Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
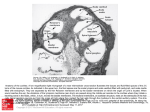
Hearing In terms of performance, hearing is the most remarkable of our senses The sense of hearing Frequency analysis: responds selectively to frequencies in range 20!20,000 Hz Tom DUKE Cavendish Laboratory, University of Cambridge Sensitivity: faintest audible sounds impart no more energy than thermal noise: 4 zJ per cycle Dynamic range: responds and adapts over 12 orders of magnitude of energy: 0!120 dB PHYSBIO 2007 St Etienne de Tinée Dynamic range 20µPa 0dB 20Pa 30dB whisper The ear 60dB conversation 90dB motorcycle auditory nerve 120dB pneumatic drill rifle displacement of air molecule at threshold of hearing ~ 0.1nm external middle inner Middle ear Cochlea . Impedance matches the air-filled ear canal with the fluid-filled cochlea Provides an acoustic gain owing to different areas of eardrum and stapes footplate basilar membrane Cochlea Organ of Corti Theories of hearing Theories of hearing Helmholtz 1857 von Békésy 1923-47 Detection apparatus consists of a set of strings of differing length which vibrate in resonance with the incoming sound Careful experiments on cadavers showed that a wave travels along the basilar membrane, and reaches a peak amplitude at a position that depends on frequency Notion of Theories of hearing Gold 1948 Argued that damping by the fluid in the cochlea would not permit resonant oscillation and proposed that the ear is powered Gold’s regeneration hypothesis place code Gold’s regeneration hypothesis Regeneration hypothesis: Otoacoustic emissions The ear in not just a sound receiver; it also spontaneously emits sounds at a very low level An energy supply provides a feedback proportional to velocity, and in phase with it Critical point near critical point, sharply tuned response at frequency ‘!The magnitude of the feedback we require is so large as to come precariously close to cancelling the resistive losses. Some sort of self-regulating device would have to exist!’ Hair cells in the bullfrog sacculus Kemp ‘79; Manley & Köppl ‘98 Detection apparatus Hair bundle is composed of ~50 ‘stereocilia’, which lean against each other. Each sterocilium is a bundle of actin filaments, surrounded by the cell membrane, which tapers at the base. Adjacent stereocilia are connected by a fine filament — the ‘tip link’ stereocilia transduction channel tip link + K kinocilium (absent in mammals) tip links source: Hudspeth Detection apparatus Hair bundle is composed of ~50 ‘stereocilia’, which lean against each other. Each sterocilium is a bundle of actin filaments, surrounded by the cell membrane, which tapers at the base. Adjacent stereocilia are connected by a fine filament — the ‘tip link’ Mechano-chemo-electrical transduction When bundle is pushed in direction of tallest stereocilium, increased tension in tip links pulls open transduction channels & admits K+ which depolarizes the membrane & opens voltage-gated channels at the base of the cell to nerve synapse tip link Rapidity of transduction process preserves information about timing of the signal Transduction current Active detetion by hair bundles Active bundle movement Spontaneous hair-bundle oscillations Crawford & Fettiplace ‘86; Howard & Hudspeth ‘87; Benser, Marquis & Hudspeth ‘96 In the correct physiological conditions, hair bundles actively oscillate spontaneous oscillation ? twitch Self-tuned critical oscillators Building a critical oscillator: inertial system Camalet, Duke, Jülicher & Prost ‘00 Gold ‘48 Vibration sensor is a nonlinear mechanical system which can generate self-sustained oscillations at a characteristic frequency internal active force amplitude A feedback control mechanism maintains it on the verge of oscillating Hopf bifurcation critical point: characteristic frequency: control parameter remarkable response properties at critical point Building a critical oscillator: non-inertial system An internal active process can cause a heavily damped mechanical system to oscillate Hopf resonance force: displacement: active force with its own dynamics, coupled to displacement control parameter: C bifurcation point: Suppose effective elasticity can be made negative by changing control parameter C • stimulus at characteristic frequency: Critical point at gain diverges for weak stimuli Characteristic frequency Critical Hopf resonance: single tone response Hopf resonance force: Gain and active bandwidth depend on level of stimulus displacement: gain control parameter: C • 0 db bifurcation point: ! fa stimulus at different frequency: 20 db if active bandwidth fc frequency Critical Hopf resonance: response in presence of noise Canonical equation Eguiluz et al. ‘00 Camalet, Duke, Jülicher & Prost ‘00 phase-locking Response at ! = !c for different f r ! /!c Self-adjustment to critical point Self-tuned critical oscillations Slow dynamics of control parameter, coupled to displacement, provides negative feedback which automatically adjusts system to the critical point x " C Cc C Eg. Ca2+ concentration is control parameter stimulus Spontaneous fluctuations Hair bundle response Martin, Hudspeth & Julicher ‘01 Auto-correlation function Martin, Julicher & Hudspeth ‘01 Response of a frog hair bundle forced by a microneedle Linear response Test of fluctuation-dissipation relation Martin, Hudspeth & Julicher ‘01 Response function # defined by: Martin, Hudspeth & Julicher ‘01 At thermal equilibrium, the linear response is related to the fluctuations Hair bundle response Two adaptation mechanisms Martin & Hudspeth ‘01 -2/3 fast process slow process Ca2+ binding to transduction channel movement of myosin-1C motors ~ 1 ms ~ 100 ms Negative elasticity Channel gating compliance Martin & Hudspeth ’00 Instantaneous force response to an applied displacement Suppose channel incorporates a lever arm opening of channel can substantially reduce the tension in the tip link • negative elasticity if Channel gating compliance Influence of motors Physical basis of oscillations channels closed channels open Ca2+ released from motors; causes them to produce more force Ca2+ binds to motors; causes them to produce less force Self-tuning mechanism ? Tinévez & Martin ‘06 Tinévez & Martin ‘06 Maybe Ca2+ affects rigidity of tip-links, or size of channel movement force/displacement bundle position motor position transduction current Alternative physical basis of self-tuned oscillations Alternative physical basis of self-tuned oscillations Vilfan & Duke ‘02 • Oscillations generated by interaction of Ca2+ with transduction channels Vilfan & Duke ‘02 • Oscillations generated by interaction of Ca2+ with transduction channels motors Ca2+ frequency depends on bundle geometry Ca Ca • An active role for the kinocilium ? • kinocilium is motile (Rüsch & Thurm ‘90) + K kinocilium tip link • axoneme can vibrate at ~1000 Hz (Kamimura & Kamiya ‘89) • but spontaneous oscillations of frog hairbundles still occur when the bundle is knocked down (Martin) detection apparatus Hearing in fruit flies The antenna of mosquitoes sense both odorants and sounds stereocilia transduction channel Self-tuning accomplished by movement of molecular motors, regulated by Ca2+ force generator Spontaneous oscillations and nonlinear response of antennae level (dB) Göpfert ‘03 Köppl & Manley ‘93 Spontaneous otoacoustic emissions in reptiles frequency (Hz) Reptilian inner ear Coupled critical oscillators Köppl & Manley ‘93 . Vilfan & Duke bobtail lizard mi-1 mi mi+1 sallet: massive structure !i-1 !i !i+1 real for dissipative coupling Spontaneous oscillations of a coupled chain oscillation frequency Cochlear mechanics Ermentrout & Koppel ‘84 !i Human cochlea Békésy’s travelling wave . Incoming sound excites a slow travelling wave on the basilar membrane cochlea tectoral membrane basilar membrane inner hair cells outer hair cells • location of peak depends on frequency Place code Cochlear travelling wave Travelling wave: one-dimensional model Zwislocki ‘48 membrane displacement h oval window pressure difference difference in flows round window p = P1 - P2 j = J1 - J2 helicotrema • • fluid flow • incompressibility • membrane response sound sets fluid into motion • variation in flow rate is accommodated by movement of membrane • membrane acceleration is caused by difference in fluid pressure wave velocity Travelling wave: damped dispersive waves decreases by two orders of magnitude from base to apex as wave propagates, it slows down & its amplitude increases Wave peaks when damping becomes significant Location of peak depends on frequency 4.6 kHz Sound level required to see an observable response at a given place level (dB) Stiffness Tuning curve 1.3 kHz 370 Hz frequency (kHz) Tuning curve Energy flow in the cochlea Lighthill ‘81 In order to obtain a very localized peak, require: • only waves with frequency lower than !c(x) carry energy past point x • as wave approaches characteristic place, velocity of energy transport falls to zero • velocity drops so fast that there is time to dissipate all energy before wave reaches characteristic place energy piles up at characteristic place Outer hair cells Active cochlear partition Kachar et al. ‘86; Ashmore ‘87 Critical oscillators are ranged along basilar membrane, and are positioned so that they can drive its motion • characteristic frequency diminishes from base to apex, spanning the audible range Partition is an excitable medium with a nonlinear active response inner hair cells outer hair cells • active oscillators negate friction and damping at the characteristic place where the oscillator frequency matches the sound frequency Active travelling wave: waveforms Basilar membrane motion Rhode ‘71; Ruggero et al. ‘97; Russell & Nilsen ‘97 BM response is nonlinear 4.6 kHz 1.3 kHz 370 Hz Basilar membrane motion Energy flow in the cochlea Rhode ‘71; Ruggero et al. ‘97; Russell & Nilsen ‘97 Lighthill ‘81 Cochlea is an unusual type of waveguide: BM response is nonlinear 1/3 1 • only waves with frequency lower than !c(x) carry energy past point x • as wave approaches characteristic place, velocity of energy transport falls to zero • velocity drops so fast that there is time to dissipate all energy before wave reaches characteristic place energy piles up at characteristic place Nonlinearities due to active amplification A critical oscillator is ideal for detecting a single tone … … but it causes tones of different frequency to interfere Psychoacoustic effects & their physical origin Response to multiple tones: Two-tone suppression Two-tone suppression f1 Psychoacoustics & auditory illusions Range of phenomena associated with multiple tones: f1, f2, ... Combination tones: presence of a second tone diminishes the neural response to the first tone Tartini 1714 frequencies not present in the stimulus can be heard, most prominently 2f1 - f2 f2 Residue pitch: Schouten 1938 frequencies close to, but not necessarily equal to "f = f2 - f1 can be heard f1 & f2 suppression is greater if frequencies are close Consonance: Pythagoras two musical notes whose frequencies are in a simple integer ratio sound pleasant basis of diatonic scale Two-tone suppression Distortion products Presence of second tone can extinguish the nonlinear amplification =0 Nonlinearities create a characteristic spectrum of distortion products ! 0 Active travelling wave: two-tone response Active travelling wave: two-tone response Andor !2 !1 2 ! 1- !2 2!1-!2 !1 !2 distance along cochlea frequency component location vibrating almost exclusively at 2!1-!2 distortion product can be heard Neural representation of pitch and level Auditory nerve: spontaneous activity Liberman ‘77 Two competing theories: pitch represented by the place in the cochlea where nerves fire most frequently Time code pitch represented by time interval between spikes threshold (dB) level represented by firing rate at that place number of units Place code level represented by number of nerves firing at an elevated rate spontaneous rate (spikes/sec) Inference of pitch and level First passage time to traversal of threshold Jülicher, Andor & Duke ‘01 Model • a spike is elicited whenever the hair bundle deflection traverses a threshold • there are hair cells with a wide range of thresholds for each characteristic frequency Algorithm • • construct histogram of inter-spike intervals T, summing over all hair cells in the cochlea • perceived tones correspond to peaks in histogram and are assigned pitch 1/T perceived level corresponds to height of peak spontaneous rate (spikes/sec) Temporal coding permits detection of stimuli 20 dB below rate-threshold First passage time to traversal of threshold Temporal coding permits detection of stimuli 20 dB below rate-threshold Distortion products: spectra & waveforms spontaneous high level stimulus Spike timings: two tones Two-tone stochastic resonance Chialvo et al. ‘02 . firing prob. density high threshold neuron interval / s 0.009s ~ 110 Hz 0.045s ~ 22 Hz residue pitch Dissonance: psychophysical experiments Nature of dissonance Kaestner 1909 Helmholtz ascribed dissonance to close mis-matches in frequency between harmonics 5:4 Integer ratios of frequencies are preferred when notes are played on musical instruments 4:3 For pure tones, there is no such preference, but only a dislike of close frequencies 1:1 8:5 6:5 4:3 3:2 5:3 5:4 0 0.2 2:1 1 pleasantness 0.8 " 2 :1 0.6 0.4 0.2 pure tones + harmonics 0 Two tones can be clearly distinguished if they differ in frequency by more than 15% Close frequency mis-matches generate complicated hair bundle responses f2 / f1 loudness 1.03 0.8 time 1.10 The difficulty of inferring frequency components from partial information about a complex waveform results in an indeterminacy of pitch 1.3 rough inferred pitch Two tones are interpreted ambiguously if they differ in frequency by 5 - 15% 0.6 Dissonance: a physical interpretation Pitch discrimination Two tones are heard as a single tone if they differ in frequency by less than 5% 0.4 !f / f log(frequency) 1.2 1.1 1 1.30 1 1.1 1.2 f2 / f1 0.8 1 1.2 1.4 inferred pitch T -1/ f1 ‘What distinguishes dissonances from consonances is not a greater or lesser degree of beauty, but a greater or lesser degree of comprehensibility’ Arnold Schoenberg 1.3 1