraperitoneal injection; TACE, Transarterial chemoembolization. Competing interests The authors declare that they have no competing interests. Authors' contributions Quan Liu, Yonghui Tao and Dong Hua designed and performed the experiments, Quan Liu and Ruizhen Bai contributed to manuscript writing. Shujian chang analyzed the data. All authors read and approved the final manuscript. 11 REFERENCES 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. Jemal, A., F. Bray, M.M. Center, J. Ferlay, E. Ward, and D. Forman, Global cancer statistics. CA Cancer J Clin, 2011. 61(2): p. 69-90. El-Serag, H.B., Hepatocellular carcinoma. N Engl J Med, 2011. 365(12): p. 1118-27. Llovet, J.M., A.M. Di Bisceglie, J. Bruix, B.S. Kramer, R. Lencioni, A.X. Zhu, et al., Design and endpoints of clinical trials in hepatocellular carcinoma. J Natl Cancer Inst, 2008. 100(10): p. 698-711. Aapro, M., P.A. Abrahamsson, J.J. Body, R.E. Coleman, R. Colomer, L. Costa, et al., Guidance on the use of bisphosphonates in solid tumours: recommendations of an international expert panel. Ann Oncol, 2008. 19(3): p. 420-32. Di Salvatore, M., A. Orlandi, C. Bagala, M. Quirino, A. Cassano, A. Astone, et al., Anti-tumour and anti-angiogenetic effects of zoledronic acid on human non-small-cell lung cancer cell line. Cell Prolif, 2011. 44(2): p. 139-46. Koto, K., H. Murata, S. Kimura, N. Horie, T. Matsui, Y. Nishigaki, et al., Zoledronic acid inhibits proliferation of human fibrosarcoma cells with induction of apoptosis, and shows combined effects with other anticancer agents. Oncol Rep, 2010. 24(1): p. 233-9. Sewing, L., F. Steinberg, H. Schmidt, and R. Goke, The bisphosphonate zoledronic acid inhibits the growth of HCT-116 colon carcinoma cells and induces tumor cell apoptosis. Apoptosis, 2008. 13(6): p. 782-9. Rachner, T.D., S.K. Singh, M. Schoppet, P. Benad, M. Bornhauser, V. Ellenrieder, et al., Zoledronic acid induces apoptosis and changes the TRAIL/OPG ratio in breast cancer cells. Cancer Lett, 2010. 287(1): p. 109-16. Matsumoto, S., S. Kimura, H. Segawa, J. Kuroda, T. Yuasa, K. Sato, et al., Efficacy of the third-generation bisphosphonate, zoledronic acid alone and combined with anti-cancer agents against small cell lung cancer cell lines. Lung Cancer, 2005. 47(1): p. 31-9. Lamoureux, F., B. Ory, S. Battaglia, P. Pilet, M.F. Heymann, F. Gouin, et al., Relevance of a new rat model of osteoblastic metastases from prostate carcinoma for preclinical studies using zoledronic acid. Int J Cancer, 2008. 122(4): p. 751-60. Wakchoure, S., M.A. Merrell, W. Aldrich, T. Millender-Swain, K.W. Harris, P. Triozzi, et al., Bisphosphonates inhibit the growth of mesothelioma cells in vitro and in vivo. Clin Cancer Res, 2006. 12(9): p. 2862-8. Ory, B., M.F. Heymann, A. Kamijo, F. Gouin, D. Heymann, and F. Redini, Zoledronic acid suppresses lung metastases and prolongs overall survival of osteosarcoma-bearing mice. Cancer, 2005. 104(11): p. 2522-9. Coleman, R.E., H. Marshall, D. Cameron, D. Dodwell, R. Burkinshaw, M. Keane, et al., Breast-cancer adjuvant therapy with zoledronic acid. N Engl J Med, 2011. 365(15): p. 1396-405. Bradford, M.M., A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem, 1976. 72: p. 248-54. 12 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. Okamoto, S., K. Kawamura, Q. Li, M. Yamanaka, S. Yang, T. Fukamachi, et al., Zoledronic Acid Produces Antitumor Effects on Mesothelioma Through Apoptosis and S-Phase Arrest in p53-Independent and Ras prenylation-Independent Manners. J Thorac Oncol, 2012. Romani, A.A., S. Desenzani, M.M. Morganti, S. La Monica, A.F. Borghetti, and P. Soliani, Zoledronic acid determines S-phase arrest but fails to induce apoptosis in cholangiocarcinoma cells. Biochem Pharmacol, 2009. 78(2): p. 133-41. Ohnuki, H., K. Izumi, M. Terada, T. Saito, H. Kato, A. Suzuki, et al., Zoledronic acid induces S-phase arrest via a DNA damage response in normal human oral keratinocytes. Arch Oral Biol, 2011. Giraudo, E., M. Inoue, and D. Hanahan, An amino-bisphosphonate targets MMP-9-expressing macrophages and angiogenesis to impair cervical carcinogenesis. J Clin Invest, 2004. 114(5): p. 623-33. Hiraga, T., P.J. Williams, A. Ueda, D. Tamura, and T. Yoneda, Zoledronic acid inhibits visceral metastases in the 4T1/luc mouse breast cancer model. Clin Cancer Res, 2004. 10(13): p. 4559-67. Chen, T., J. Berenson, R. Vescio, R. Swift, A. Gilchick, S. Goodin, et al., Pharmacokinetics and pharmacodynamics of zoledronic acid in cancer patients with bone metastases. J Clin Pharmacol, 2002. 42(11): p. 1228-36. Gao, L., H. Deng, H. Zhao, A. Hirbe, J. Harding, L. Ratner, et al., HTLV-1 Tax transgenic mice develop spontaneous osteolytic bone metastases prevented by osteoclast inhibition. Blood, 2005. 106(13): p. 4294-302. Daubine, F., C. Le Gall, J. Gasser, J. Green, and P. Clezardin, Antitumor effects of clinical dosing regimens of bisphosphonates in experimental breast cancer bone metastasis. J Natl Cancer Inst, 2007. 99(4): p. 322-30. Peng, H., Y. Sohara, R.A. Moats, M.D. Nelson, Jr., S.G. Groshen, W. Ye, et al., The activity of zoledronic Acid on neuroblastoma bone metastasis involves inhibition of osteoclasts and tumor cell survival and proliferation. Cancer Res, 2007. 67(19): p. 9346-55. Bruix, J., M. Sala, and J.M. Llovet, Chemoembolization for hepatocellular carcinoma. Gastroenterology, 2004. 127(5 Suppl 1): p. S179-88. Huppert, P., Current concepts in transarterial chemoembolization of hepatocellular carcinoma. Abdom Imaging, 2011. 36(6): p. 677-83. 13 FIGURE LEGENDS Figure 1. Antiproliferative activity of ZOL SK-HEP-1 and H22 cells were incubated for 48 h in the absence (control, the inhibition rate = 0%) or presence of 0.5-350 μM ZOL. Viability of SK-HEP-1 (◆) and H22 ( ■ ) cells were evaluated by SRB. ZOL exhibited a significant antiproliferative activity against SK-HEP-1 and H22 with IC50 of 21.1 μM and 29.6 μM. Figure 2. Effect of ZOL on the cell cycle distribution Flow cytometry of DNA was performed on SK-HEP-1 (top) and H22 (bottom) cells. SK-HEP-1 and H22 cells were treated with 0 (control), 25, 50 and 100μM of ZOL for 48 h and were stained with PI for flow cytometric analysis. A clear cell cycle perturbation effect of ZOL was observed in a dose-dependent manner. With increased concentration of ZOL, the cells accumulated in the S phase, and G1 and G2/M phase cells were gradually decreased. A representative figure is shown from three separate experiments. Figure 3. Effect of ZOL on the expression of cell cycle regulatory proteins SK-HEP-1 and H22 cells were incubated with or without 20 μM ZOL. After 48 h of treatment, cell lysates were evaluated for levels of Cyclin B1, Cyclin A, cdc2 and p53 expression by western blotting. Cyclin B1 and p53 showed no significant change in expression, cdc2 was deeply decreased, and Cyclin A was upregulated in SK-HEP-1 14 and H22 cell lines after treatment with ZOL for 48 h. A representative figure is shown from three separate experiments. Figure 4. Effect of ZOL on H22- s.c. mice Kunming mice were subcutaneously implanted with 1x107 H22 cancer cells. When tumors reached a size of 60-90 mm3 (~10 days), mice were intratumorally treated with ZOL 100 μg/kg(■) or 400 μg/kg(▲) every other day for a total of four injections. PBS injection was used as control (◆). Tumor volumes were monitored every 3 days and mice were killed when tumors reached 2,000 mm3. The average tumor volume of ZOL-treated mice was statistically smaller than that of untreated mice. The average tumor volume was not significant in the 100 μg /kg and 400 μg/kg groups. Statistically significant difference is indicated (*) (P< 0.05). Figure 5. Kaplan–Meier survival curve ZOL prolonged the survival of both H22- s.c. (A) and H22- i.p. mice (B). Both ZOL groups(◆) were treated with 100μg/kg intratumorally every other day for a total of four injections. PBS injection (●) was used as control. Among the H22- s.c. mice, the survival time in the ZOL-treated group was significantly longer than in the control group (P < 0.05), with five mice having a long survival and the survival of the dead mice over 45 days after tumor inoculation. Among the H22- i.p. mice, all animals (n = 10) in control group died within 10-25 days, whereas those in the ZOL-treated group 15 died within 15 – 35 days, with two mice having a long survival. Statistically significant difference is indicated (*) (P< 0.05). FIGURE FIGURE 1 FIGURE 2 FIGURE 3 16 FIGURE 4 FIGURE 5 17 Appendix Appendix 1. Effect of ZOL on SK-HEP-1 cellular morphology. SK-HEP-1 cells were incubated for 48 hour in the absence (A) or presence of 21.1 μM ZOL (B). The cells were photographed through a phase-contrast microscope x100. No evident characteristics of apoptosis (such as cell shrinkage or cytoplasmic blebs) were observed. Appendix 2. The gating of each FACS data. 18 19