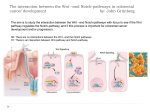
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
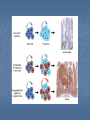
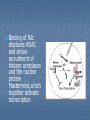
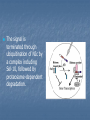
Signaling Pathways that Depend on Regulated Proteolysis 7 May 2007 Identified through genetic studies in drosophila. Highly conserved in evolution and are very important in animal development Wnt Notch Hedgehog NFB Wnts Wnt proteins released from or presented on the surface of signaling cells act on target cells by binding to Frizzled (Fz)/LDL-related protein (LDR) complex at the cell surface. Receptors Frizzled receptors, like GPCRs, are transmembrane proteins that span 7 times the plasma membrane. Their ligand-binding site is exposed outside the surface of the cell. Their effector site extends into the cytosol. Ligands Their ligands are Wnt proteins. These get their name from two of the first to be discovered, proteins encoded by wingless (wg) in Drosophila and its homolog Int-1 in mice. Fz receptors transduce signals to intra-cellular proteins including Dsh, GSK-3ß, Axin, APC and ßcatenin Nuclear ß-catenin interacts with lymphoid enhancer-binding factor 1/T cell-specific transcription factor (LEF/TCF) to affect transcription. In vertebrates, Wnt proteins are inhibited by direct binding to either secreted frizzled-related protein (SFRP) or Wnt inhibitory factor (WIF). SFRP is similar in sequence to the cysteinerich domain (CRD) of Frizzled, one of the Wnt receptors. GSK-3b dependent phosphorylation, ubiquitination and complex formation with the proteins axin and APC are important to regulate the cytoplasmic stability of beta-catenin protein in the wnt-signal transduction pathway Interaction with TCF transcription factors and the transactivation domains of betacatenin are instrumental to activate/derepress wnt-target genes in the nucleus. Complex formation with cadherins and alpha-catenin at the plasma membrane is essential for the role of betacatenin in cell adhesion. In vertebrate development, loss of a single Wnt gene can produce dramatic phenotypes that range from embryonic lethality and CNS abnormalities to kidney and limb defects These diverse phenotypes indicate that the Wnt pathway has distinct transcriptional outputs. In many cases, the cell, rather than the signal, determines the nature of the response, and up- or down-regulation of Wnt target genes is cell-type specific. WNT TARGET GENES AND FEEDBACK LOOPS WNT SIGNALING IN CANCER AND HUMAN DISEASE In adults, mis-regulation of the Wnt pathway also leads to a variety of abnormalities and degenerative diseases From crypt physiology to colon cancer Current evidence indicates that the Wnt cascade is the single most dominant force in controlling cell fate along the crypt-villus axis. In Tcf4 -/- neonatal mice, the villus epithelial compartment appears unaffected but the crypt progenitor compartment is entirely absent, implying that physiological Wnt signalling is required for maintenance of the crypt progenitor phenotype. The Wnt pathway in colon cancer The APC gene was originally discovered to be the culprit in a hereditary cancer syndrome termed familial adenomatous polyposis (FAP). FAP patients, inheriting one defective APC allele, develop large numbers of colon polyps, or adenomas, early in life. Individual polyps are clonal outgrowths of epithelial cells in which the second APC allele is inactivated. Mutational inactivation of APC leads to the inappropriate stabilization of b-catenin, implying that the absence of functional APC transforms epithelial cells through activation of the Wnt cascade. In some cases of colorectal cancer in which APC is not mutated, the scaffolding protein axin 2 is mutant, or activating (oncogenic) point mutations in b-catenin remove its N-terminal Ser/Thr destruction motif. Multiple studies have used a candidate gene approach to address the nature of the Tcf4 target gene programme in colorectal cancer. “Non-canonical” wnt signaling Non-canonical wnt signaling Involved in different development processes, but its intracellular mediators are ill-known. In vertabrates, Wnt-11 & Wnt-5a can activate Wnt/JNK pathway (planar cell polarity?) Wnt/Ca2+ pathway (described only in Xenopus & Zebrafish) Additional wnt pathways (described only in drosophila) In drosophila, non-canonical wnt signaling regulates “cell motility” through focal adhesion kinase (FAK)Planar cell polarity involved in tissue engineering In vertabrates, non-canonical wnt signaling regulates “gastrulation” through protein kinase C (PKC)Planar cell polarity involved in tissue engineering Wnt/Ca2+ signaling regulates dorso-ventral axis patterning (Wnt5aCa2+CalcineurinNFATActivation of target genesNegative regulation of canonical wnt signalingVentralization) Notch Signaling Pathway A brief history In 1917, Thomas Hunt Morgan described a strain of Drosophila with notches at the end of their wing blades, which result from haploinsufficiency Notch gene was cloned in the mid-1980s Notch is an essential gene encoding a signalling receptor that is required throughout development to regulate the spatial patterning, timing and outcomes of many different cell fate decisions in both vertebrate and invertebrate species. Notch is a single spanning transmembrane protein,which has a modular architecture. Ligands: Delta1, 3 and 4; Jag1, 2 Receptors: Notch1-4 Notch signaling has effects in many different organs •Notch signalling can maintain stem cells or precursor populations in an undifferentiated state •Notch signalling influences binary cell-fate decisions via lateral or inductive signalling •A third property of Notch is its ability to influence differentiation and cell-cycle progression The best known role of notch signaling is in nerve cell development Nerve cells arise within an epithelial sheet of precursor cells. Each future nerve cells signals to its immediate neighbours not to develop in the same way at the same time Lateral inhibition Lateral inhibition is a contact-dependent signaling that is mediated by the ligand delta. Delta displayed on the surface of future neural cell binds to notch on neighbours If the signal is defective, excess production of neurons at the expense of epidermal cell causes lethality. Processing and Activation of Notch S1 cleavage occurs within the secretory pathway so that a processed heterodimeric form is transported to the cell surface (furin protease in the golgi apparatus. Processing and Activation of Notch S2 cleavage occurs following ligand binding by Delta or Serrate(Jagged in mammals) through their DSL domains (Delta/Serrate/Lag2), and releases a membrane tethered form of the Notch intracellular domain. The latter is a constitutive substrate for the S3 cleavage, which releases the soluble intracellular domain of Notch (NIc). NIc is translocatedto the nucleus where it binds via the RAM domainand ankyrin repeats to a transcription factor, Suppressor of Hairless (Su(H)), or CBF1 in vertebrates. In the absence of a Notch signal, Su(H)/CBF1 can repress transcription through the recruitment of a histone deacetylase (HDAC) Binding of NIc displaces HDAC and allows recruitment of histone acetylases and the nuclear protein Mastermind,which together activate transcription The signal is terminated through ubiquitination of NIc by a complex including Sel-10, followed by proteosome-dependent degradation. The NICD-RBP-Jk complex up-regulates expression of primary target genes of Notch signaling such as HES in mammals, and E(spl) (for Enhancer of Split) in Drosophila. The HES/E(spl) family is a basic helix-loop-helix (bHLH) type trancriptional repressor and acts as Notch effectors by negatively regulating expression of downstream target genes such as tissue-specific transcription factors. Consistent with this view, HES1 and HES5, for instance, were shown to be upregulated by NICD and necessary to prevent neuronal differentiation of neural precursor cells from mouse embryos Modulators of Notch signalling Fringe: regulate notch Numb: notch inhibitor Neuralized (E3 ubiquitin ligases): regulate ligand Delta. Mib (mindbomb): (E3 ubiquitin ligase): function on Delta Fringe Fringe encodes a glycosyltransferase that adds O-fucoseglycans to the Notch EGF repeats. Fringe proteins might differentially modulate the response of Notch receptors to different DSL (Delta,Serrate and LAG2) ligands. Fringe can enhance Delta binding to the notch receptor. Whether fringe modification of Notch inhibits Serrate/Jagged binding is much less certain. Function autonomously Numb Numb domains: 1. PTB domain; N-terminal phosphotyrosine binding domain 2. proline-rich C-terminal region. In vitro studies have shown that Numb binds directly to NICD. The Cterminal half of the PTB domain and the Nterminus of Numb are required to inhibit Notch. Numb also has two motifs associated with endocytic proteins. mammalian Numb (mNumb) localizes to clathrin coated pits and early endosomes, might target endocytosed NICD for proteosomal destruction. Numb acts either upstream of S3 cleavage site of Notch or inhibit the endocytosis of membrane-bound activated Notch. Hedgehog Signaling Sonic Desert Indian Hedgehog Processing of Hedgehog (Hh) precursor protein Adding cholesterol to a glycine residue, Splitting the molecule into two fragments, Leaving the N-terminal signaling fragment with an attached hydrophobic cholesterol moiety. Addition of a palmitoyl group to the N-terminus Findings from genetic studies in Drosophila indicate that two membrane proteins, Smoothened (Smo) and Patched (Ptc), are required to receive and transduce a Hedgehog signal to the cell interior. They are required to activate transcription of the same target genes (e.g., wingless) during embryonic development. Model of the Hedgehog (Hh) signaling pathway In the absence of Hh, Patched (Ptc) protein inhibits Smoothened (Smo) protein. In the absence of Smo signaling, a complex containing the Fused (Fu), Costal-2 (Cos2), and Cubitis interuptus (Ci) proteins binds to microtubules. Ci is cleaved in a process requiring the ubiquitin/proteasome-related F-box protein Slimb, generating the fragment Ci75, which functions as a transcriptional repressor. Model of the Hedgehog (Hh) signaling pathway In the presence of Hh, inhibition of Smo by Ptc is relieved. Signaling from Smo causes hyperphosphorylation of Fu and Cos2, and disassociation of the Fu/Cos2/Ci complex from microtubules. This leads to the stabilization of a full-length, alternately modified Ci, which functions as a transcriptional activator inconjunction with CREB binding protein (CBP). Hedgehog signaling is conserved throughout the animal kingdom, and functions in the formation of many tissues and organs. Mutations in components of the Hedgehog signaling pathway have been implicated in birth defects such as cyclopia, a single eye resulting from union of the right and left brain primordia, and in multiple forms of human cancer (basal cell carcinoma).