Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
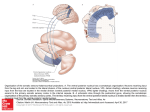
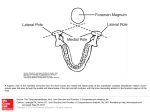
ACTA NEUROBIOL. EXP. 1977, 37: 247-252 THE ORGANIZATION OF AMYGDALOPETAL PROJECTIONS FROM THE LATERAL HYPOTHALAMUS AND PREOPTIC AREA IN THE RAT Liliana NITECKA, Olgierd NARKIEWICZ and Czeslaw JAKIEL Department of Anatomy, Institute of Medical Biology, School of Medicine Gdahsk, Poland In the light of physiological data it is certain that there is a close functional correlation between the amygdaloid body and the hypothalamus and preoptic area. These areas are vital for emotional reactions and affect drives and endocrinal functions. A damage of amygdaloid nuclei causes changes in behavior very similar to those following partial lesions of the hypothalamus and preoptic area (7, 8). As various investigations have shown, there are numerous connections running from the amygdaloid nuclei to the preoptic area and hypothalamus (3-5, 9, 11). They run both in stria terminalis and in the ventral amygdalofugal (amygdalar) pathway. The stria terminalis is a compact bundle of fibers, which divides near the anterior commissure into several components running in various directions (4, 5, 10); we distinguish, according to de Olmos (4), its dorsal, ventral and commissural component. The ventral amygdalofugal pathway assumes a compact bundle form only in the higher mammals, but is rather dissipated in the rat (15). In contrast to the fairly well-known amygdalofugal connections we havz only some data on amygdalopetal axons emerging from the diencephalon (1-3, 12-14, 16), while their topographical and physiological organization is almost unknown. For our studies on amygdalopetal connections we used 110 Wistar rats of both sexes, weighing 200-250 g. Various parts of the hypothala- mus, preoptic and neighboring areas were coagulated by direct current of 1 mA lasting 15-30 s. In several cases the stria terminalis was partly or wholly damaged in its supracapsular part. Postoperative survival time of animals was 3-5 days. Frozen sections of the brain were prepared according to the ' ~ a u t a - ~ y g a and x Fink-Heimer methods. Location and extent of lesion were determined on specimens stained by the Kliiver-Barrera method. Various lesions in the anteromedial part of the hypothalamus, in its posterior part, in the ventromedial and dorsomedial hypothalamic nuclei and in the medial preoptic area have not produced any marked degenerative changes in the amygdala. Massive terminal and preterminal degenerations in its nuclei were found in rats following lesions of the most lateral parts of the lateral preoptic area and lateral hypothalamus in its rostra1 segment, above the optic chiasma (Fig. 1 and 3). The whole area was defined by us as the preopticohypothalamic lateral area. Fig. 1. Localization of lesions causing degenerations in the amygdala. Extent of lesions is marked in black. A : Lesion in the lateral preoptic area (LP) causing severe degeneration of axons in the lateral amygdalopetal pathway of the stria terminalis. B: Lesion situated more caudally, in the rostrolateral part of the lateral hypothalamus (LH)resulting in degeneration of axons in the medial amygdalopetal pathway of the stria terminalis and in the ventral amygdalar pathway. After damage to the lateral preopticohypothalamic area, we found three bundles of fibers passing from that area to the amygdala, seen microscopically as bands of degenerating axons. Two bundles ran in the ventral component of the stria terminalis, occupying in its supracapsular part the lateral (lateral amygdalopetal pathway of the stria) and medial area (medial amygdalopetal pathway of the stria) (Fig. 2). The third pathway comprised fibers running directly to the amygdala (without entering the stria terminalis) in the ventral amygdalar pathway. They ran dispersed in a fan-like pattern. We named them as "ventral amygdalopetal pathway" (Fig. 2C). Fig. 2. Localization of degenerations in the amygdala following lesions in the lateral preopticohypothalamic area. In each row sections of the amygdaloid body from rostra1 to caudal. The broken lines indicate degenerating fibers of passage, the dotspreterminal and terminal degenerations. A: Degeneration of the lateral amygdalopetal pathway of the stria terminalis. B: Degeneration of the medial amygdalopetal pathway of the stria terminalis. C: Degeneration of the ventral amygdalopetal pathway (both, orsal and ventral component). At the upper left corner localization of the lateral and medial amygdalopetal pathways show11 on the cross-sectional schema of the components of the stria terminalis, Abbreviations: A, anterior amygdaloid area; BD, basal dorsal nucleus of amygdala; BV, basal ventral nucleus of amygdala; BN, bed nucleus of stria terminalis; C, commissural component of stria terminalis; Ce, central nucleus of amygdala; Co, cortical nucleus of amygdala; D, dorsal component of stria terminalis; L, lateral nucleus of amygdala; LH, lateral hypothalamus; LO, nucleus of lateral olfactory tract; LS, lateral amygdalopetal pathway of stria terminalis; M, medial nucleus of amygdala; MS, medial amygdalopetal pathway of stria terminalis; ST, stria terminalis; V, ventral component of stria terminalis. Fig. 3. Terminal degenerations in the anterior amygdaloid area (A), and in the medial nucleus of amygdala (B) following a lesion in the rostrolateral part of the lateral hypothalamus. C: Terminal degenerations in the basal dorsal nucleus of amygdala following a lesion in the rostra1 part of the lateral preoptic area. Nauta, Gygax. (>: 240). 1. The lateral amygdalopetal pathway of the stria terminalis begins in the cells lying rostrally in the preoptic lateral area (Fig. 1A). The fibers of this tract initially pass through the lateral area of bed nucleus of the stria terminalis. Farther on they penetrate the ventral component of the stria terminalis and in its supracapsular part lie laterally to the comrnissural component (Fig. 2). They enter the amygdala posteriorly, passing mainly through the medial part of the central nucleus. Terminal and preterminal degeneration was found, above all, in the medioventral area of the medial and posterior segment of the basal dorsal nucleus and, in a lesser degree in the medial part of the central nucleus (Fig. 2A). If the area of lesion was small and occupied only the rostra1 part of the preopticohypothalamic lateral area, there was no definite degeneration in the other amygdalopetal pathways nor in the rest of the amygdaloid nuclei. 2. The medial amygdalopetal pathway starts in the neurons of the preopticohypothalamic lateral area more caudally mainly in the lateral hypothalamus (Fig. 1B).The amygdalopetal fibers of this pathway enter the stria terminalis after passing through the lateral area of the bed nucleus of the stria terminalis more caudally than the fibers of the lateral amygdalopetal pathway. In the supracapsular part of the stria terminalis the fibers of the medial amygdalopetal pathway are situated in the ventral component of the stria, medially to its commissural component (Fig. 2). They are slightly less numerous and penetrate the amygdaloid body more medially than the former fibers. They end mainly dorsally, in the posterior and intermediate portions of the medial nucleus of amygdala (Fig. 2B). After cutting the supracapsular part of the stria terminalis we found degenerating fibers in the stria between the site of lesion and the amygdaloid body. These fibers, located mainly in the ventral portion of the stria, lay in two bundles medially and laterally similar to those after damage to the preopticohypothalamic lateral area. Following the lesion of the stria the extent of degeneration in the amygdaloid nuclei was generally similar to that of the preopticohypothalamic lateral lesion. The only definite difference was the additional degeneration in the ventroposterior part of the cortical nucleus of the amygdala. 3. The ventral amygdalopetal pathway similarly to the medial one, begins in the neurons lying for the most part caudally in the hypothalamic lateral area. Its fibers are slightly finer than the amygdalopetal axons of the stria terminalis. Owing to the location of the terminal degeneration in the amygdala (Fig. 2C) as well as to the course of the amygdalopetal fibers, it is possible to distinguish two not clearly separated components in the ventral amygdalofugal pathway: the ventral and the dorsal. The fibers of the ventral component run as a rule transversally to the medial part of the anterior amygdaloid area and to the anterior part of the medial nucleus. Here the terminal degeneration is very massive. In the medial nucleus of amygdala the projection area of the ventral amygdalopetal pathway partly overlaps the one caused by lesion in the stria terminalis. It was comparatively easy to distinguish the degenerating fibers of both systems, as the fibers of the ventral amygdalopetal pathway generally finer and their course is transversal, while the fibers of the stria terminalis, enter the medial nucleus of amygdala from the dorsocaudal side. The fibers of the dorsal component run not only in a lateral direction but also posteriorly. They pass through the dorsal part of the anterior amygdaloid area, through the central nucleus and terminate a t the neurons of the anterior pole of the basal dorsal nucleus. The above shows that the lateral amygdalopetal pathway of the stria and dorsal component of the ventral amygdalopetal pathway terminate in the basal dorsal nucleus, partly in the central nucleus, i.e., in the areas of a high AChE activity. On the other hand, the medial pathway of the stria and the ventral component of ventral amygdalopetal pathway end in the nucleus medialis and in the anterior amygdaloid area, where the AChE activity is low. Some terminals of these pathway can be found in the cortical nucleus and rarely in the basal ventral nucleus, although our data do not provide sufficient evidence of the latter suggestion. Neither we found degenerations in the lateral nucleus. The area where according to our findings, the amygdalopetal axons begin may be defined as the most lateral part of the hypothalamus. Here the characterjstically large neurons of high AChE activity can be found. From them emerge axons rich in AChE, which, after passing through the lateral part of the bed nucleus of stria terminalis, appear mainly in the lateral area of the ventral portion of the stria and run towards the amygdaloid body. Some data seem to favor the idea that these axons connect the lateral preopticohypothalamic area with the amygdaloid body. Naturally cells of high AChE activity are not the only nerve cells of this area and amygdalopetal connections may in a great part begins in other types of neurons as well. On the other hand, experiments with horseradish peroxidase ( 6 ) show that large cells of this area send axons to the cortex and to the reticular formation. Conrad and Pfaff (I, 2) found some fibers projecting from the medial preoptic area and anterior hypothalamus to the medial amygdaloid nucleus. Observations of Swanson (14) and our results show the localization of amygdalofugal neurons in a more lateral region. Data concerning the localization of terminals arising from neurons of the lateral preoptic- ohypothalamic region in the amygdaloid body are also somewhat contradictory. Cowan et al. (3), following the lesions of the lateral hypothalamus, found terminal degenerations in all nuclei of the rat amygdala except its central nucleus. Swanson (14), using autoradiographic techniques, traced in the rat an amygdalopetal projection from the lateral preoptic area; axons of that projection terminate in most of the amygdala, but they were not found in its lateral nucleus and the nucleus of the lateral olfactory tract. The above-mentioned authors observed no striking differences in the supply of various areas of the amygdaloid nuclei by axons arising from the preopticohypothalamic area. In our material, massive terminal degenerations almost equally distributed in all areas of the amygdala (except the nucleus of the lateral olfactory tract) were observed only in those cases where a lesion localized in the lateral preopticohypothalamic region encroached, even slightly, on the anterior amygdaloid area. If a lesion was localized exactly in the preopticohypothalamic region without touching the neighboring structures, degenerating axons running in all the above amygdalopetal pathway terminated in defined areas of the amygdaloid body. The investigation was supported by Project 10.4.1.01 of the Polish Academy of Sciences. REFERENCES 1. CONRAD, L. C. A. and PFAFF, D. W. 1976. Efferent from medial basal forebrain and hypothalamus in the rat. I. An autoradiographic study of the medial preoptic area. J. Comp. Neur. 169: 185-219. 2. CONRAD, L. C. A. and PFAFF, D. W. 1976. Efferent from medial basal forebrain and hypothalamus in the rat. 11. An autoradiographic study of the anterior hypothalamus. J. Comp. veur. 169: 221-261. 3. COWAN, W. M., RAISMAN, G. nad POWELL, T. P. S. 1965. The conections of the amygdala. J. Neurol. Neurosurg. 28: 137-151. 4. De OLMOS, J. S. 1972. The amygdaloid projection field in the rat as studied with the cupric silver method. In B. E. Eleftheriou (ed.), The neurobiology of the amygdala. Plenum Press, New York, p. 145-205. 5. De OLMOS, J. S., and INGRAM, W. R. 1972. The projection field of the stria terminalis in the rat brain. J. Comp. Neurol. 146: 303-335. 6. DIVAC, J. 1975. Magnocellular nuclei of the basal forebrain project to neocortex, brain stem, and olfactory bulb. Review of some functional correlates. Brain Res. 93: 385-399. 7. FONBERG, E. 1976. The role of the amygdaloid nucleus in animal behaviour. Progr. Brain. Res. 22: 273-280. 8. FONBERG, E. 1969. The role of the hypothalamus and amygdala; food intake, alimentary motivation, and emotional reactions. Acta Biol. Exp. 29: 335-358. 9. HALL, E. 1963. Efferent connections of the basal and lateral nuclei of the amygdala in the cat. Am. J. Anat. 113: 139-151. 10. JOHNSTON, J. B. 1923. Further contribution to the study of the evolution of the forebrain. J. Comp. Neurol. 35: 337-481. 11. LEONARD, C. M. and SCOTT, J. W. 1971. Origin and distribution of the amygdalofugal pathways in the rat. An experimental neuroanatomical study. J. Comp. Neurol. 141: 313-331. 12. NAUTA, W. J. H. 1958. Hippocampal projection and related neural pathways to the mid-brain in the cat. Brain 81: 319-340. 13. NITECKA, L. and JAKIEL, C. 1977. Connections of the hypothalamicopreoptic lateral area with the amygdaloid nuclei in the rat. Folia Morph. (in press). 14. SWANSON, L. W. 1976. An autoradiographic study of the efferent connections of the preoptic region in the rat. J. Comp. Neurol. 167: 227-256. 15. VALVERDE, F. 1965. Studies on the piriform lobe. Harvard Univ. Press. Boston. 16. WAKEFIELD, C. and HALL, E. 1974. Hypothalamic projections to the amygdala in the cat. Cell. Tiss Res. 151: 499-508. Accepted 21 February 1977 Liliana NITECKA, Olgierd NARKIEWICZ and Czeslaw JAKIEL, Institute of Medical Biology. School of Medicine, Dqbinki 1, 80-211 Gdafisk, Poland.