Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
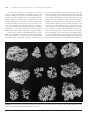
Rhodoliths and Rhodolith Beds Michael S. Foster, Gilberto M. Amado Filho, Nicholas A. Kamenos, Rafael Riosmena-Rodríguez, and Diana L. Steller ABSTRACT. Rhodolith (maërl) beds, communities dominated by free living coralline algae, are a common feature of subtidal environments worldwide. Well preserved as fossils, they have long been recognized as important carbonate producers and paleoenvironmental indicators. Coralline algae produce growth bands with a morphology and chemistry that record environmental variation. Rhodoliths are hard but often fragile, and growth rates are only on the order of mm/yr. The hard, complex structure of living beds provides habitats for numerous associated species not found on otherwise entirely sedimentary bottoms. Beds are degraded locally by dredging and other anthropogenic disturbances, and recovery is slow. They will likely suffer severe impacts worldwide from the increasing acidity of the ocean. Investigations of rhodolith beds with scuba have enabled precise stratified sampling that has shown the importance of individual rhodoliths as hot spots of diversity. Observations, collections, and experiments by divers have revolutionized taxonomic studies by allowing comprehensive, detailed collection and by showing the large effects of the environment on rhodolith morphology. Facilitated by in situ collection and calibrations, corallines are now contributing to paleoclimatic reconstructions over a broad range of temporal and spatial scales. Beds are particularly abundant in the mesophotic zone of the Brazilian shelf where technical diving has revealed new associations and species. This paper reviews selected past and present research on rhodoliths and rhodolith beds that has been greatly facilitated by the use of scuba. Michael S. Foster, Moss Landing Marine Laboratories, 8272 Moss Landing Road, Moss Landing, California 95039, USA. Gilberto M. Amado Filho, Instituto de Pesquisas, Jardim Botânico do Rio Janeiro, Rua Pacheco Leão 915, 22460-30, Rio de Janeiro, Brazil. Nicholas A. Kamenos, School of Geographical and Earth Sciences, Gregory Building, University of Glasgow, Glasgow, G12 8QQ, Scotland. Rafael Riosmena-Rodríguez, Programa de Investigacíon en Botaníca Marina, Departamento de Biologia Marina, Universidad Autónoma de Baja California Sur, Apartado postal 19-B, La Paz, B.C.S. 23080, Mexico. Diana L. Steller, Moss Landing Marine Laboratories, 8272 Moss Landing Road, Moss Landing, California 95039, USA. Correspondence: Michael S. Foster, foster@mlml .calstate.edu. Manuscript received 25 January 2012; accepted 5 March 2013. INTRODUCTION Relative to other abundant nearshore communities like kelp forests and coral reefs, rhodolith or maërl beds have been little investigated by marine ecologists. Even in clear, shallow water, their lack of vertical structure in the water column makes them difficult to distinguish from entirely soft bottoms. Observed and sampled from the bottom with scuba, however, these purple and pink fields are complex communities formed by aggregations of unattached, nongeniculate (lacking uncalcified joints) coralline algae with individual thalli ranging in size on the order of one to tens of cm and in shape from highly branched twigs and fans to spheres. Live individuals may be stacked a few cm deep, often grading below into dead fragments mixed with carbonate and terrigenous sediment (Figure 1). They provide hard habitat for numerous other marine algae that live on their surfaces, and for invertebrates living on and in the rhodoliths and surrounding sediments (see the “Rhodolith Systems: Communities and Conservation” section in this paper). There are few descriptive or experimental data to characterize the environmental conditions that lead to rhodolith bed formation and persistence. It does appear that beds are generally found on fairly level bottoms where light is sufficient for growth and water motion and bioturbation are sufficient to prevent burial and anoxia from sedimentation and to move the rhodoliths. High water motion, however, can inhibit bed formation by transporting individuals out of otherwise suitable habitat or causing mechanical 144 • smithso n i a n co n trib u tio n s to the m a ri n e scie n ces FIGURE 1. Rhodolith beds around Isla El Requéson, Baja California Sur, Mexico. Aerial shot: Dense aggregations of rhodoliths appear as dark ribbons and occur to a depth of 12 m. The island is 100 m across at widest point. The wide gray diagonal bar in lower right is the airplane wing strut (photo by M. S. Foster). In situ photo: spherical Lithophyllum margaritae rhodoliths at a depth of 6 m; diameters are 4–5 cm (photo by D. L. Steller). Core cylinder photo: Core taken through the bed at depth of 6 m. Note stratification, with living plants at the surface grading into dead fragments and fine sediment below; diameter = 9 cm (photo by D. L. Steller). destruction (reviewed in Foster, 2001). These conditions can result in free-living individuals of other organisms such as corals and bryozoans co-occurring with rhodoliths (e.g., James et al., 2006). Once thought to be primarily subtropical, they are now known to occur from tropical to polar waters and from the low intertidal zone to 150 m deep (Bosence, 1983; Foster, 2001). Our understanding of rhodoliths and rhodolith beds has grown from a synergy between geologists and biologists. Geologists were the first to become interested in rhodoliths, albeit dead ones, because these corallines are well preserved in the fossil record. At least since Darwin’s (1844) observations of rhodoliths (“nullipores”) in calcareous strata in the Cape Verde Archipelago, rhodolith deposits have been used to interpret geological change and paleoenvironments (e.g., Bassi et al., 2009). Early ecological work on live beds was pursued in the intertidal zone, or subtidally with dredges or grabs, and was focused on species composition and natural history (e.g., Weber-van Bosse and Foslie, 1904). In contrast to attached organisms on hard bottoms, dredging in rhodolith beds can provide good samples of the n u mber 3 9 organisms present. This method is still being used to locate beds and their boundaries (e.g., Foster et al., 1997), especially over large areas where scuba is inefficient and acoustical mapping equipment (e.g., Ehrhold et al., 2006) is not available. Dredging can be effective for species information and mapping, but as in other shallow-water habitats, observation of microhabitat distribution and use, studies of processes, and field experimental manipulations require scuba. One of the first and the most comprehensive in situ studies on rhodolith systems was by Bosence (1976), a geologist interested in using characteristics of living rhodoliths and rhodolith beds to better interpret fossil deposits. Bosence (1976) carefully mapped rhodolith distribution in a bay in Ireland, and examined the relationships between the environment (especially water motion) and rhodolith species distribution and morphology. He wrote the first review of rhodolith ecology (Bosence, 1983), highlighting the advantages of scuba as a research tool and providing a foundation and stimulus for future studies that now include the full spectrum of ecological topics and approaches, including past and present climate change. Rhodoliths are particularly useful for the latter as they are widely distributed, grow slowly (~mm/yr) and, like temperate trees and hermatypic corals, leave a record of their growth as bands within the thallus (see the “Rhodoliths as Environmental Recorders” section in this paper). Sadly, reduced growth and dissolution caused by future changes in ocean pH and temperature may result in the absence of bands and the decline of rhodolith beds. A thorough review of rhodoliths and rhodolith bed ecology is beyond the scope of this paper. Instead, each author discusses a current topic in his or her area of interest and expertise: Introduction (MSF), Diving to the Edge of Rhodolith Taxonomy and Evolution (RRR), Rhodolith Communities and Conservation (DLS), Deep Beds in Brazil (GAF), and Rhodoliths as Environmental Recorders (NAK). Investigations of these topics have all been greatly facilitated by the use of scuba. We hope the reader will find the discussions interesting and informative, stimulating further interest in and study and conservation of these tumble weeds of the sea. DIVING TO THE EDGE OF RHODOLITH SYSTEMATICS AND EVOLUTION Seaweed distribution and the factors that affect it are key scientific questions; Setchell (1893) clearly understood the importance of temperature limits to the geographic distribution of kelp species and the importance of distribution to systematics. The utility of such information, however, depends on the validity of the species identification and how well collections represent the geographic and depth distributions of species in nature. These problems have been particularly difficult to overcome in coralline red algae (Corallinales, Rhodophyta), and especially with the nongeniculate species that occur as rhodoliths. The characteristics used to classify rhodoliths in the early 1700s could not segregate • 145 them from corals or stones (Woelkerling, 1988) and names such as Lithophyllum are still in use for both algae and corals (Veron, 1995). After recognizing rhodoliths as calcified, photosynthetic organisms, most investigators thought that species of Lithophyllum were characteristic of warm waters and Lithothamnium of cold waters (Foslie, 1900). Such relationships were based on sampling and observations from the intertidal zone or on specimens dredged from the shallow subtidal. Distributional knowledge was further compromised by the numerous species that were described based largely on external morphology. For example, investigators like Mikael Foslie described 485 species and varieties (Woelkerling et al., 2005) based primarily on external morphology, at least half of which were rhodolith-forming species. As a consequence of developments in underwater technology including scuba, other diving methods, and better remote sampling devices, it soon became clear that coralline red algae were abundant, widespread, and often a key component of the ocean floor at depths to nearly 300 m. These developments also dramatically increased extraction of live and dead rhodoliths (maërl) for commercial purposes, an activity that has occurred since the seventeenth century along French and U.K. coasts (Grall and Hall-Spencer, 2003; see the “Rhodolith Systems: Communities and Conservation” section in this paper). While large rhodolith beds have long been known in the northeast Atlantic, the surge in ocean exploration beginning in the twentieth century revealed the presence of coralline banks in Malaysia (Weber-van Bosse, 1904), the Galapagos (Lemoine, 1930), the eastern Pacific (Dawson 1960a, 1960b), and Brazil, where beds cover hundreds of square km (Kempf, 1970; see the “Deep Beds in Brazil” section in this paper). These explorations were followed by extensive surveys in Brazil (Lavrado, 2006) and around the Hawaiian Islands (Adey et al., 1982). Hundreds of species were described from deeper areas, strongly suggesting that more species would be found as surveys continued. However, these collections were all made by dredging, and the few specimens obtained may not have been representative of local variation. Taxonomic analyses could be biased by describing new species based on a limited number of specimens. This situation greatly improved in the 1950s when E.Y. Dawson started to use scuba for his research, diving with only mask, fins, tank, regulator, and a very rudimentary buoyancy compensator, with shorts and a shirt for a diving suit (Norris, 2010: figs. 7, 8). Dawson could observe and collect numerous representative specimens under water, including what became Porolithon castellum and Sporolithon pacificum, the latter a very distinctive species (Dawson, 1960b). The allure of the visual record meant that underwater photography began to be used in the early days of diving. Dawson soon added a camera to his equipment, and his dive partner took one of the first underwater photographs of a coralline alga. Scuba diving quickly became part of the usual collecting approach for many researchers. It was a boon for taxonomists, who need to collect hundreds of individuals for rigorous morphological, anatomical, and now molecular analyses. 146 • smithso n i a n co n trib u tio n s to the m a ri n e scie n ces As scuba was recognized as an essential tool for marine scientists, scientific diving facilities and training programs, both fixed and portable, were developed around the world. There are a number of excellent examples of the success of such facilities in helping contribute to our knowledge of coralline taxonomy and diversity, including the descriptions of Tenarea tesselatum (Litt ler, 1971) from the tropics and Ezo eppiyessoense (Adey et al., 1974) from a temperate region. Scuba investigations continue to discover new and interesting subtidal coralline species, including Synarthrophyton schielianum (Woelkerling and Foster, 1989), which has one of the strangest morphologies among the red algae. In situ ecological studies of rhodolith-forming species began in the 1970s, including the seminal studies of Bosellini and Ginsburg (1971) and Bosence (1976). These studies documented the large effects of the environment, especially variation in water motion, on rhodolith shape and branching characteristics. The results enlightened paleoenvironmental interpretations but also served as a warning to taxonomists that many described rhodolith species might be morphological variants of the same species. For Mediterranean waters, Ballesteros (1988) wrote the first review of rhodolith species using information obtained by diving, and Basso (1998) critically evaluated species boundaries. Taxonomic evaluations based on scuba collections using research stations in the western Pacific were done by Verheij (1993) in Indonesia and Ballesteros and Afonso-Carrillo (1995) in the Indo-Pacific. In the eastern Pacific, the diving study of Steller and Foster (1995) further showed the influence of the environment on rhodolith morphology. This environmentally induced morphological plasticity (Figure 2) obscured the real taxonomic boundaries used for the species until detailed anatomical work was done (Riosmena-Rodríguez et FIGURE 2. Growth-form variation in the rhodolith Lithophyllum margaritae from the Gulf of California. The small rhodolith in lower left corner is 1.5 cm wide. From Riosmena-Rodríguez et al., 1999. n u mber 3 9 al., 1999). Additional genetic information from Schaeffer et al. (2002) has shown that some isolation occurs among populations. New descriptions and studies of rhodolith beds (e.g., Harvey et al., 2005; Konar et al., 2006) and collections and species analyses of the rhodoliths found are helping to refine our understanding of growth forms, species, and species distributions. Rhodolith beds are distributed worldwide but a thorough bathymetric evaluation has not been produced at the global scale. Harvey and Woelkerling (2007) did an excellent review of the rhodolith- forming species in the coralline algae, concluding that there are 8 genera and 71 species cited in modern studies. Most of the records in this review came from dredging, not scuba collection, again suggesting that more of the latter is needed to clarify the boundaries between species. New technologies for working under water, along with the traditional methods of ocean exploration, provide very exciting opportunities that will contribute to understanding the evolutionary history of rhodolith-forming species via comparisons of subtidal findings with the geological record of well-preserved coralline red algae. Collections using scuba diving and/or remotely operated underwater vehicles (ROVs) have already provided new records and evolutionary insights based on research occurring around the world (e.g., Brazilian continental shelf, Farias et al., 2010, and Bahia et al., 2011; Galician estuaries, Peña et al., 2011; Gulf of California, Riosmena-Rodríguez et al., 2010). In the future, it might be particularly interesting to examine very old fossil deposits along the coast of the Bahamas where Littler et al. (1985) described the deepest known plant life, a coralline alga. Coralline algae originated in the Cambrian (Woelkerling, 1988) and are one of the oldest clades in red algae. New studies will likely change our current views on rhodolith taxonomy and systematics due to probable discoveries of presently unknown evolutionary clades. The current challenge with regard to rhodolith systematic biology is to use information from field collections in combination with genetic analyses to better resolve species and answer evolutionary questions. This needs to involve the use of nuclear, chloroplast, and mitochondrial genes to delimit the number of clades among rhodoliths. These genetic approaches, combined with correlations between molecular clock and isotopic ages from carbonate deposits, should allow the construction of a model based on evolution and development (EVO DEVO), as suggested by Aguirre et al. (2010). The origins and development of rhodolith deposits can also be explored using mitochondrial genes to determine the origins of populations and the relative importance to bed persistence of fragmentation versus recruitment from spores. This is the edge of taxonomy and evolution, and scuba will therefore continue to be essential to advances. RHODOLITH SYSTEMS: COMMUNITIES AND CONSERVATION Individual rhodoliths, or “nodules,” can form intricately branched structures, and large aggregations of nodules form • 147 beds, complex biogenic matrices of hard branches that create numerous interstitial spaces. Beds typically form as a living layer on top of an otherwise sedimentary bottom (Figure 1) and harbor a diverse and often unique assemblage of associated species (Cabioch, 1969; Keegan, 1974). In addition to being considered biodiversity hot spots, rhodolith beds are also settlement sites and nursery grounds for important commercial species. Fleshy macroalgae are found on the rhodolith surfaces; invertebrates are found among or bored into rhodolith branches (cryptofauna), moving over and among the rhodoliths, or burrowed into underlying sediments (infauna) (Grall and Glemarec, 1997; Steller et al., 2003). Burrowing fish add further benthic complexity, making these habitats rich sites for ecological exploration. Rhodolith beds are generally more species rich and support higher population densities than adjacent sedimentary habitats (Birkett et al., 1998; Steller et al., 2003). For example, stratified sampling of subhabitats in a rhodolith bed and a sand flat found richness to be 1.7 times, and total abundance 900 times, greater in a rhodolith bed than in adjacent non-rhodolith habitat (Steller et al., 2003). This has been attributed to the higher number of available niches in rhodoliths relative to sand. Beds support complex food webs of predators, suspension feeders, detritivores, and micrograzers in the eastern Atlantic (Grall et al., 2006). Early studies using scuba were largely devoted to determining the distribution of rhodoliths and associated species (Keegan, 1974; Bosence, 1979), and such studies encouraged the development of diver-operated sampling devices such as suction dredges to more precisely sample soft-bottom species (Keegan and Konnecker, 1973). This ecological community research continues as new beds are discovered and explored, and many recent studies have shifted their focus to understanding the mechanisms underlying ecological relationships, reflecting the evolution of ecological research as well as changes in research methods. Traditional ship-based benthic sampling with dredges and grabs has been useful in establishing general patterns of distribution and diversity. Combining this type of sampling with scuba allows for (1) efficient visual surveys of large areas, (2) more precise habitat sampling, (3) enhanced detection of rare, seasonal, and fragile associated species and species interactions, and (4) in situ environmental measurements and experimental manipulations. As a result, surveys worldwide now report that the greatest contributors to high richness and abundance are cryptofauna (primarily arthropods, annelids, and cnidarians) that increase as live cover and rhodolith size and branching increases. Such microhabitat associations can be obscured or eliminated in homogenized samples from dredges, grabs, and cores. Large-scale studies comparing species in rhodolith beds to those in entirely sedimentary habitats have measured species diversity of either the entire community or a dominant group of organisms. In general, higher diversity and abundance of a community (Grall et al., 2006), or of specific invertebrate groups (e.g., polychaetes; Figueiredo et al., 2007), has been correlated with higher density and cover of live rhodoliths and more complex habitat. This pattern is not consistent for all species; infauna 148 • smithso n i a n co n trib u tio n s to the m a ri n e scie n ces such as soft-sediment meiofaunal bivalves can be more diverse on dead (versus live) rhodolith substrate (Jackson et al., 2004). When rhodoliths grow as distinct, densely branched, spherical individuals (Figure 2) rather than mats of intertwined, jack- like structures, the richness and abundance of species in and on them increases with rhodolith size, branching, and available space (Steller et al., 2003; Hinojosa-Arango and Riosmena-Rodríguez, 2004; Foster et al., 2007). Elucidating such relationships requires using scuba to carefully collect individual specimens, followed by proper fixation and careful dissection. Such studies have shown that 2–6 cm–diameter, highly branched Lithophyllum margaritae and 5–9 cm–diameter, knobby, branched Lithothamnion muellerii from the Gulf of California can harbor 10–90 cryptofaunal species per rhodolith, with more species in larger individuals. Rhodolith densities can beupwards of 5,000–10,000 m−2 (D. Steller, personal observation), resulting in exceptionally high cryptofaunal abundance. The most abundant taxa often include crustaceans, polychaetes, ophiuroids, and mollusks. Collectively, these findings support the notion that large, old individuals act as “old-growth rhodoliths,” and are inhabited by species such as stomatopods and burrowing clams not found on and in smaller rhodoliths (Foster et al., 2007). Diving studies have also revealed rare and new species, and some unique and interesting ecological relationships. Eight species of small (2–10 mm) cryptofaunal chitons were collected from between the branches of Lithophyllum margaritae in the Gulf of California, four of which were previously undescribed (Clark, 2000). Reports of complex, fragile nests built by the mollusk Limaria hians, a gaping file shell, came from in situ observations (Hall-Spencer et al., 2003). Macroinvertebrates such as the urchin Toxopneustes roseus can cover themselves with rhodoliths and also prefer to eat them rather than fleshy algae (James, 2000). The burrowing crustacean Upogebia deltaura creates extensive burrows among the rhodoliths that can withstand disturbance (Hall-Spencer and Atkinson, 1999). Such investigations are still few but the use of standardized scuba-enabled subtidal sampling designs and methods (Steller et al., 2007) promises to reveal more new species, as well as ecological and biogeographic insights. The productivity of rhodolith beds is enhanced by a diverse associated flora that grows on the hard substrata they provide (Jacquotte, 1962; Grall et al., 2006), and some algal species appear to be confined to rhodolith habitat (Leliaert et al., 2009). The associated flora often changes seasonally, as well as with depth. These changes are correlated with variation in rhodolith characteristics such as cover, depth, and the amount of live material. The changes are also correlated with changing oceanographic conditions, such as water motion, which can turn and roll rhodoliths, as well as temperature (Cabioch, 1969; Lieberman et al., 1979; Hily et al., 1992; Birkett et al., 1998; Steller et al., 2003; Amado- Filho et al., 2007; Riul et al., 2009; Peña and Barbara, 2010). In a subtropical Gulf of California bed, Steller et al. (2003) reported up to thirty common macroalgal species in winter, but a maximum of eight species in summer. Year-long sampling at 1½-month intervals has shown similar large seasonal changes in the flora of a European Atlantic bed, as well as relationships between these changes and the depth of the living rhodolith layer and the live/ dead rhodolith ratio (Peña and Barbara, 2010). Qualitative in situ observations in the Gulf of California and elsewhere suggest that the diversity of large fishes in the water column above rhodolith beds is generally low, probably because there is little structure above the bottom. Reports of numerous associated water column fishes (e.g., Aburto-Oropeza and Balart, 2001) are likely an artifact of the beds being in close proximity to reefs that provide macrostructure. Numerous cryptic demersal, benthic, and/or burrowing species such as gobies and blennies are found on, in, and among rhodoliths, and juveniles of species such as cod have been reported to use rhodolith beds as feeding areas (Hall-Spencer et al., 2003). Large, bottom- dwelling or burrowing species can be quite common, including sheephead (Semicossyphus pulcher) in California, and tiger snake eels (Myrichthys maculosus), bullseye electric (Diplobatis ommanata) and other rays, and Cortez garden eels (Taeniconger digueti) in the Gulf of California (D. Steller and M. Foster, pers. obs.). The distribution of rhodolith beds appears to be largely constrained by abiotic variables such as water motion and light, but within-bed structure and perhaps even bed persistence may be strongly affected by biological interactions. Bioturbation by invertebrates and fishes moves rhodoliths and resuspends fine sediment, likely facilitating rhodolith growth and bed maintenance (Marrack, 1999). Rhodolith beds may act as nursery habitat for a variety of species. Bivalves are abundant in rhodolith beds in areas such as Georges Bank (Thouzeau, 1991), the Gulf of California (Steller and Caceras-Martinez, 2009), and the eastern Atlantic (Hily et al., 1992; Hall-Spencer, 1998; Hall-Spencer and Moore, 2000), and rhodolith beds may play a role as nursery habitats for scallops. Scallop–rhodolith bed interactions have been of particular interest because scallops are commercially harvested and their life history is well known. Scallop larvae have been shown to preferentially settle on living (versus dead) rhodoliths (Steller and Caceras-Martinez, 2009). This preference likely contributes to high juvenile scallop densities (Kamenos et al., 2004a)h@ due to greater scallop selection, attachment, and growth (Kamenos et al., 2004b, 2004c). Rhodolith beds may also benefit adult scallops by providing a refuge from predation (Steller et al., 2003; Kamenos et al., 2006). Patterns of water motion responsible for rhodolith maintenance may also influence larval delivery of a number of associated species, a mechanism that remains to be investigated. While natural disturbances are an essential feature of rhodolith ecosystems, anthropogenic disturbances can be catastrophic. This is readily clear from diving observations made over a once- thriving rhodolith bed that had been crushed by a scallop dredge. Beds have been entirely or partially destroyed by extraction of calcareous sediment, bottom fisheries, and degraded water quality from fish farms (De Grave, 1999; Hall-Spencer and Moore, 2000; Hall-Spencer et al., 2006). Smaller-scale disturbances such n u mber 3 9 as anchoring or mooring in coastal bays also occur, but have received less attention. In addition to localized destruction, rhodolith beds worldwide will likely be impacted by decreasing ocean pH linked to rising atmospheric CO2 (Kleypas et al., 2006). Coralline algae, with their very soluble high-Mg calcite, are predicted to be in the first suite of species severely impacted by ocean acidification (Kuffner et al., 2007; Jokiel et al., 2008). Chemical dissolution could make them more susceptible to other physical and biological disturbances and decrease post-disturbance recovery rates. The positive relationship between biodiversity and rhodolith density, nodule size, and branching strongly suggests that factors negatively impacting nodule characteristics would likely lead to negative community-level impacts. The need for protecting rhodolith beds from dredging, trawling, and other relatively small-scale disturbances has been recognized by conservation directives in the European Union, Australia, New Zealand, and Mexico (review in Riosmena- Rodríguez et al., 2010). On the largest scale, however, little progress has been made toward reducing CO2 emissions. It remains to be seen how effective local conservation measures will be in the face of global change. DEEP BEDS IN BRAZIL The Brazilian coast supports the largest known rhodolith beds in the world, covering extensive areas of the north, northeastern, and southeastern Brazilian continental shelf (Kempf, 1970; Milliman, 1977; Amado-Filho et al., 2007). However, our understanding of the latitudinal distribution and community structure of these beds is still in its infancy. Many rhodolith studies in Brazil, especially the early ones, were focused on the characterization of shelf sediments and prospects for potential commercial exploitation (Milliman and Amaral, 1974; Dias, 2000). Only a few published studies have considered the beds in a biological context that includes consistent information about meso-and small-scale distribution, bed structure, associated organisms, and the species composition of the rhodoliths (e.g., Gherardi, 2004; Riul et al., 2009; Villas-Boas et al., 2009; Amado-Filho et al., 2010). These latter studies have found a high diversity of organisms associated with rhodolith beds and highlight their ecological relevance. A general evaluation of the marine biota in the Brazilian exclusive economic zone (the marine area extending 320 km offshore) indicated rhodolith beds increase the diversity of epibenthic organisms, with Shannon diversity indices (H’) of 4.0–5.1 found to depths of 250 m in this ecosystem (Lavrado, 2006). These and other data obtained during the 1990s and the early 2000s were, however, based on dredging and the analyses were largely qualitative. Most rhodolith beds in Brazil still remain unexplored or poorly known ecologically. This is largely because the beds occur mostly in mesophotic habitats (~30–120 m depth) that are difficult to adequately access with standard scuba. • 149 As pointed out by Bridge et al. (2010), information on biotic and abiotic aspects of mesophotic habitats remains extremely scarce due to logistical and technological restrictions, particularly when compared with shallow- water habitats. This is true for rhodolith beds in Brazil, but recent advances in mixed-gas diving techniques (Figure 3), complemented by ROV observations and high-resolution, multibeam, bathymetric mapping systems, allow us to begin determining their extent, structure, and dynamics. These tools are now being used to investigate extensive areas of the continental shelf (e.g., Abrolhos Bank), tops of seamounts (Vitoria-Trindade Ridge), and around oceanic islands (e.g., Trindade Island and Fernando de Noronha Island; Figure 4). One of the most interesting features of these mesophotic habitats, recently investigated using technical diving, are rhodolith beds inhabited by populations of the deep water endemic kelp Laminaria abyssalis. This kelp occurs on the continental shelf at latitudes of 19°–23° S and depths of 45–120 m where the bottom is completely covered by rhodoliths (Amado-Filho et al., 2007). Kelp populations are closely associated with the rhodoliths; all L. abyssalis holdfasts are attached to one or more rhodoliths. Graham et al. (2007) suggested that such deep water kelp refugia are potential hot spots of tropical marine diversity and productivity. This suggestion has been confirmed in the Brazilian deep water rhodolith–kelp beds, as endemic species of different taxonomic groups have been cited to this area. The northern limit of Brazilian L. abyssalis populations is determined by the southern edge of Abrolhos Bank. The Abrolhos Shelf (16°50′S–19°45′S; Figure 4) is an ~6,000 km2 enlargement of the eastern Brazilian continental shelf, and encompasses the largest (~325 km2) and richest reefs in the South Atlantic (Leão and Ginsburg, 1997). These reefs are well known for their unique coral assemblages dominated by Brazilian-endemic, Neogene relics belonging to the genus Mussismilia. Investigations combining side scan sonar, ROV images, and scuba diving show that rhodolith beds are the predominant feature from north to south across the Abrolhos Shelf at depths from ~25 m to ~110 m. These beds are composed of at least five non-geniculate coralline species. The Vitoria- Trindade Ridge (VTR; 20°S–21°S) is a 1,150 km east–west chain of nine seamounts in the South Atlantic that includes two small and highly isolated islands at its eastern end (Figure 4). Previous biological sampling on the VTR was largely restricted to (1) dredging, (2) data from commercial fisheries, and (3) a few scientific diving operations in the shallow waters surrounding Trindade Island. The mesophotic zone on the top of two seamounts (Davis and Jaseur) were surveyed in March 2009 using a combination of ROVs, mixed-gas technical dives, and a single-beam bathymetry system. The seamounts have predominantly flattened tops that result from alternating periods of growth and erosion of carbonate algal deposits over volcanic pedestals during periods of high and low sea level (Almeida, 1965). At present sea level, the flattened summits of the seamounts are all situated in relatively shallow water, with minimum depths ranging from 40 m to 110 m. The seamount tops 150 • smithso n i a n co n trib u tio n s to the m a ri n e scie n ces FIGURE 3. Technical diving equipment being used to take video images at 60 m on Fernando de Noronha Island shelf, Brazil. Photo by Z. Matheus, courtesy of G. M. Amado Filho. rise up to 50 m from the bottom, and are dominated by rhodoliths with a high diversity of associated species including seaweeds, sponges, corals, black corals, octocorals, and reef fishes. Rhodolith beds are also the main feature surrounding Fernando de Noronha, a typical tropical island (03°50′S, 32°25′W, about 345 km east of the coast of Brazil; Figure 4) known as the best diving location in Brazil. These beds, found down to 120 m at the edge of the island shelf, were also discovered using technical diving. The ongoing description and study of these unique coralline environments will provide essential information for understanding the ecology, biodiversity, and connectivity of South Atlantic reef communities. RHODOLITHS AS ENVIRONMENTAL RECORDERS FIGURE 4. Locations of mesophotic rhodolith beds on the Brazilian Shelf. Coralline algae have several characteristics that make them ideal candidates for recording the environment in which they grow. When these algae grow, they lay down annual and subannual carbonate-derived growth bands composed of high-Mg calcite (Figure 5; Henrich et al., 1996; Kamenos et al., 2008). Groups of subannual primary growth bands resulting from variation in cell size and wall thickness can be grouped into dark n u mber 3 9 • 151 FIGURE 6. A Lithothamnion glaciale bed and associated fauna on the west coast of Scotland. Scale = 5 cm. Photo by N. Kamenos. FIGURE 5. Annual banding patterns in a transverse sectioned Lithothamnion glaciale branch. Scale = 1 mm. Photo by N. Kamenos. and light pairs that form higher-order annual banding patterns (Freiwald and Henrich, 1994). While Lithothamnion glaciale and Clathromorphum compactum show clear annual banding patterns (Halfar et al., 2008; Kamenos et al., 2008), other species such as Phymatolithon calcareum show clearer subannual banding patterns (Blake and Maggs, 2003). In some species, growth is not hampered by prolonged periods of low temperature (e.g., L. glaciale; Henrich et al., 1996) or darkness (e.g., L. glaciale; Freiwald and Henrich, 1994), while in others there is reduced growth in winter (e.g., C. compactum; Halfar et al., 2008). As new thalli grow at the surface of the rhodolith bed, older thalli become covered by sediment, die, and form part of the dead deposit. Individual coralline algal thalli can live to at least ~850 years (Frantz et al., 2005) and accumulations of these algal thalli have created maërl deposits spanning the last 20,000 years (Figure 6; Bosence, 1983). Unlike most carbonate-depositing organisms, which have a more restricted distribution, coralline algae occur from polar (Schwarz et al., 2005) to tropical (Littler et al., 1991) shallow seas. Three key steps are typically used in the development of an environmental recorder (known as a proxy; Kamenos et al., 2009). For coralline algae this involves the following: (1) calibration, which involves understanding the biology of the algae and experimentally determining how growth, for example, responds to environmental stimuli such as temperature (Kamenos et al., 2008); (2) validation of the algae as proxies, which requires appropriate biogeochemical analyses to determine if the observed response of the alga’s physical or chemical characteristics to environmental change are a direct response to that change or the record of a tertiary response to a related physiological process (Kamenos et al., 2009); and, (3) application, which establishes that the coralline algae are recording a specific environmental stimulus and can be used to reconstruct and understand environmental changes that occurred before the availability of instrumental records (Kamenos, 2010; Burdett et al., 2011). Unfortunately, many proxies do not undergo this rigorous and important three-step process, rendering their utility questionable. Scuba has been instrumental in the development of rhodoliths as environmental recorders. Because of their fragile nature, hand collection is the only suitable technique for calibrating the algae in the field or collecting them for calibration in the laboratory. Subsequent to calibration, collection of samples for paleoenvironmental reconstruction involves determination of the most suitable sampling location (e.g., the location likely to provide the longest record) and precise manipulation of sampling apparatus (e.g., corer placement or airlift attachment). These tasks can only be reliably achieved using on-site sampling via scuba. Remote mechanical collection is not suitable due to the absence of specific sampling location information (e.g., what was the orientation of the surface algae on collection?). While suitable remote coring equipment can be used in softer sediments (e.g., vibra corer), the dense nature of rhodolith deposits requires coring techniques capable of penetrating the sediment without causing excessive disturbance. At present, scuba is the ideal tool for conducting such coring activities. 152 • smithso n i a n co n trib u tio n s to the m a ri n e scie n ces Relationships between environmental parameters and growth have been observed in coralline algae. Significant negative relationships are present between temperature and calcite density in Lithothamnion glaciale (Kamenos and Law, 2010), as well as temperature and growth-band width in Clathromorphum compactum (Halfar et al., 2011). In both species growth characteristics were correlated to decadal-scale temperature records at collection sites but were characterized by noticeable variability. That variability was attributed to localized control of incident radiation (e.g., algal blooms) and thus it is likely that growth contains a record of both temperature and irradiance (Kamenos and Law, 2010). This was addressed by the development of a growth-environment model that accounts for both temperature and light (as photosynthetically active radiation or PAR; Burdett et al., 2011). By combining a record of summer calcification within individual Lithothamnion glaciale growth bands with known temperature records for the area, cloud cover (indicative of PAR) history on the west coast of Scotland was reconstructed that indicated a modest increase in cloud cover trends since 1910 (Burdett et al., 2011). The chemistry of the calcite deposited by coralline algae within their growth bands also serves as an environmental recorder. Initial observations indicated elemental concentrations within coralline algae varied directly (Chave and Wheeler, 1965) and indirectly (Moberly, 1968) with environmental fluctuations. More recently, Mg concentrations within Lithothamnion glaciale have been observed to represent ambient temperature of the seawater in which the algae grew at fortnightly resolutions (Kamenos et al., 2008), as well as sea-surface temperature at seasonal resolution (Halfar et al., 2000). This response allowed Lithothamion glaciale to be validated as a paleotemperature recorder (Kamenos et al., 2009), enabling the first biweekly resolution reconstruction of Atlantic marine temperatures since ~1350 (Kamenos, 2010). δ18O in Clathromorphum nereostratum and Lithothamnion glaciale represents ambient seawater temperature at seasonal resolutions and has been used to reconstruct northeastern Atlantic sea-surface temperature since 1970 (Halfar et al., 2000, 2008). Lithothamnion muellerii and C. nereostratum have been shown to record atmospheric 14C concentrations (Frantz et al., 2000, 2005), which are critical for determining long-term growth rates and age in coralline algae as well as dating the exact timing of climatic events recorded by the growth-banding structure or chemistry within the algae. Overall, coralline algae are ideally suited as chemical environmental recorders because (1) they do not suffer diagenetic effects due to the presence of the living membrane covering the carbonate skeleton (Alexandersson, 1974); (2) there is no stress-related geochemical deviation in Mg/Ca–temperature relationships (Kamenos et al., 2008); and, (3) there is no non- equilibrium- associated oxygen isotope fractionation in maërl (Rahimpour-Bonab et al., 1997; Halfar et al., 2007). Scuba has played a central role in facilitating an understanding of how coralline algae act as environmental recorders through their utility in paleoenvironmental reconstructions. With the need for increasingly resolved spatio-temporal paleoclimate records to better understand recent climate trends, these algae are beginning to receive increased attention as environmental recorders. CONCLUSIONS As they have for other shallow subtidal environments, in situ investigations using scuba have improved our understanding of rhodoliths and rhodolith beds far beyond that achievable by dredging and other remote research techniques. Scuba enabled detailed sampling that continues to clarify the taxonomy and evolutionary relationships of rhodoliths and associated species. It has allowed the direct determination of bed distribution and abundance patterns, and field experiments to test hypotheses about the causes of these patterns. Such studies, including those in deep water that use technical diving, continue to provide new insights into bed structure and function. Careful collection and underwater experimentation have also revealed rhodoliths to be exceptional environmental recorders. Beds are, however, very susceptible to disturbance and climate change. Their conservation must be a priority if we are to continue to enjoy, better understand, and learn from these remarkable communities and the plants that dominate them. ACKNOWLEDGMENTS We thank the diving community for providing locations and in situ observations of new rhodolith beds that have contributed to understanding rhodolith distribution and provided new opportunities for research. Coauthor GAF thanks the Brazilian research agencies CNPq, CAPES, and FAPERJ for financial support for development projects. Coauthor NK is in receipt of a Royal Society of Edinburgh/Scottish Government Personal Research Fellowship (RSE 48704/1). This chapter is dedicated to the memory of Isabella A. Abbott (1919–2010), who contributed enormously to our knowledge of marine algae, championed international scientific collaboration, and always encouraged searches for subtidal seaweeds. REFERENCES Aburto-Oropeza, O., and E. F. Balart. 2001. Community structure of reef fish in several habitats of a rocky reef in the Gulf of California. PSZNI Marine Ecology, 22:283–305. Adey, W. H., T. Masaki, and H. Akioka. 1974. Ezo epiyessoense, a new parasitic genus and species of Corallinaceae (Rhodophyta, Cryptonemiales). Phycologia, 13:329–344. http://dx.doi.org/10.2216/i0031-8884-13-4-329.1. Adey, W. H., R. A. Townsend, and W. T. Boykins. 1982. The crustose coralline algae (Rhodophyta: Corallinaceae) of the Hawaiian Islands. Smithsonian Contributions to Marine Science, No. 15. Washinton, D.C.: Smithsonian Institution Scholarly Press. Aguirre, J., F. Perfectti, and J. C. Braga. 2010. Integrating phylogeny, molecular clocks and the fossil record in the evolution of coralline algae (Corallinales, Rhodophyta). Paleobiology, 36:519–533. http://dx.doi.org/10.1666/09041.1. n u mber 3 9 Alexandersson, T. 1974. Carbonate cementation in coralline algal nodules in Skagerrak, North Sea: Biochemical precipitation in undersaturated waters. Journal of Sedimentary Petrology, 44:7–26. Almeida, F. F. M. 1965. As ilhas oceânicas brasileiras e uma hipótese sobre a origem do Atlântico. Anais da Academia Brasileira Ciencias, 37:141–145. Amado-Filho, G. M., G. Maneveldt, R. C. C. Manso, B. V. Marins-Rosa, M. R. Pacheco, and S. Guimarães. 2007. Structure of rhodolith beds from 4 to 55 meters deep along the southern coast of Espírito Santo State, Brazil. Ciencias Marinas, 32:399–410. Amado-Filho, G. M., G. W. Maneveldt, G. H. Pereira-Filho, R. C. C. Manso, R. G. Bahia, M. B. Barros-Barreto, and S. M. P. B. Guimarães. 2010. Seaweed diversity associated with a Brazilian tropical rhodolith bed. Ciencias Marinas, 36(4):371–391. http://dx.doi.org/10.7773/cm.v36i4.1782. Bahia, R. G., R. Riosmena-Rodríguez, G. W. Maneveldt, and G. M. Amado-Filho. 2011. First report of Sporolithon ptychoides (Sporolithales, Corallinophycidae, Rhodophyta) for the Atlantic Ocean. Phycological Research, 59:64–69. http://dx.doi.org/10.1111/j.1440-1835.2010.00599.x. Ballesteros, E. 1988. Composición y estructura de los fondos de maërl de tossa de Mar (Gerona, España). Collectanea Botanica, 17:161–182. http://dx.doi.org /10.3989/collectbot.1989.v17.137. Ballesteros, E., and J. Afonso-Carrillo. 1995. Species records and distribution of shallow-water coralline algae in a western Indian Ocean coral reef (Trou d’Eau Douce, Mauritius). Botanica Marina, 38:203–213. http://dx.doi.org/10 .1515/botm.1995.38.1-6.203. Bassi, D., J. H. Nebelsick, A. Checconi, J. Hohenegger, and Y. Iryu. 2009. Present- day and fossil rhodolith pavements compared: Their potential for analyzing shallow-water carbonate deposits. Sedimentary Geology, 214:74–84. http:// dx.doi.org/10.1016/j.sedgeo.2008.03.010. Basso, D. 1998. Deep rhodolith distribution in the Pontian Islands, Italy: A model for the paleoecology of a temperate sea. Paleogeography, Paleoclimatology, Paleoecology, 137:173–187. http://dx.doi.org/10.1016/S0031-0182(97)00099-0. Birkett, D., C. Maggs, and M. Dring. 1998. Maërl, Volume V : An overview of dynamic and sensitivity characteristics for conservation management of marine SACs. Oban, UK: Scottish Association for Marine Science. Blake, C., and C. Maggs. 2003. Comparative growth rates and internal banding periodicity of maërl species (Corallinales, Rhodophyta) from northern Europe. Phycologia, 42:606–612. http://dx.doi.org/10.2216/i0031-8884-42-6-606.1. Bosellini, A., and R. N. Ginsburg. 1971. Form and internal structure of recent algal nodules (Rhodolites) from Bermuda. Journal of Geology, 79: 669–682. http:// dx.doi.org/10.1086/627697. Bosence, D. W. J. 1976. Ecological studies on two unattached coralline algae from western Ireland. Paleontology, 19:365–395. ———. 1979. Live and dead faunas from coralline algal gravels, Co. Galway. Paleontology, 22:449–478. ———. 1983. The occurrence and ecology of recent rhodoliths: A review. In Coated grains, ed. T. M. Peryt, pp. 225–242. Berlin: Springer-Verlag. http://dx.doi .org/10.1007/978-3-642-68869-0. Bridge, T. C. L., T. J. Done, R. J. Beaman, A. Friedman, S. B. Williams, O. Pizarro, and J. M. Webster. 2010. Topography, substratum and benthic macrofaunal relationships on a tropical mesophotic shelf margin, central Great Barrier Reef, Australia. Coral Reefs, doi:10.1007/s00338-010-0677-3. http://dx.doi .org/10.1007/s00338-010-0677-3. Burdett, H. L., N. A. Kamenos, and A. Law. 2011. Using coralline algae to understand historic marine cloud cover. Paleogeography, Paleoclimatology, Paleoecology, 302:65–70. http://dx.doi.org/10.1016/j.palaeo.2010.07.027. Cabioch, J. 1969. Les fonds de maërl de la Baie de Morlaix et leur peuplement vegetal. Cahiers de Biologie Marine, 9:139–161. Chave, K. E., and B. D. Wheeler. 1965. Mineralogic changes during growth in red algae, Clathromorphum compactum. Science, 147:621. http://dx.doi.org/10 .1126/science.147.3658.621. Clark, R. N. 2000. The chiton fauna of the Gulf of California rhodolith beds (with descriptions of four new species). Nemouria, 43:1–20. Darwin, C. R. 1844. Geological observations on the volcanic islands visited during the voyage of H.M.S. Beagle. London: Smith Elder and Co. Dawson, E. Y. 1960a. New records of marine algae from Pacific Mexico and Central America. Pacific Naturalist, 1:31–52. ———. 1960b. Marine red algae of Pacific Mexico. Part III. Cryptonemiales, Corallinaceae, subf. Melobesioideae. Pacific Naturalist, 2:3–125. De Grave, S. 1999. The influence of sedimentary heterogeneity on within maërl bed differences in infaunal crustacean community. Estuarine, Coastal and Shelf Science, 49:153–163. http://dx.doi.org/10.1006/ecss.1999.0484. • 153 Dias, G. T. M. 2000. Granulados bioclásticos – algas calcárias. Brazilian Journal of Geophysics, 18:307–318. Ehrhold, A., D. Hamon, and B. Buillaumont. 2006. The REBENT monitoring network, a spatially integrated, acoustic approach to surveying nearshore macrobenthic habitats: Application to the Bay of Concarneau (South Brittany, France). ICES Journal of Marine Science, 63:1604–1615. http://dx.doi .org/10.1016/j.icesjms.2006.06.010. Farias, J., R. Riosmena-Rodríguez, and P.A. Horta. 2010. Lithothamnion superpositum (Corallinales, Rhodophyta): First description for Brazil or a rediscovery of a species? Phycological Research, 51:210–216. http://dx.doi.org/10.1111/j .1440-1835.2010.00581.x. Figueiredo, M. A. D. O., K. Santos de Menezes, E. M. Costa-Paiva, P. C. Paiva, and C. R. R. Ventura. 2007. An experimental evaluation of rhodoliths as living substrata for infauna in the Abrolhos Bank, Brazil. Ciencias Marinas, 33: 427–440. Foslie, M. 1900. Die Systematik der Melobesieae. (Eine Berichtigung). Berichte der Deutschen Botanischen Gesellschaft, 18:239–241. Foster, M. S. 2001. Rhodoliths: Between rocks and soft places. Journal of Phycology, 37:659–667. http://dx.doi.org/10.1046/j.1529-8817.2001.00195.x. Foster, M. S., L. M. McConnico, L. Lundsten, T. Wadsworth, T. Kimball, L. B. Brooks, M. Medina-Lopez, R. Riosmena-Rodríguez, G. Hernandez-Carmona, R. Vasquez-Elisando, S. Johnson, and D. L. Steller. 2007. The diversity and natural history of a Lithothamnion muelleri–Sargassum horridum community in the Gulf of California. Ciencias Marinas, 33:367–384. Foster, M. S., R. Riosmena-Rodríguez, D. L. Steller, and W. J. Woelkerling. 1997. Living rhodolith beds in the Gulf of California and their implications for paleoenvironmental interpretation. In Pliocene carbonates and related facies flanking the Gulf of California, Baja California, Mexico, ed. M. E. Johnson and J. Ledesma-Vazquez, pp.127–139. Geological Society of America Special Paper 318. Boulder, Colo.: Geological Society of America. http://dx.doi.org/10 .1130/0-8137-2318-3.127. Frantz, B. R., M. S. Foster, and R. Riosmena-Rodríguez. 2005. Clathromorphum nereostratum (Corallinales, Rhodophyta): The oldest alga? Journal of Phycology, 41:770–773. http://dx.doi.org/10.1111/j.1529-8817.2005.00107.x. Frantz, B. R., M. Kashgarian, K. H. Coale, and M. S. Foster. 2000. Growth rate and potential climate record from a rhodolith using C-14 accelerator mass spectrometry. Limnology and Oceanography, 45:1773–1777. http://dx.doi .org/10.4319/lo.2000.45.8.1773. Freiwald, A., and R. Henrich. 1994. Reefal coralline algal build-ups within the Arctic circle: Morphology and sedimentary dynamics under extreme environmental seasonality. Sedimentology, 41:963–984. http://dx.doi.org/10.1111/j.1365 -3091.1994.tb01435.x. Gherardi, D. F. M. 2004. Community structure and carbonate production of a temperate rhodolith bank from Arvoredo Island, southern Brazil. Brazilian Journal of Oceanography, 52:207–224. Graham, M. H., B. P. Kinlan, L. D. Dreuhl, L. E. Garske, and S. Banks. 2007. Deep- water kelp refugia as potential hotspots of tropical marine diversity and productivity. Proceedings of the National Academy of Sciences, 104:16576– 16580. http://dx.doi.org/10.1073/pnas.0704778104. Grall, J., and M. Glemarec. 1997. Biodiversite des fonds de maërl en Bretagne: approache fonctionnelle et impacts anthropogeniques. VIE MILIEU, 47:339–349. Grall, J., and J. M. Hall-Spencer. 2003. Problems facing maërl conservation in Brittany. Aquatic Conservation: Marine and Freshwater Ecosystems, 13:55–64. http://dx.doi.org/10.1002/aqc.568. Grall, J., F. Le Loc’h, B. Guyonnet, and P. Riera. 2006. Community structure and food web based on stable isotopes (DELTA15N and DELTA13C) analysis of a North Eastern Atlantic maërl bed. Marine Ecology Progress Series, 338:1–15. Halfar, J., S. Hetzinger, W. H. Adey, T. Zack, G. Gamboa, B. Kunz, B. Williams, and D. E. Jacob. 2011. Coralline algal growth-increment widths archive North Atlantic climate variability. Paleogeography, Paleoclimatology, Paleoecology, 302:71–80. http://dx.doi.org/10.1016/j.palaeo.2010.04.009. Halfar, J., R. S. Steneck, M. Joachimski, A. Kronz, and A. D. Wanamaker. 2008. Coralline red algae as high-resolution climate recorders. Geology, 36:463–466. http://dx.doi.org/10.1130/G24635A.1. Halfar, J., R. Steneck, B. Schone, G. W. K. Moore, M. Joachimski, A. Kronz, J. Fietzke, and J. Estes. 2007. Coralline alga reveals first marine record of subarctic North Pacific climate change. Geophysical Research Letters, 34:L07702. http://dx.doi.org/10.1029/2006GL028811. Halfar, J., T. Zack, A. Kronz, and J. C. Zachos. 2000. Growth and high resolution paleoenvironmental signals of rhodoliths (coralline red algae): A new biogenic archive. Journal of Geophysical Research, C, Oceans, 105: 22,107– 22,116. http://dx.doi.org/10.1029/1999JC000128. 154 • smithso n i a n co n trib u tio n s to the m a ri n e scie n ces Hall-Spencer, J. M. 1998. Conservation issues relating to maërl beds as habitats for molluscs. Journal of Conchology, Special Publication, 2:271–286. Hall-Spencer, J. M., and R. J. A. Atkinson. 1999. Upogebia deltaura (Crustacea: Thalassinidea) in Clyde Sea maërl beds, Scotland. Journal of the Marine Biological Association of the United Kingdom, 79:871–880. http://dx.doi.org/10 .1017/S0025315498001039. Hall-Spencer, J. M., J. Grall, P. G. Moore, and R. J. A. Atkinson. 2003. Bivalve fishing and maërl-bed conservation in France and the UK: Retrospect and prospect. Aquatic Conservation: Marine and Freshwater Ecosystems, 13:S33– S41. http://dx.doi.org/10.1002/aqc.566. Hall-Spencer, J. M., and P. G. Moore. 2000. Scallop dredging has profound, long- term impacts on maërl habitats. ICES Journal of Marine Science, 57:1407– 1415. http://dx.doi.org/10.1006/jmsc.2000.0918. Hall-Spencer, J. M., N. White, E. Gillespie, K. Gillham, and A. Foggo. 2006. Impact of fish farms on maërl beds in strongly tidal areas. Marine Ecology Progress Series, 326:1–9. http://dx.doi.org/10.3354/meps326001. Harvey, A. S., and W. J. Woelkerling. 2007. A guide to nongeniculate coralline red algal (Corallinales, Rhodophyta) rhodolith identification. Ciencias Marinas, 33:411–426. Harvey, A. S., W. J. Woelkerling, T. Farr, K. Neill, and W. Nelson. 2005. Coralline Algae of Central New Zealand: An Identification Guide to Common ‘Crustose’ Species. Wellington, New Zealand: NIWA. Henrich, R., A. Freiwald, A. Wehrmann, P. Schafer, C. Samtleben and H. Zankl. 1996. Nordic cold water carbonates: Occurrence and controls. In Global and Regional Controls on Biogenic Sedimentation, ed. J. Reitner, F. Neuweiler, and F. Gunkel, pp. 35–53. Gottingen, Germany: Gottinger Arbeiten Geologie und Paläontologie. Hily, C., P. Potin, and J.-Y. Floc’h. 1992. Structure of subtidal algal assemblages on soft-bottom sediments: Fauna/flora interactions and role of disturbances in the Bay of Brest, France. Marine Ecology Progress Series, 85:115–130. http:// dx.doi.org/10.3354/meps085115. Hinojosa-Arango, G., and R. Riosmena-Rodríguez. 2004. Influence of rhodolith- forming species and growth form on associated fauna of rhodolith beds in the central west Gulf of California, Mexico. Marine Ecology, 25:109–127. http:// dx.doi.org/10.1111/j.1439-0485.2004.00019.x. Jackson, C. M., N. A. Kamenos, P. G. Moore, and M. Young. 2004. Meiofaunal bivalves in maërl and other substrata: Their diversity and community structure. Ophelia, 58:49–60. http://dx.doi.org/10.1080/00785236.2004.10410212. Jacquotte, R. 1962. Etude des fonds de maërl de Mediterranee. Recueil des Travaux de la Station Marine D’Endoume, Bulletin, 26:143–235. James, D. W. 2000. Diet, movement, and covering behavior of the sea urchin Toxopneustes roseus in rhodolith beds in the Gulf of California, Mexico. Marine Biology, 137:913–923. http://dx.doi.org/10.1007/s002270000423. James, D. W., M. S. Foster, and J. O’Sullivan. 2006. Bryoliths (Bryozoa) in the Gulf of California. Pacific Science, 60:117–124. http://dx.doi.org/10.1353/psc .2005.0057. Jokiel, P. L., K. S. Rodgers, I. B. Kuffner, A. J. Andersson, E. F. Cox, and F. T. Mackenzie. 2008. Ocean acidification and calcifying reef organisms: A mesocosm investigation. Coral Reefs, 27:473–483. http://dx.doi.org/10.1007/s00338-008 -0380-9. Kamenos, N. A. 2010. North Atlantic summers have warmed more than winters since 1353 and the response of marine zooplankton. Proceedings of the National Academy of Sciences, 107:22442–22447. http://dx.doi.org/10.1073 /pnas.1006141107. Kamenos, N. A., P. Calosi, and P. G. Moore. 2006. Substratum-mediated heart rate responses of an invertebrate to predation threat. Animal Behaviour, 71:809– 813. http://dx.doi.org/10.1016/j.anbehav.2005.05.026. Kamenos, N. A., M. Cusack, T. Huthwelker, P. Lagarde, and R. E. Scheibling. 2009. Mg-lattice associations in red coralline algae. Geochimica et Cosmochimica Acta, 73:1901–1907. http://dx.doi.org/10.1016/j.gca.2009.01.010. Kamenos, N. A., M. Cusack, and P. G. Moore. 2008. Red coralline algae are global paleothermometers with bi-weekly resolution. Geochimica et Cosmochimica Acta, 72:771–779. http://dx.doi.org/10.1016/j.gca.2007.11.019. Kamenos, N. A., and A. Law. 2010. Temperature controls on coralline algal skeletal growth. Journal of Phycology, 46:331–335. http://dx.doi.org/10.1111/j .1529-8817.2009.00780.x. Kamenos, N. A., P. G. Moore, and J. M. Hall-Spencer. 2004a. Nursery-area function of maërl grounds for juvenile queen scallops Aequipecten opercularis and other invertebrates. Marine Ecology Progress Series, 274:183–189. http://dx .doi.org/10.3354/meps274183. ———. 2004b. Attachment of the juvenile queen scallop (Aequipecten opercularis L.) to maërl in mesocosm conditions; juvenile habitat selection. Journal of Experimental Marine Biology and Ecology, 306:139–155. http://dx.doi.org /10.1016/j.jembe.2003.10.013. ———. 2004c. Maërl grounds provide both refuge and high growth potential for juvenile queen scallops (Aequipecten opercularis L.). Journal of Experimental Marine Biology and Ecology, 313:241–254. http://dx.doi.org/10.1016/j .jembe.2004.08.007. Keegan, B. F. 1974. The macrofauna of maërl substrates on the west coast of Ireland. Cahiers de Biologie Marine, 4:513–530. Keegan, B. F., and G. Konnecker. 1973. In situ quantitative sampling of benthic organisms. Helgolander Wissenschaftliche Meeresuntersuchungen, 24:256– 263. http://dx.doi.org/10.1007/BF01609516. Kempf, M. 1970. Notes on the benthic bionomy of the N-NE Brazilian Shelf. Marine Biology, 5: 213–224. http://dx.doi.org/10.1007/BF00346909. Kleypas, J. A., R. A. Feely, V. J. Fabry, C. Langdon, C. L. Sabine, and L. L. Robbins. 2006. Impacts of ocean acidification on coral reefs and other marine calcifers: A guide for future research. Seattle: NOAA/Pacific Marine Environmental Laboratory. Konar, B., R. Riosmena-Rodríguez, and K. Iken. 2006. Rhodolith bed: A newly discovered habitat in the North Pacific Ocean. Botanica Marina, 49:355–359. http://dx.doi.org/10.1515/BOT.2006.044. Kuffner, I. B., A. J. Andersson, P. L. Jokiel, K. S. Rodgers, and F. T. Mackenzie. 2007. Decreased abundance of crustose coralline algae due to ocean acidification. Nature Geoscience, 1:114–117. http://dx.doi.org/10.1038/ngeo100. Lavrado, H. P. 2006. Caracterização do ambiente e da comunidade bentônica. In Biodiversidade da costa central da zona econômica exclusiva brasileira, ed. H. P. Lavrado and B. L. Ignácio, pp. 19–66. Rio de Janeiro: Museu Nacional. Leão, Z. M., and R. N. Ginsburg. 1997. Living reefs surrounded by siliciclastic sediments: The Abrolhos coastal reefs, Bahia, Brazil. Proceedings of the eighth International Coral Reef Symposium, 2:1767–1772. Leliaert, F., C. Boedeker, V. Peña, F. Bunker, H. Verbruggen, and O. De Clerck. 2009. Cladophora rhodolithicola sp. Nov. (Cladophorales, Chlorophyta), a diminutive species from European maërl beds. European Journal of Phycology, 44:155–169. http://dx.doi.org/10.1080/09670260802573113. Lemoine, M. 1930. Les Corallinacées de l’archipel des Galapagos et du Golfe de Panama. Archives du Muséum National d’Histoire Naturelle, Paris, séries, 6, 4:37–88. Lieberman, M., D. M. John, and D. Lieberman. 1979. Ecology of subtidal algae on seasonally devastated cobble substrates off Ghana. Ecology, 60:1151–1161. http://dx.doi.org/10.2307/1936963. Littler, M. M. 1971. Tenarea tesselatum (Lemoine) Littler comb. nov., an unusual crustose coralline (Rhodophyceae, Cryptonemiales) from Hawaii. Phycologia, 10:355–359. http://dx.doi.org/10.2216/i0031-8884-10-4-355.1. Littler, M. M., D. S. Littler, S. M. Blair, and J. N. Norris. 1985. Deepest known plant life discovered on an uncharted seamount. Science, 227:57–59. http://dx.doi .org/10.1126/science.227.4682.57. Littler, M. M., D. S. Littler, and M. D. Hanisak. 1991. Deep-water rhodolith distribution, productivity, and growth history at sites of formation and subsequent degradation. Journal of Experimental Marine Biology and Ecology, 150:163– 182. http://dx.doi.org/10.1016/0022-0981(91)90066-6. Marrack, E. 1999. The relationship between water motion and living rhodolith beds in the southwestern Gulf of California, Mexico. Palaios, 14:159–171. http://dx.doi.org/10.2307/3515371. Milliman, J. D. 1977. Role of calcareous algae in Atlantic continental margin segmentation. In Fossil algae, ed. E. Flugel, pp. 232–247. Berlin: Springer-Verlag. http://dx.doi.org/10.1007/978-3-642-66516-5_26. Milliman, J. D., and C. A. B. Amaral. 1974. Economic potential of Brazilian continental margin sediments. Anais do Congresso Brasieiro de Geologia, 28:335– 344. Moberly, R. J. 1968. Composition of magnesian calcites of algal and pelecypods by electron microprobe analysis. Sedimentology, 11:61–82. http://dx.doi.org /10.1111/j.1365-3091.1968.tb00841.x. Norris, J. N. 2010. Marine algae of the Northern Gulf of California: Chlorophyta and Phaeophyceae. Smithsonian Contributions to Botany, No. 94. Washington, D.C.: Smithsonian Institution Scholarly Press. Peña, V., W. Adey, R. Riosmena- Rodríguez, H. G. Choi, J. Afonso- Carrillo, and I. Bárbara. 2011. Mesophyllum sphaericum sp. nov. (Corallinales, Rhodophyta): A new maërl-forming species from the NE Atlantic. Journal of Phycology, 47(4):911–927. http://dx.doi.org/10.1111/j.1529-8817.2011.01015.x. Peña, V., and I. Bárbara. 2010. Seasonal patterns in the maërl community of shallow European Atlantic beds and their use as baseline for monitoring studies. European Journal of Phycology, 45:327–342. http://dx.doi.org/10.1080 /09670261003586938. n u mber 3 9 Rahimpour- Bonab, H., Y. Bone, R. Moussavi- Harami, and K. Turnbull. 1997. Geochemical comparisons of modern cool-water calcareous biota, Lacepede Shelf, south Australia. Society for Sedimentary Geology, Special Publication, 56:77–92. Riosmena-Rodríguez, R., D. L. Steller, G. Hinojosa-Arango, and M. S. Foster. 2010. Reefs that rock and roll: Biology and conservation of rhodolith beds in the Gulf of California. In Marine biodiversity and conservation in the Gulf of California, ed. R. Brusca, pp. 49–71. Tucson, Ariz.: University of Arizona Press. Riosmena-Rodríguez, R., W. J. Woelkerling, and M. S. Foster. 1999. Taxonomic reassessment of rhodolith-forming species of Lithophyllum (Corallinales, Rhodophyta) in the Gulf of California, México. Phycologia, 38:401–417. http:// dx.doi.org/10.2216/i0031-8884-38-5-401.1. Riul, P., P. Lacouth, P. R. Pagliosa, M. L. Christoffersen, and P. A. Horta. 2009. Rhodolith beds at the easternmost extreme of South America: Community structure of an endangered environment. Aquatic Botany, 90:315–320. http:// dx.doi.org/10.1016/j.aquabot.2008.12.002. Schaeffer, T. N., G. J. Smith, M. S. Foster, and A. DeTomaso. 2002. Genetic differences between two growth-forms of Lithophyllum margaritae (Rhodophyta) in Baja California Sur, México. Journal of Phycology, 38:1090–1098. http:// dx.doi.org/10.1046/j.1529-8817.2002.01108.x. Schwarz, A. M., I. Hawes, N. Andrew, S. Mercer, V. Cummings, and S. Thrush. 2005. Primary production potential of non-geniculate coralline algae at Cape Evans, Ross Sea, Antarctica. Marine Ecology Progress Series, 294:131–140. http://dx.doi.org/10.3354/meps294131. Setchell, W. A. 1893. On the classification and geographical distribution of the Laminariaceae. Transactions of the Connecticut Academy of Arts and Sciences, 9:333–375. Steller, D. L., and C. Caceras-Martinez. 2009. Coralline algal rhodoliths enhance larval settlement and early growth of the Pacific calico scallop Argopecten ventricosus. Marine Ecology Progress Series, 396:49–60. http://dx.doi.org/10 .3354/meps08261. Steller, D. L., and M. S. Foster. 1995. Environmental factors influencing distribution and morphology of rhodoliths in Bahía Concepción, B.C.S., México. Journal of Experimental Marine Biology and Ecology, 194:201–212. http://dx.doi .org/10.1016/0022-0981(95)00086-0. Steller, D. L., M. S. Foster, and R. Riosmena- Rodríguez. 2007. Sampling and monitoring rhodolith beds. In Sampling biodiversity in coastal communi- • 155 ties: N aGISA protocols for seagrass and macroalgal habitats, ed. P. R. Rigby, K. Iken, and Y. Shirayama, pp. 93–97. Kyoto: Kyoto University Press. Steller, D. L., R. Riosmena-Rodriguez, M. S. Foster, and C. A. Roberts. 2003. Rhodolith bed diversity in the Gulf of California: The importance of rhodolith structure and consequences of disturbance. Aquatic Conservation: Marine and Freshwater Ecosystems, 13:S5–S20. http://dx.doi.org/10.1002/aqc.564. Thouzeau, G. 1991. Experimental collection of postlarvae of Pecten maximus (L.) and other benthic macrofaunal species in the Bay of Saint-Brieuc, France. I. Settlement patterns and biotic interactions among the species collected. Journal of Experimental Marine Biology and Ecology, 148:159–180. http:// dx.doi.org/10.1016/0022-0981(91)90080-G. Verheij, E. 1993. The genus Sporolithon (Sporolithaceae fam. nov., Corallinales, Rhodophyta) from the Spermonde Archipelago, Indonesia. Phycologia, 32: 184–196. http://dx.doi.org/10.2216/i0031-8884-32-3-184.1. Veron, J. E. N. 1995. Corals in space and time: The biogeography and evolution of the Scleractinia. Sydney: University of New South Whales Press. Villas-Boas, A. B., R. Riosmena-Rodríguez, G. M. Amado-Filho, G. W. Maneveldt, and M. Figueiredo. 2009. Taxonomy of rhodolith-forming species of Lithophyllum (Corallinales; Rhodophyta) from Espírito Santo State, Brazil. Phycologia, 48:237–248. http://dx.doi.org/10.2216/08-35.1. Weber-van Bosse, A. 1904. Corallineae verae of the Malay Archipelago. In The Corallinaceae of the Siboga Expedition, Siboga-Expeditie LXI, ed. A. Weber-van Bosse and M. Foslie, pp. 78–110. Leiden, Germany: E. J. Brill. Weber-van Bosse, A., and M. Foslie, eds. 1904. The Corallinaceae of the Siboga Expedition. Siboga-Expeditie LXI. Leiden, Germany: E. J. Brill. Woelkerling, W. J. 1988. The coralline red algae: An analysis of the genera and subfamilies of nongeniculate Corallinaceae. New York: Oxford University Press. Woelkerling, W. J., and M. S. Foster. 1989. A systematic and ecographic account of Synarthrophyton schielianum sp. nov. (Corallinaceae, Rhodophyta) from the Chatham Islands. Phycologia, 28:39–60. http://dx.doi.org/10.2216/i0031 -8884-28-1-39.1. Woelkerling, W. J, G. Gustavsen, H. E. Myklebost, T. Presto, and S. Såstad. 2005. The Coralline Red Algal Herbarium of Mikael Foslie: Revised Catalogue with Analyses. Trondheim, Norway: Museum of Natural History and Archaeology, Norwegian University of Science and Technology.