Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
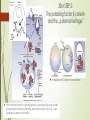
1869-1941 Nobel Prize 1935 Otto Mangold 1891-1962 Hilde Mangold 1898-1924 Pieter Nieuwkoop 1917-1996 AXIS FORMATION II. Vertebrates „Theories come and theories go. The frog remains.” Jean Rostand (1960) XENOPUS: D-V THE THREE SIGNAL MODEL (3SM) 2 before gastrulation during gastrulation 1) vegetal cells secrete a mesendoderm-inducing factor that converts the marginal zone (MZ) into a ring of mesoderm 2) The vegetal cell mass is subdivided into dorsal (= Nieuwkoop Centre) and ventral parts that induce dorsal and ventral mesoderm, respectively; 3) the most dorsal mesoderm (Spemann’s / SpemannMangold organiser /SMO/) secretes signals that establish DV polarity by promoting dorsal identity Spemann-organizer induces patterning of the germ layers the secondary Experiments leading up to the three-signal model (3SM). a Schematic of an amphibian late blastula stage embryo after germ layer induction; animal points up, vegetal points down, ectoderm is shown in blue, mesoderm in red, endoderm in yellow. b When cultured in isolation, animal and vegetal explants differentiate into ectoderm and endoderm, respectively; in animal–vegetal co-cultures, mesoderm (and some endoderm) is induced in the animally derived tissue suggesting an inducing signal emanating from the vegetal tissue (red arrows). c Dorsovegetal cells induce dorsal mesoderm (purple) whereas ventrovegetal cells induce ventral mesoderm, even if the animal tissue is rotated by 180. d The 3SM for germ layer formation: (1) a signal emanating from vegetal cells of the embryo induces the mesoderm in an equatorial ring (red arrows), (2) a signal from dorsovegetal cells dorsalises the mesoderm on the dorsal side (purple arrow) and (3) signals from the dorsal mesoderm (Spemann’s organiser) pattern the embryo along its DV axis (black arrow). D, dorsal; V, ventral Maternal heritage → 3SM: STEP 1 secreted TGF-b proteins: Activin Xnrs (Xenopus Nodalrelated factors) activation of the Smad2/4 pathway in the MZ Buc: Bucky ball Macf1: Microtubule-actin cross-linking factor 1 MZ → mesoderm MZ→MES Vg1, VegT activin, Xnr ↑ Vg1, VegT 4 Xenopusban a MZ több rétegű • a felszíni réteg entoderma precursorokat ad, • a mélyebb rétegek mesodermalis precursorokat adnak Rana: • az MZ felszíni rétege is ad mesodermalis sejtvonalakat Kofron et al., Dev 2007 3SM STEP 2: The polarizing factor b-catenin and the „paternal heritage” localization of β-catenin accumulation Dobrowolski and De Robertis, Nat. Rev. 2012 Wnt11 mRNA and protein signaling ventrally → balancing of Axin level ventrally by sequestration of the axin containing „destruction complex” into MVB → newly synthesized β-catenin accumulation b-catenin accumulation → Nieuwkoop centre Eomes: Eomesodermin • direct result of TGFbBMP mediated mesoderm induction • T-box transcription factor Eomes→Xbra SMO MZ Nieuwkoop centre Goosecoid, etc. Nieuwkoop Centre ↓ Spemann-Mangold Organizer (SMO) 7 MZ Vg1, VegT Activin, Xnr → ADMP (anti-dorsalizing morphogenetic protein, a BMP-like paracrine factor) Functions of the SMO The ability to self-differentiate dorsal (axial) mesoderm (prechordal plate: pharyngeal entoderm, axial head mesoderm, chordamesoderm) ((= roof of the archenteron in Rana and Hemichordata! (see the neural induction)) Initiate the gastrulation – remember bottle cells and the mechanoeffector b-catenin The ability to dorsalize 3SM STEP 3 the surrounding mesoderm into paraxial (somite-forming) mesoderm when it would otherwise form ventral mesoderm by Smad2/4, Twin/ Siamois → chordin, noggin, follistatin secretion by axial mesoderm : Bmp inhibitors SMO chorda 10 • Dorsalizing the ectoderm = induce formation of the neural tube nervous system forms from that region of the ectoderm that is protected from epidermal induction by BMP-inhibiting molecules A Hemichordata ősbéltető homológ a Chordata gerinchúrral, De • Rana ősbéltető Nem zajlik le az enterocoelia folyamata, így nem alakul ki lateralis mezoderma (hiányzik pl. a somitomeria) Hemichordata neuruláció, Balanoglossus simodensis Ketzer András BSc szakdolgozata, 12. ábra (Miyamoto és Wada, 2013) Larsen’s Human embriology Fate of the ectoderm / NEURULATION (1) "default fate„ is to become neural tissue (this feature originates from the nuclear b-catenin); 11 (2) certain parts of the embryo induce the ectoderm to become epidermal tissue by secreting BMPs (Bmp4/2/7); (3) the organizer tissue acts by secreting molecules that block BMPs, thereby allowing the ectoderm "protected" by these BMP inhibitors to become neural tissue. Deuterostomia – Prostomia D-V axis inversio 12 CNS CNS Deuterostome anchestor, living Hemichordata INVERTER: SMO = „Vertebrate innovation” References 13 Gilbert, S.F. Developmental Biology, 9th ed. 2010 Kiecker, C., Bates, T. Bell, E. Molecular specification of germ layers in vertebrate embryos Cell. 2016 Mol. Life Sci. 73:923–947 DOI 10.1007/s00018-015-2092-y Kofron, M. at al. 2007 Wnt11/β-catenin signaling in both oocytes and early embryos acts through LRP6-mediated regulation of axin. Development 134: 503-513; doi: 10.1242/dev.02739 Levin, AJ., Brivanlou, AH. Proposal of a model of mammalian neural induction. 2007 Developmental Biology 308: 247–256 doi:10.1016/j.ydbio.2007.05.036 Rossant, J. Tam, PPL. Blastocyst lineage formation, early embryonic asymmetries and axis patterning in the mouse. 2009 Development 136: 701-713; doi: 10.1242/dev.017178 Saitou, M., Yamaji, M. Germ cell specification in mice: signaling, transcription regulation, and epigenetic consequences. 2010 Reproduction 139: 931-942 2010, doi: 10.1530/REP-10-0043