Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
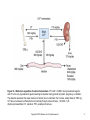
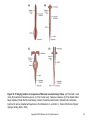
Chapter 14 Regulation of Calcium and Phosphate Homeostasis in Vertebrates Copyright © 2013 Elsevier Inc. All rights reserved. Figure 14-1 Endocrine regulation of calcium homeostasis. PTH and 1,25-DHC are hypercalcemic agents, and CT is the only hypocalcemic agent protecting the skeleton during periods of growth, pregnancy, or lactation. The skeleton represents the major reservoir of calcium ions in mammals. For humans, a daily intake of 1000 mg Ca2+/day is necessary to offset what is lost normally through urine and feces. 1,25-DHC, 1,25dihydroxycholecalciferol; CT, calcitonin; PTH, parathyroid hormone. Copyright © 2013 Elsevier Inc. All rights reserved. 2 Figure 14-2 An osteoclast. This multinucleate bone-destroying cell is a target for calcitonin that inhibits its activity. Interleukin-1 (IL-1) and macrophage colony-stimulating factor (MCSF) can enhance osteoclast survival. Tartrateresistant acid phosphatase (TRAPase) is considered a marker for osteoclast activity. Osteoclast activity and the ratio of osteoclasts to osteoblasts is increased by PTH, but indirectly (see text). Note the ruffled border of the cell adjacent to the resorption space (white area) that is involved in the active resorption of Ca 2+. Production of H+ and the lysosomal enzyme cathepsin K results in dissolution of the bone matrix and release of calcium and phosphate ions. (Adapted with permission from Martin, T.J. and Udagawa, N., Trends in Endocrinology and Metabolism, 9, 6–12, 1998. © Elsevier Science, Inc.) Copyright © 2013 Elsevier Inc. All rights reserved. 3 Figure 14-3 A multinucleate turtle osteoclast. (Courtesy of the late Nancy B. Clark, University of Connecticut, Storrs.) Copyright © 2013 Elsevier Inc. All rights reserved. 4 Figure 14-4 An osteoblast. Boneforming osteoblasts are targets for PTH, PTHrP, and 1,25-DHC. Stimulated osteoblasts transform osteoclast progenitor cells into mature osteoclasts through direct surface contact. The osteoclastdifferentiation factor (ODF) has not been identified. gp 130, glycoprotein 130; PTHrP, parathyroid hormone-related peptide; IL, Interleukin; VDR, Vitamin D receptor. (Adapted with permission from Martin, T.J. and Udagawa, N., Trends in Endocrinology and Metabolism, 9, 6–12, 1998. © Elsevier Science, Inc.) Copyright © 2013 Elsevier Inc. All rights reserved. 5 Figure 14-5 Embryonic derivatives of the pharyngeal pouches in vertebrates. The numbers refer to pouch number with anterior to the left. Pouch 1 remains only as the spiracle (selachians) or the eustachian tube. The thyroid actually develops from the pharynx floor between the first and second pouches. Parathyroids appear first in amphibians. In reptiles, the origin of the thymus (not shown) may be from pouches 2 and 3 (lizards), 3 and 4 (turtles), or 4 and 5 (snakes). In mammals, the origins of parathyroids and thymus are reversed. The ultimobranchial body (UB) is absent in mammals, and the calcitonin-secreting cells migrate to the thyroid instead of the UB. Copyright © 2013 Elsevier Inc. All rights reserved. 6 Figure 14-6 Amino acid sequences of 31 mammalian PTHs and PTHrP. Only the first 34 amino acids are shown. Explanation of the amino acid abbreviations is provided in Appendix C. Copyright © 2013 Elsevier Inc. All rights reserved. 7 Figure 14-7 A history of the parathyroid hormone receptor (PTHr) gene family. The simplest explanation for this phylogenetic tree is that an ancestral PTHr gene gave rise to a precursor gene at the first vertebrate genome duplication (R1) to a PTHr1/3 precursor gene that at the second vertebrate duplication (R2) gave rise to PTHr1 (red line) and PTH3r (blue line). The second precursor gene resulting from R1 (PTHr2/4) gave rise at R2 to PTHr2 (green line) and a fourth gene, Pth4r (yellow line), that somehow was lost. Initially, both fish and tetrapod lineages had three PTH receptor genes (PTHr1, PTHr2, and PTHr3), but later PTHr3 was lost in mammals and PTHr2 was lost in birds. (Adapted with permission from Bhattacharya, P. et al., Journal of Endocrinology, 211, 187–200, 2011.) Copyright © 2013 Elsevier Inc. All rights reserved. 8 Figure 14-8 Relationship of calf parathyroid secretion of PTH to experimental manipulation of plasma calcium levels. Plasma Ca2+ levels above 8 mg/100 mL (about 1 nM) suppress PTH secretion whereas lower levels increase PTH secretion. (Adapted with permission from Mayer, G.P. and Hurst, J.G., Endocrinology, 102, 1036–1042, 1978. © The Endocrine Society.) Copyright © 2013 Elsevier Inc. All rights reserved. 9 Figure 14-9 A classification of chordates, including the tunicates, in relation to their phylogenetic origins and time in terms of palaeontologicalperiods with some members of the parathyroid hormone gene family. Parathyroid hormone genes have been identified in the elephant shark (Callorhinchus milii; Pth1 and Pth2), the pufferfish (Takifugu rubripes; Ptha and Pthb), chicken, and mammal (Pth). Parathyroid hormonerelated protein genes (PTHrP) have been identified in elephant shark, pufferfish, chicken, and mammals. PTHr1 and PTHr2 genes have been identified in an ascidian (Ciona intestinalis) and zebrafish (Danio rerio) and PTHr1 in a frog (Xenopus laevis). The PTHr3 gene has only been identified in zebrafish. (Adapted with permission from Danks, J.A. et al., General and Comparative Endocrinology, 170, 79–91, 2011; Bhattacharya, P. et al., Journal of Endocrinology, 211, 187–200, 2011.) Copyright © 2013 Elsevier Inc. All rights reserved. 10 Figure 14-10 Unrooted phylogenetic tree of parathyroid hormone family peptides. PTH-L, PTH-like; TIP39, Tuberoinfundibular peptide of 39 residues. (Adapted with permission from Pinheiro, P.L.C. et al., BMC Evolutionary Biology, 12, 110, 2012.) Copyright © 2013 Elsevier Inc. All rights reserved. 11 Figure 14-11 Amino acid sequences for calcitonins. The sequence of the first 10 residues is highly conserved when comparing teleosts, birds, and mammals. The presence of valine (V) at position 8 increases the biological activity of the molecule four to five times. Hence, salmon-I is more potent in humans than is human CT. Note also that human and rat CTs are more like each other than like bovine or porcine, which are both similar. Explanation of the amino acid abbreviations is provided in Appendix C. (Adapted with permission from Matsumoto, A. and Ishii, S., “Atlas of Endocrine Organs,” Springer-Verlag, Berlin, 1992.) Copyright © 2013 Elsevier Inc. All rights reserved. 12 Figure 14-12 Synthesis of 1,25-dihydroxycholecalciferol (1,25-DHC). In terrestrial vertebrates, 7dehydrocholesterol is changed sequentially by the skin, liver, and kidney. Although some 24,25-DHC is also produced, it exhibits very weak binding to the vitamin D receptor and is probably not very important. Note the rearrangement of the A ring as a consequence of opening up the B ring. Copyright © 2013 Elsevier Inc. All rights reserved. 13 Figure 14-13 Sites of 1,25-DHC action in vertebrates. Targets associated specifically with calcium regulation are highlighted. (Adapted with permission from LeBlanc, G.A. et al., “Detailed Review Paper: State of the Science on Novel In Vitro and In Vivo Screening and Testing Methods and Endpoints for Evaluating Endocrine Disruptors,” prepared by RTI International for U.S. Environmental Protection Agency, Washington, D.C., 2012.) Copyright © 2013 Elsevier Inc. All rights reserved. 14 Figure 14-14 “The Ugly Duchess.” This painting originally was thought to be done by Leonardo Da Vinci but is now attributed to the painter Quentin Matsys in 1513. The subject of the painting is Margaret, Countess of Tyrol, who is suspected to have had a form of Paget’s disease that led to severely enlarged bones. The original painting resides in the National Gallery in London and is supposed to have been a source for some of the original illustrations in Lewis Carroll’s “Alice’s Adventures in Wonderland.” Copyright © 2013 Elsevier Inc. All rights reserved. 15 Figure 14-15 Varying location of corpuscles of Stannius in selected bony fishes. (A) The bowfin, Amia calva. (B) A salmonid, Salvelinus pluvius. (C) The Crucian carp, Carassius carassius. (D) The striped mullet, Mugil cephalus. Recall that the head kidney contains the adrenocortical cells in teleosts and is otherwise lymphoid in nature. (Adapted with permission from Matsumoto, A. and Ishii, S., “Atlas of Endocrine Organs,” Springer-Verlag, Berlin, 1992.) Copyright © 2013 Elsevier Inc. All rights reserved. 16 Figure 14-16 Distribution and phylogenetic relationships of vertebrate stanniocalcins (STCs). (Adapted with permission from Roch, G.J. and Sherwood, N.M., Integrative and Comparative Biology, 50, 86–97, 2010.) Copyright © 2013 Elsevier Inc. All rights reserved. 17 Figure 14-17 Location of ultimobranchial tissue in nonmammalian vertebrates. (A) Goldfish. (B) Newt. (C) Snake. (D) Lizard. (E) Bird. (Adapted with permission from Matsumoto, A. and Ishii, S., “Atlas of Endocrine Organs,” Springer-Verlag, Berlin, 1992.) Copyright © 2013 Elsevier Inc. All rights reserved. 18 Figure 14-18 Location of parathyroid glands in selected tetrapod vertebrates. (A) The newt Cynops pyrrhogaster; urodele amphibians typically have one pair of parathyroids. (B) Frogs and other anurans typically have two pairs of parathyroids. (C) Lizards have one pair of parathyroids except for Anolis spp. that have two. (D) Snakes, such as Agkistrodon halys, have two pairs of parathyroids; the posterior pair is associated with the thymus gland. (E) Many birds such as the seagull (Larus argentatus), shown here, and domestic species have two pairs of parathyroids located near the thyroid although some (e.g., stork, quail) have one pair. (Adapted with permission from Matsumoto, A. and Ishii, S., “Atlas of Endocrine Organs,” Springer-Verlag, Berlin, 1992.) Copyright © 2013 Elsevier Inc. All rights reserved. 19 Figure 14-19 Phylogenetic relationship of the vertebrate vitamin D receptor (VDR) for 1,25dihydroxycholecalciferol (1,25-DHC). (Adapted with permission from Reschly, E.J. et al., BMC Evolutionary Biology, 7, 222, 2007.) Copyright © 2013 Elsevier Inc. All rights reserved. 20