Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
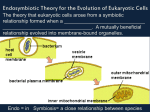
Master Microbiology Evolution of the Eukaryotic Cell LITERATURE Dagan T, Martin W (2004) The tree of one percent. Genome Biology 7: 118 Embley TM, Martin W (2006) Eukaryotic evolution, changes and challenges. Nature Reviews 440: 623-630 Timmis JN, Ayliffe MA, Huang CY, Martin W (2004) Endosymbiotic gene transfer: organelle genomes forge eukaryotic chromosomes. Nature Reviews Genetics 5: 123-135 Prof. Dr. Ralf Rabus AG Allgemeine und Molekulare Mikrobiologie Institut für Chemie und Biologie des Meeres (ICBM) © Ralf Rabus, www.icbm.de Contents Prokaryotes versus Eukaryotes Tree of life? Five current views of microbial evolution Archezoa - early-branching eukaryotic lineages? Mitochondria in multiple guises Models of eukaryote origin Timing and ecological context of eukaryote origins Genetic control of biogenesis of mitochondria and chloroplasts Genome reduction in mitochondria and plastides © Ralf Rabus, www.icbm.de Prokaryotes versus Eukaryotes Bacterial cell Animal cell Plant cell Cell wall Plasmamembrane Plasmamembrane Cell wall Plasmamembrane Ribosome Polysome Cytoplasm Chromosome Plasmid Ribosome Polysome Cytoplasm Mitochondrium Goli apparatus ER Nuclear membrane Nucleus Nucleolus Ribosome Polysome Cytoplasm Mitochondrium Goli apparatus ER Nuclear membrane Nucleus Nucleolus Pore Chlorplast Vacuole © Ralf Rabus, www.icbm.de Fuchs Allg. Mirko. (2006) Abb. 2.2 Tree of life: the bifurcation dilemma Classical view of evolution: phylogeny as a tree-like process of lineage splitting tree of life = single bifurcating tree lateral gene transfer (LGT) no major impact on evolution microbial genomes are related by a series of bifurcations LGT is important among prokaryotes proportion of prokaryotic genes affected by LGT: 2-60% LGT occurred throughout microbial history LGT is not tree-like microbial evolution undepictable by a single bifurcating tree Endosymbiotic gene transfer (among eukaryotes) adds to the bifurcation dilemma mitochondria originate from an ancestral D-proteobacterium chloroplasts originate from an ancestral cyanobacterium © Ralf Rabus, www.icbm.de Dagan Allg. & Mirko. Martin (2006) Tree of life: genomic perspective (I) Automatable procedure for reconstructing the tree of life identify protein families that are universally distributed among all genomes detect cases of LGT (unusual tree topologies) exclude such proteins and reiterate the procedure 31 presumably orthologous proteins sequences present in 191 genomes each 31 proteins represent about 1% / 0.1% of an average prokaryotic / eukaryotic proteome tree of 1% or 0.1% exclusion of all non-universally distributed proteins and suspected cases of LGT Support a Gram-positive origin of Bacteria and suggest a thermophilic last universal common ancestor © Ralf Rabus, www.icbm.de Ciccarelli et al. (2006) Tree of life: the genomic perspective (II) Archaea Eukaryota Bacteria © Ralf Rabus, www.icbm.de Ciccarelli et al. (2006) Fig. 2 Tree of life: the genomic perspective (III) non-redundant set of human proteins against all proteins from 224 prokaryotic genomes 200 eubacteria 31 universal proteins for tree of life no. of proteins % amino-acid identity 24 archaebacteria © Ralf Rabus, www.icbm.de Dagan & Martin (2006) Fig. 2 Five current views of microbial evolution The classical rRNA-derived tree The introns-early tree The neomuran tree The symbiotic tree The prokaryote-host tree © Ralf Rabus, www.icbm.de Dagan & Martin (2006) The classical rRNA-derived tree The universal ancestor (progenote): communal collection of information-storing and processing entities not yet organized as cells LGT main mode of genetic novelty "Genetic annealing" gives rise to cells: order of domain branching off: Eubacteria, Archaebacteria and then Eukaryotes refractory to LGT and endosymbiotic GT negligible for evolution © Ralf Rabus, www.icbm.de Dagan & Martin (2006) Fig. 1a The introns-early (or eukaryotes-first) tree The ancestral state of genes might be "split" some introns in eukaryotes = carryovers from the assembly of primordial protein-encoding regions organization of eukaryotic genes (having introns) represents that of the first genome intronless prokaryotes would be a derived conditions © Ralf Rabus, www.icbm.de Dagan & Martin (2006) Fig. 1b The neomuran tree The common ancestor: free-living eubacterium (Chlorobium-like anoxygenic phototroph) Eubacteria only organisms on Earth until 900 Mio. years ago At this time an Actinobacterium-like eubacterium lost its murein-containing cell wall reinvention of the cell wall by a group of rapidly evolving organisms (Neomura) invention of isoprene ether lipid synthesis Æ archaebacteria phagotrophy Æ eukaryotes Accounts for cell biological characters, but not for sequence similarities among genes © Ralf Rabus, www.icbm.de Dagan & Martin (2006) Fig. 1c The symbiotic tree: a merger of distinct branches Ancestor of eukaryotes: endosymbiosis of prokaryote X in host prokaryote Y formation of nucleated (n) cells Subsequent separate endosymbiotic events between eukaryotic host and prokaryotes: ancestor of plastides (p) and mitochondria (m) © Ralf Rabus, www.icbm.de Dagan & Martin (2006) Fig. 1d The prokaryote-tree with LGT: a merger of ephemeral genomes Extensive LGT throughout microbial evolution (inset 1) No mitochondria-lacking eukaryotes observed so far Endosymbiotic origin of plastids and mitochondria (endosymbiotic event in a prokaryote!) ring-like relationship (inset 2) between ancestral organisms rather than a tree © Ralf Rabus, www.icbm.de Dagan & Martin (2006) Fig. 1e Where to go? Recover and depict both, the tree-like and non-tree-like mechanisms of microbial evolution tree-like: vertical inheritance through common descent non-tree-like: LGT and endosymbiosis © Ralf Rabus, www.icbm.de Dagan & Martin (2006) Archezoa - early-branching eukaryotic lineages? Archezoa mostly anaerobic or parasitic eukaryotes once thought to lack mitochondria first: divergence of Archezoa; second: mitochondrial aquisition © Ralf Rabus, www.icbm.de Embley & Martin (2006) Fig. 1 Mitochondria in multiple guises Archezoa are now known to contain mitochondria-related organelles hydrogenosomes and mitosomes (also in ciliates and fungi, not grouped with Archezoa) common trait: double membrane and conserved mechanisms of protein import share at least one further trait with mitochondria Absence of traditional mitochondria and presence of a specialized anaerobic phenotype are neither rare nor "primitive" as once thought Aerobic and anaerobic eukaryotes, harbouring mitochondrial homologues of various sorts, have co-existed throughout eukaryote history © Ralf Rabus, www.icbm.de Embley & Martin (2006) Mitochondria Possess a genome encoding components involved in oxidative phosphorylation Transport of ATP into cytosol Key enzymes of anaerobic metabolism (e.g. pyruvate:ferredoxin oxidoreductase) in anaerobically functioning mitochondria (e.g. protists like Euglena) © Ralf Rabus, www.icbm.de Embley & Martin (2006) Fig. 2a Hydrogenosomes No genome Oxidation of pyruvate to H2, CO2 and acetate Key enzymes of anaerobic metabolism (e.g. pyruvate:ferredoxin oxidoreductase) ATP-generation via substrate-level phosphorylation Transport of ATP into cytosol Single common ancestry of mitochondria and hydrogenosomes very likely © Ralf Rabus, www.icbm.de Embley & Martin (2006) Fig. 2b Mitosomes No genome Undergone more evolutionary reduction than hydrogenosomes No direct role in ATP synthesis In Giardia: two mitochondrial proteins of Fe/S cluster assembly Occurrence in: eukaryotes with cytosolic ATP synthesis energy parasites © Ralf Rabus, www.icbm.de Embley & Martin (2006) Fig. 2b Models of eukaryote origin 1. Nucleus bearing, amitochondriate cells 2. Acquisition of mitochondria in an eukaryotic host © Ralf Rabus, www.icbm.de 1. Origin of mitochondria in a prokaryotic host 2. Acquistion of eukaryote-specific features Embley & Martin (2006) Fig. 4 Timing and ecological context of eukaryote origins: classical view Diversified unicellular microfossils (widely accepted as eukaryotes) appear in strata of ~1.45 billion years (Gyr) Minimum age of eukaryotes at ~1.45 Gyr Fossilized Bangiomorpha (strongly resembling modern bangiophyte red algae) appear in strata of ~1.2 billion years (Gyr) Minimum age of plant kingdom at ~1.2 Gyr Two main stages in early eukaryotic evolution early emergence and diversification of anaerobic, amitochondriate lineages acquisition of an oxygen-respiring mitochondrial ancestor in one lineage Æ diversification of aerobic eukaryotic lineages global rise in atmospheric oxygen levels at ~2 Gyr ago Æ "environmental disaster" for cells lacking the mitochondrial endosymbiont © Ralf Rabus, www.icbm.de Embley & Martin (2006) Timing and ecological context of eukaryote origins: critical view Contemporary anaerobic eukaryotes did not branch off before the origin of mitochondria New isotopic studies indicate that anaerobic environments persisted locally and globally over the past 2 Gyr oxygen appeared first in the atmosphere at ~ 2 Gyr ago up until ~ 600 Myr ago the ocean existed in an intermediate oxidation state • oxygenated surface water (where photosynthesis occurred) • sulfide-rich (sulfidic) and oxygen-lacking (anoxic) subsurface water "oxygen event" in the atmosphere has to be decoupled from anoxic marine environments: • anaerobic eukaryotes living on the margins of an oxic world • still valid today (e.g. water column of the Black Sea) © Ralf Rabus, www.icbm.de Embley & Martin (2006) Genetic control of biogenesis of mitochondria and chloroplasts c Massive transfer of ancestor derived genes into the nucleus Æ only few genes retained in the genomes of the organelle d Organelles strongly depend on nuclear genes Æ 90% of proteins imported from cytosol e DNA still transferred from organelles to nulceus c e d e c © Ralf Rabus, www.icbm.de Timmis et al. (2006) Fig. 1 Genome reduction in mitochondria and plastides Sequenced mitochondrial genomes encode 3 - 67 proteins modern D-proteobacterium Mesorhizobium loti: 7 Mb genome encodes >6,700 proteins Sequenced plastide genomes encode 20 - 200 proteins cyanobacterium Nostoc punctiforme: >9 Mb genome encodes >7,200 proteins enormous reduction of organelle genome Genome reduction in organelles versus parasites parasites: reduction through specialisation to a nutrient-rich intracellular environment Æ loss of genes that are no longer needed organelles: reduction through export of essential genes to the host´s genetic apparatus Æ import of thousands of essential proteins from the cytosol © Ralf Rabus, www.icbm.de Reduction of chloroplast genome Time course of gene relocation massively at the onset of endosymbiosis continued during lineage diversification The same core set of genes (photosynthesis and translation) retained in all lineages Displayed lineage diversification red algae (Porphyra) angiosperms (flowering plants) Cyanophora (belonging to the most ancient lineage of photosynthetic eukaryotes) © Ralf Rabus, www.icbm.de Timmis et al. (2006) Fig. 2 Acquisition of plant-like genes in trypanosomes Martin and Borst (2003) PNAS 100:765-767