Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
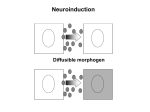
Bio 3411, Fall 2006 Aguan Wei 979 McDonnell Sciences Building, Medical School Campus, WUMS. email: [email protected] Lecture 18 – Molecular Mechanisms of Neural Induction. 1. During development, embryonic cells signal to each other with secreted diffusible molecules that instruct neighboring cells to change their pattern of gene expression. This signaling event is termed induction. Because the signaling molecule instructs the developmental fate of the recipient cell, it is called a morphogen. 2. Induction is observed in classic experiments manipulating amphibian blastula: a. (1924) Hilde Mangold and Hans Spemann. Transplanting the dorsal lip of the blastopore (presumptive mesoderm) from one embryo (in late blastula stage) to the ventral pole of a second embryo (same late blastula stage) results in an embryo with a second neuroaxis! The transplanted tissue is capable of “organizing” a second neuroaxis, so this region of the embryo is termed the “Spemann-Mangold Organizer”. b. (1969) Pieter Nieuwkoop, (1989) Grunz and Tacke, (1991) Godsave and Slack. Isolated animal caps (neuroectodermal cells) from blastulas can be cultured in vitro. Unlike the embryo, cultured intact animal caps develop into epidermal cells, instead of neurons. However, dissociated animal cap cells develop into neurons. Close contact between developing neuroectodermal cells results in expression of epidermal cell fate, whereas loss of close contact results in neuronal cell fate. c. These two classic experiments form the basis for the molecular analysis of neural induction. 3. Experimental manipulations show that inductive signaling occurs between neighboring cells of the developing neuroectoderm, and between developing neuroectoderm and mesoderm to generate the neural plate. Normal morphogenic movements during gastrulation permit induction to occur, by placing signaling and recipient tissues together in close apposition. 4. Developing neuroectodermal cells secrete a protein which signals neighboring cells to inhibit the neural fate and promote the epidermal fate. This signaling protein is Bone Morphogenic Protein-4 (BMP-4), a member of the Transforming Growth Factorβ (TGF-β) family of proteins. Experimental evidence; (1990-1994) Ali Hemmati-Brivanlou and Doug Melton: a) Expression of exogenous dominant-negative TGF-β receptors, blocks the function of native TGF-β receptors, leading to neuronal differentiation by intact animal caps. b) BMP-4 is expressed in neuroectodermal cells. c) Addition of BMP-4 to cultures of dissociated animal caps, causes differentiation of epidermal cells, due to repression of the neural fate. 1 5. Developing mesodermal cells secrete multiple protein factors that antagonize the action of BMP-4. Three main antagonists of BMP-4 are chordin, noggin and follistatin. All antagonist factors are: a) Expressed in the Spemann-Mangold organizer and in developing mesoderm at the right time to act in neuroinduction. b) Capable of inducing a second neural axis in embryos, similar to a transplanted Spemann-Mangold organizer. c) Capable of inducing the neuronal fate in cultures of intact animal caps. 6. Identification of chordin and noggin relied upon clever molecular cloning strategies. A) (1992) William Smith and Richard Harland. A functional expression cloning strategy yields noggin. a) Ventralized blastula (no neural plate develop) were produced en masse, by treating blastulas with ultraviolet irradiation. Ventralized blastula used to assay for candidate factors promoting neural plate formation. b) mRNAs expressed by blastula were converted to cDNAs, then cloned into a bacterial plasmid vector to generate a representative cDNA library. c) Pools of cDNAs clones were transcribed into cRNA, in vitro, then injected into ventralized blastula. d) Each positive pool of cDNAs (capable of inducing a neural plate in ventralized blastula) was fractionated and re-assayed. e) Reiterative rounds of fractionation and screening for positive pools of cDNAs, with each successive positive pool containing fewer numbers of clones, resulted ultimately in the isolation of a single clone capable of full neuralizing activity. 10 pools of ~10,000 clones screened, yielding one clone for noggin. B) (1994) Yoshiki Sasai and Eddy De Robertis. A differential screening strategy yields chordin. a) An embryonic cDNA library was plated in duplicate, then screened with probes made from mRNAs enriched for “dorsalized” (expanded Spemann organizer, with LiCl treatment) or “ventralized” (no Spemann organizer, with UV treatment) embryos. Clones isolated which were positive for “dorsalized” probe and negative for “ventralized” probe. ~25,000 clones screened, yielding 6 clones. b) Positive clones further assayed for ability to induce neural axis in ventralized blastula and for appropriate pattern of tissue expression by in situ hybridization. c) Chordin identified from 3 of the 6 candidate clones. Sequence of chordin reveals evolutionary conservation with Drosophila short gastrulation (sog), which functions in Drosophila neuronal induction. 2 7. Chordin/Noggin/Follistatin antagonizes BMP-4 activity by directly binding and inactivating BMP-4. Evidence provided by in vitro protein association assays. 8. Molecular model of neural induction: a) The “default” fate of neuroectodermal cells is neuronal. b) BMP-4 secreted by neuroectodermal cells inhibits neuronal fate and promotes epidermal fate. This inductive signal between neighboring cells acts to promote neuroectodermal cells towards the epidermal fate, and maintains the epidermal fate in intact cultured animal caps. c) Antagonists of BMP-4 (Chordin, Noggin and Follistatin) are secreted by mesoderm, which underlies developing neuroectoderm in the gastrula. d) Chordin/Noggin/Follistatin diffuse to overlying neuroectoderm, and inactivate BMP-4 activity by directly binding BMP-4. e) Inactivation of BMP-4 activity in the developing neuroectoderm, releases repression of neuronal fate, allowing expression of “default” neuronal fate and formation of the neural plate. Loss of BMP-4 activity is mimicked in dispersed neuroectodermal cultures by dilution of secreted BMP-4 in vitro, resulting in expression of the neuronal fate. 9. This model of neuronal induction by a BMP-4 signaling pathway is evolutionarily conserved in Drosophila. Studies of flies carrying mutations in decapentapelegic (dpp) [homolog of BMP-4] and short gastrulation (sog) [homolog of chordin], suggest that these molecules serve the same function in invertebrate neural induction. 10. The TGF-β signaling pathway, exemplified by BMP-4, is reiterated throughout the body of the developing embryo by other members of the TGF-β gene family to generate many other vertebrate tissues and organs. References: 1. Hemmati-Brivanlou, A. and Melton, D. (1997) Vertebrate embryonic cells will become nerve cells unless told otherwise. Cell 88: 13-17. 2. Harland, R and Gerhart, J. (1997) Formation and function of Spemann’s organizer. Annual Review of Cell and Developmental Biology 13: 611-67. 3. Gilbert, S. F. Developmental Biology, 3rd Edition. (1997) Chapter 15, Specification of cell fate by progressive cell-cell interactions. pages 591-633. 3 Neurogenesis: Inductive Mechanisms in a Nutshell. 1. Neuroectodermal cells choose either a neuronal or epidermal cell fate. 2. Interactions between mesoderm and neuroectoderm induce neuroectoderm to adopt the neural fate. 3. Induction acts through signaling by a secreted protein, Bone Morphogenic Protein-4 (BMP-4), made by neuroectodermal cells. 4. BMP-4 inhibits neuralization and promotes the epidermal fate in neighboring cells. 5. Mesodermal cells secrete proteins (Chordin, Noggin, Follistatin) which directly bind and antagonizes BMP-4 activity. 6. Neuroectodermal cells become neurons by suppression of BMP-4 activity by secreted proteins from underlying mesodermal cells. 7. The “default” state of neuroectodermal cells is neuronal. 8. This inductive mechanism is conserved between vertebrates and invertebrates. 9. BMP-4 is a member of the Transforming Growth Factor (TGF-β) family of signaling molecules. Similar signaling events maybe locally re-employed later in the developing nervous system. 4