Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
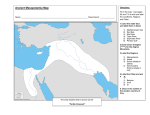
DISCUSSION Weight Fresh and dry weight data similar to that reported here were reported by Popp (1926) and Shirley plants in greenhouses glassed with Corning Glass filters trans- mitting separate spectral full sunlight house (1929), who grew many regions with equal energies. (289-720 nm) was more efficient duction of dry matter than was their skylight which gave "satisfactory" skylight and canopy shade treatments, weight were produced. no wavelengths However, spectral band permitted comparable concluded plants of nearly equal Shirleyrs canopy shade house had present. This more natural extended efficiency in weight production in photosynthesis of the HIR, or High Irradiance will be considered Reaction Shirley was a result of under various spectra; conclusion. The role (Mohr, 1962) in growth in later discussion. During the pre-emergence stage of seed germination, major energy source is endosperm-stored independence synthesizing plants to have dry to plants under skylight conditions. data from this research reinforce Shirleyts Seedling (529- In our corresponding canopy shade-grown that the decrease the decreased (374-585 nm) house, shorter than 529 nm, while our canopy shade treat- ments had blue wavelengths weights for the pro- results, or their canopy shade 720 nm) house, which gave poor results. Their seedlings materials the (Friend, 1966). is assumed when the dry weight of photo(minus seed coats) is equal to the initial 50 weight of the embryo all treatments and endosperm. At week 3, plants from had just passed this weight of 0.44 mg per Golf- rood red fescue seed (Fig. 5). Although germination at the same time in all treatments, lation had greater dry weights dence slightly occurred plants under the sun simu- and probably reached indepen- sooner than those under skylight and canopy shade treatments. Plants grown in sun-simulated weights and had a higher growth rate even at 3 weeks. growth rate may indicate photosynthesis, running that physiological transpiration, respiration) are are better integrated plants do rate at weeks 3 and 4, and lower points and respiration plants from other treatments, organized (i.e., Since sun simulation-grown have a higher photosynthetic light compensation A higher processes translocation, faster or that these processes and hence more efficient. better spectra had higher dry rates compared to we conclude that the processes are and hence the plants are more physiologically mature. Fresh Weight/Dry Higher reported ings. Ratio fresh weight/dry weight ratios in shade leaves were as early as 1917 by Hanson. (1929) confirmed ditions. Weight Popp (1926) and Shirley this finding under more controlled Data presented Hanson concluded in this report substantiate growth conthese find- that the lower fresh weight/dry weight ratios in sun leaves showed that more dry matter was laid down 51 in sun leaves. His sun leaves had 2 layers of compact palisade cells, compared to a single palisade rangement in shade leaves. The higher fresh weight/dry ratio of the fescue is probably tercellular layer and a loose cell ar- a result of larger and more in- spaces holding more water. useful under conditions This would be particularly of soil moisture stress where sun leaves, by virtue of having a higher osmotic pressure experience stomatal 2 than shaded leaves. plants have a lower photosynthetic synthesis reserves Although shade compensate for the re- Since sun plants usually have higher carbohydrate (Daubenmire, 1959), the sun environment logically more desirable, survival uptake at high- rate, the time of active photo- may be longer and may partially duced rate. (Marsh, 1941), may closure and hence reduced CO er soil water potentials weight but mechanisms may be physio- appear to exist allowing and growth under less than optimum conditions. Leaves Mitchell (l953b) studied leaf initiation in ryegrass and found that no new leaf primordia were initiated in darkness the rate of leaf appearance was proportional thetic activities of the plants. to the photosyn- In red fescue, however, plants grown in the skylight or in the canopy shade simulations same photosynthetic the skylight rates. simulation, and had the There were more leaves per plant in the individual in the canopy shade simulation, leaf weight was greater and total leaf biomass (essen- tially total plant weight) was the same under these two spectra. 52 As seen in Figs. 2 and 3, the skylight spectrum was proportionally higher in blue wavelengths than was the canopy shade spec- truro and also had higher red/far-red lations contained The work of Mohr that both the classical in regulating Tentatively, (1972) supports the hy- red/far-red system and the blue High Irradiance involved Canopy shade simu- lower absolute radiant fluxes in the blue re- gion of the spectrum. pothesis ratios. phytochrome Reaction leaf initiation reaction (HIR) system are and leaf development. it would appear that leaf number is more likely to be under the control of the HIR, blue system than the red/far-red phytochrome system because the high red/far-red ratio in the skylight would tend to negate the formative effects of inducing in the skylight red radiation spectrum (Kasperbauer, weight may be more influenced canopy shade. Leaf biomass, the result of interactions red systems. Obviously, and the blue HIR flux is high 1971). Individual leaf by the relatively high red in on the other hand, appears to be between both the HIR and the red/far- such conclusions are highly speculative and tentative. Leaf length was higher in all simulated radiation than under full sunlight with the exception treatment. in beans. and red light inhibited, that far- stem elongation In full sunlight the far-red is counterbalanced the red component, lectively of chamber skylight Downs et ale (1957) and others demonstrated red light stimulated, spectra by but under conditions where red light is se- reduced while far-red is less modified (as in the 53 simulated canopy shade treatments) be expected. increased elongation would This is supported by the observation that in the growth chamber, the sun treatment had a greater ratio of red to far-red light than did the skylight treatment, leaves were longer. treatments Similarly, transmitted the greenhouse and the sun sun and skylight nearly the same red to far-red ratio and the plants showed comparable leaf lengths. Tillering The tillering response also appears to be controlled by the availability of photosynthate (Mitchell, 1953a). Leopold and by other factors (1949) showed that tillering ley and teosinte is controlled by auxin. ported that increased ler production Although both Friend light intensity enhanced in balance with increased in bar- (1966) re- the rate of til- leaf production. fescue seedlings were used and not mature plants, sun-simulation plants in the growth chamber had approximately 4 leaves before tillering while those in the sun-simulation house had only 3 leaves before tillering. expanded lar environment Ryle (1964) showed that for a particu- and species, the number of leaves which are actively growing and contributing photosynthate (1964) studied distribution of Lolium perenne panded At least 2 fully- leaves were required before any seedling tillered. In related studies, Williams green- and Phleum pratense leaves steadily contributed is constant. of assimilates in leaves and found that fully ex- less photosynthate to the 54 rest of the plant until at senescence Maximum photosynthate auxin distribution expanded, probable and minimum they were unessential. auxin production and maximum would occur when new leaves are almost fully but before old leaves have begun to abscise. that spectral and/or distribution control of growth regulator synthesis would operate in tillering. leaf area may control tiller development ergy substrate the spectral It is and growth regulator flux and quality Leaf number or by adjusting both en- concentrations available. relative to New plants (tillers) will be fonned only under conditions where the parent plant is well established and able to compete with other plants for avail- able radiation. stomates Spectral effects Mansfield on stomata were reviewed by Meidner (1968), who discussed briefly the occurrance mates under spectral shade and also the physiology induced stomatal movement. more effective and of sto- of wavelength- Blue light was approximately 7 times in causing full stomate opening than was red light. There seems to be limited data on spectral intensity or quality on stomatal action or development, environmental ferentiation entiation influenced conditions existing in the embryonic (Zucker, 1963). at the time of stomatal dif- leaf bud greatly affect differ- Zucker found that both lAA and kinetin one or both divisions gested that gibberellin but it is known that in stomate formation, might also be involved. and sug- He concluded 55 that there is no absolute ment because plying light requirement they are differentiated for stomata develop- in complete darkness, im- that any light effect is secondary. If light plays only a secondary ergy substrate synthate and growth regulator is expected in diffuse regulator skylight and canopy shade-grown ment had the least. treatments synthetic regulation plants and least plants. Growth or a balance of these from plants in both the and the growth chamber were of equal size, but difplants had the highest of the three treatments, therefore, or kinetin, In fescue., stomates fuse skylight-grown skylight Most photo- are less certain because it is not known if auxin, gibberellin, greenhouse concentrations. in sun simulation-grown concentrations is regulatory. role, it may control en- and plants from the canopy shade treat- The plants under canopy shade and diffuse had nearly equal rates of photosynthesis stomate size was not determined process. density of stomates The light-influenced only by the photo- mechanism of stomate is not clear, but it could be influenced action involving phytochrome and and the High Irradiance by a similar Reaction. Photopigments with the exception of skylight simulation in the growth chamber, no significant differences were noted in either chloro- phylls or carotenoids. This exception in the Results section. to synthesize to the rule has been noted We can only conclude that the capacity photosynthetic pigments is independent of the 56 tested spectra and one would not expect chlorophyll ing under natural Kramer, Oosting shaded conditions. and Korstian be at a selective be synthesized. adequate which at a more rapid rate than they could and maintenance serve a photo-protective tations on photosynthesis The synthesis permitted (Fig. 15) (Krinsky, 1968), limi- due to alterations in chlorophyll con- role in the economy of the plant. of the photopigments specific wavelengths major activation of carotenoids function do not play a major shaded or non-shaded chlorophyll, in full sunlight if their chloro- Since all of the spectral simulations synthesis tent probably This point was stressed by (1952) who stated that plants might disadvantage phylls were photo-oxidized to be limit- is under the control of of visible and near-visible of chlorophyll at the absorption i.e., in both the blue and the red. radiation, with peaks of protoBlue wave- lengths are low in canopy shade relative to sunlight or skylight, but the red end of the visible spectrum is relatively high. The reverse is true for the skylight spectrum. Since either red or blue can activate chlorophyll the tested shade spectra would not limit chlorophyll synthesis, production greatly. In red fescue, the plants may be considered to be genetically pre-adapted to permit chlorophyll diant fluxes, which, full sunlight. periments synthesis even under low ra- under our conditions, were 5% and 10% of This is not true for all plants; indeed, ex- from many laboratories have shown that neutral shade at 10% or even 25% of full sunlight results in plants that are 57 pale green, and which were assumed to be low in chlorophyll. However as discussed in the visual observations canopy shade plants section, also appeared pale green but contained the same amount of chlorophyll on a dry weight basis. In addition, the canopy shade and skylight plants had equal weights canopy shade plants had significantly Therefore the but the fewer but longer leaves. it is assumed that the canopy shade plants had less chlorophyll per cell, although the cells were larger. Energy Metabolism In these studies, it was noted fairly constant decrease iod of measurement (Fig. 16) that there was a in respiratory with a threefold intensity over the per- drop over the three week period. In both the growth chamber and in the greenhouse lations, the respiratory simulations ditions. intensities of the plants in full sun were lower than for either canopy or skylight Since respiration biosynthetic the decreased purposes simu- con- provides ATP and reducing power for and carbon chains for such biosyntheses, respiratory intensities suggest that the develop- ment of new biomass had passed its peak and that respiratory energy demand. and precursors for biosynthesis was no longer in high The fact that the respiratory intensity of the sun- grown plants was always lower than for canopy-grown grown plants or skylight- suggests that the sun-grown plants were, time measurements from the were started, more mature. The production of photosynthate, as measured by CO2 fixation 58 appeared to be fairly constant in plants grown under sun-simu- lation conditions. decreased These data, together with the data on the rate of respiration over the same time period suggest that in plants grown under sun-simulation be an accumulation on leaf weights of dry matter. and leaf number in sun-simulation This formulation that were measured a somewhat maturity showed fairly constant skylight point (Fig. 17). and canopy shade conditions, different pattern was noted. tory intensities In both sets, respira- decreased with time, indicating of the plants, but the rates of CO 2 creases over the period of measurement. perrnitting the accumulation necessary for the synthesis of cell components. in production showed in- We consider this to an adaptation increase increased fixation represent adaptive conditions can also be seen in the changes in the compensation Under the simulated there will This is borne out by the data (Figs. 5 and 8), where the leaf biomass increments. conditions of materials However, of photosynthate the did not reach a level where growth (dry weight, etc.) caught up with the plants under sun-simulation conditions. to whether, The question had the study been continued to five weeks, still remains as for an additional the plants under canopy shade and skylight four con- ditions would have reached the level of maturation seen in 6 weeks (Fig. 9) for full sun-grown plants. the trends in both respiration sation points a physiological and photosynthesis Nevertheless, and the converging (Fig. 17) suggest that the catching-~p adaptation, was well under way within compen- process, 5 weeks. 59 In view of the photosynthetic biochemical Obviously, basis for the findings the adaptation photopigments, nor to 'structural modifications One must look, therefore, Bensen cycle enzymes. a Calvin-Bensen Bjorkman in stomate num- (1968a) measured the activity of enzyme, carboxydismutase carboxylase), higher plants grown in habitats in the at the activity of the Calvin- cycle photosynthetic (ribulose-I, diphosphate from leaf extracts The comparatively in the shade plants indicated zymes other than carboxydismutase that low carboxydismutase capacity difference synthesis plants in weak light He concluded in shade plants. capacity cannot explain of utilization of the photosynthetic process. the This was commonly thought to (Daubenmire, 1959). probably How- of weak light in photo- for high photosynthetic that shade adaptation Practical that their content of en- photosynthesis (Bjorkman, 1968b). have been an explanation low content of soluble may also be low. in light-capturing in efficiency com- activity is a factor that limits the for light-saturated ever, differences of of various light intensities. sun plants had higher levels of enzyme activity pared to shade plants. protein requires some evaluation_ is not due to an alteration bers. Ecotypic adaptation noted above, the rates of shade Bjorkman concluded involves several component steps Applications The importance of these formulations in shade adaptation is unknown, but results of this study have practical application 60 in regard to densities planted, the practice is frequently of cultivated proper plant densities will cause the many seedlings an optimum number of seedlings is that "seeding heavy" in that area. receiving receive virtually trum, resulting in poor leaf initiation. where canopies overstory ing provides Thus, instead of a complete spectrum, a canopy shade spec- are not extremely short periods A justification in an area to compete for and also the light environment the crowded seedlings When seed is of "seeding heavy to insure a good stand" used with no apparent rationale. for maintaining to modify turfgrass. In natural situations dense, and sun fleck- of complete sun spectra, excessively high seedling densities with resultant mutual shading might negate the beneficial effects of the sun flecks.