Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
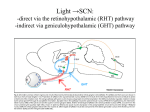
Circadian control of hormonal rhythms: important roles for suprachiasmatic nucleus efferents and the autonomic nervous system ANDRIES KALSBEEK, INGE F. PALM, SUSANNE E. LA FLEUR AND RUUD M. BUIJS. NETHERLANDS INSTITUTE FOR BRAIN RESEARCH MEIBERGDREEF 33, 1105 AZ AMSTERDAM THE NETHERLANDS. 1. Introduction Hormonal rhythms express themselves in a variety of forms. The two major processes controlling the shape of a hormonal rhythm are the circadian signal generated by the central pacemaker and the behaviour of an animal. The central neural pacemaker in humans and other mammals is localized in the suprachiasmatic nuclei of the hypothalamus (SCN). Little is known, however, about the neural mechanisms responsible for the implementation of rhythmic information generated by the SCN into behavioural and physiological rhythms. The endogenous pacemaker capacity of the SCN is evidenced by the rhythmic metabolic and electrical activity of its neurons 1,9, but also in the release of its transmitters. Best studied in this regard is the secretion of vasopressin (VP), showing a peak release during (subjective) daytime both in vivo and in vitro 9,16 . But rhythmic fluctuations in the SCN tissue content of peptide and mRNA have also been reported for somatostatin, VIP, GRP and GABA 12. Therefore, we put forward the following hypothesis:“The rhythmic release of SCN transmitters within its target areas is responsible for the expression of circadian rhythms in locomotor activity, body temperature, autonomic functions, and hormone secretion”. To test the above hypothesis additional information is needed about the brain structures towards which SCN output is directed, i.e. what are the target areas of the SCN efferents. Since the discovery of the SCN as “master clock” several attempts have been made to elucidate it neuronal projections (eg. 2,11,22,34). Taken together all animal studies indicate that the outflow of SCN information is mainly directed to medial hypothalamic target areas such as preoptic, paraventricular (PVN), dorsomedial (DMH) and ventromedial nuclei, with the exception of the paraventricular thalamus and the intergenicular leaflet. 25 2.1 Experimental strategy The main experimental strategy employed to test the above hypothesis included the intracerebral administration of SCN transmitters or their antagonists in SCN target areas of SCN-lesioned and intact animals, and the concomitant measurement of its consequences for the release pattern of the hormonal rhythm under study. We first concentrated on the SCN projection to the neuroendocrine center of the hypothalamus, i.e. PVN/DMH area, and the possible implication of this projection for the circadian control of the corticosterone rhythm. In order to test the general applicability of the hypothesis, additional experiments were extended to other hormonal rhythms and other SCN target areas. In the present chapter experiments will be described investigating the circadian control of corticosterone, melatonin, luteinizing hormone and insulin release. 2.2 Corticosterone Circadian fluctuations in circulating glucocorticoid levels have been reported for many species. In nocturnal animals such as the laboratory rat, plasma glucocorticoid levels are high at the onset of darkness and then decline, reaching a nadir in the morning. Our initial experiments using microinfusions in the PVN/DMH area showed a strong inhibitory effect of exogenous VP, but not VIP, on the release of corticosterone 17. These results suggested an inhibitory effect of VP released by SCN terminals in the PVN/DMH area on the activity of the hypothalamo-pituitary-adrenal (HPA)-axis. The initial data were further substantiated by microdialysis mediated administration of VP and its V1-antagonist. Stress-free infusion of the VP-antagonist in the DMH of freely moving, undisturbed animals during the middle of the light period (i.e. the trough of the corticosterone rhythm), caused an immediate dose-dependent increase of circulating plasma corticosterone. On the other hand, similar infusions of VP at the end of the light period completely prevented the diurnal rise in plasma corticosterone 23. In the above studies only single time points were investigated. Therefore, to further specify the nature of the SCN control, intracerebral infusions of VP-antagonist were performed at different times of the L/Dcycle. One hour infusions of the VP-antagonist were performed using in vivo microdialysis at CT2, 6, 10, 14 and 21, revealing the existence of a stimulatory SCN input to the HPA-axis as well, next to the inhibitory control of the VPergic SCN projection 24. The chemical identity of this stimulatory SCN transmitter is not know as yet, presently we only have preliminary data for a stimulatory action of VIP in the PVN/DMH area 14. Comparing congruent corticosterone and ACTH responses showed that whereas pronounced changes in circulating levels of corticosterone were induced, ACTH levels only showed minor changes. In addition, anatomical studies showed that 26 indeed the number of SCN fibers making synpatic contacts with CRH containing neurons in the PVN was very small 4,35. These results indicated that the SCN does not control the daily rhythm of corticosterone release solely via the HPA-axis (i.e. via the subsequent release of CRH and ACTH). Comparing the ACTH - corticosterone relation in different experimental conditions 7,24 revealed to us an alternative pathway for SCN control over corticosterone release. As indicated in Figure1 the circadian release of corticosterone does not depend heavily on the release of ACTH, contrary to the stress-related corticosterone release. Instead of stimulating ACTH release the SCN seems to change adrenal sensitivity for ACTH, probably via the sympathetic innervation of the adrenal gland. In order to investigate if indeed there is a neural connection between the SCN and the adrenal gland we employed the viral transneuronal tracing technique. Figure 1: Peak plasma corticosterone values during Ringer’s (ɀ, solid line) or VPantagonist (័, stippled line) administration at different times of the L/Dcycle, as a function of plasma ACTH. Contrary to novelty-stress (ᔤ, dashed line), circadian corticosterone excursions show minimal ACTH increments. This technique is based on the demonstrated ability of the virus to invade axon terminals, replicate in neurons and pass retrogradely through a multisynpatic circuit. After injection in the adrenal gland immunohistochemical localization of the viral antigen revealed the progressive appearance of infected neurons in the spinal cord, parvicellular subdivisions of the PVN and subsequently the SCN 6. The neurons in the spinal cord were shown to receive an input from 27 oxytocin fibers 30. Some of the virus-containing neurons in the PVN also exhibited oxytocin immunoreactivity. These latter neurons were shown to receive an input from VP- or VIP-containing SCN fibers 31. Furthermore, it was shown that denervation of adrenal gland by removal of its sympathetic innervation resulted in increased secretory activity of the adrenal cortex during the daytime 13, thus supporting the existence of an important daytime inhibition via the neural input to the adrenal gland. Together these results indicated that SCN inputs to PVN neurons with descending projections to preganglionic neurons of the sympathetic branch of the autonomic nervous system might be an important effector pathway for the SCN to impose its control on the corticosterone rhythm. Interestingly, SCN control on the circadian melatonin rhythm has already been known for a long time to involve the sympathetic innervation to the pineal gland. 2.3 Lutinizing Hormone In female rats the pre-ovulatory surge of luteinizing hormone (LH) occurs at regular 4 to 5 day intervals, and induces ovulation on the day of estrus. Previous studies had indicated two brain structures of crucial importance for the occurrence of LH surges. First, the medial preoptic area (MPO) containing the estrogen-receptor containing neurons necessary for the positive estrogen feedback. Secondly, the SCN providing the timing of the LH surge on the day of proestrus. Both MPO- and SCN-lesioned animals are completely incapable of showing LH surges 33. Anatomical studies have shown that estrogen-receptor containing neurons in the MPO receive direct synaptic inputs from SCN fibers, probably containing VP as a neurotransmitter 11,22,36. We hypothesized that VP release in the MPO would act as the daily timing signal of the SCN necessary for the induction of an LH surge. Indeed, a 5-h perfusion of the MPO with VP elicited a surge-like LH pattern in SCN-lesioned animals, whereas constant, basal levels were found in SCN-lesioned animals with control infusions 27. These results show that with regard to the LH surge VP may act as a stimulatory timing signal from the SCN, contrary to its inhibitory effect on the corticosterone rhythm. More importantly, however, these experiments indicate that VP can act as an output signal of the biological clock in different SCN target areas and completely different hormonal rhythms. 2.4 Melatonin Previous experiments with SCN-lesioned animals yielded equivocal results with respect to the abolishment of the circadian release of the pineal hormone melatonin. In our own experiments, using the transpineal microdialysis technique to measure melatonin release 15, 30% of the behaviourally arhythmic animals still showed nocturnal elevations of pineal melatonin levels. Effectively SCN-lesioned 28 animals, however, showed fairly constant extracellular melatonin levels, which were comparable to night time levels in control animals (Figure 2). These results indicated that SCN-lesions removed not only the circadian input to the pineal gland but also an important inhibitory input. The transneuronal viral tracing technique showed that also in the case of the melatonin rhythm SCN projections to autonomic PVN neurons were an important link 25,32. Figure 2: Melatonin release patterns during the first part of the dark period in SCNintact (ɂ) and SCN- lesioned (ɀ) animals. The shaded area shows the complete nocturnal melatonin profile of a previous group of control animals. Only animals were included which showed peak levels of melatonin significantly above those of control animals during the light period (> 400 pg/ml.). Our initial studies investigated the effect of both VP and VIP inputs to the dorsal hypothalamus on melatonin release. However, neither SCN peptide showed a clear inhibitory effect 20. One of the most abundant inhibitory transmitters in the hypothalamus is GABA, eg. in some hypothalamic nuclei about half of the total synpatic input may be accounted for by GABA-containing terminals. At that same time both anatomical and electrophysiological data indicated the presence and release of GABA from SCN terminals 3,5,10. Accordingly, experiments were initiated to determine whether the GABAergic projection from the SCN might be involved in depressing the activity of neurons in the dorsal hypothalamus that are associated with the control of the circadian me- 29 latonin rhythm. Infusion of the GABA-agonist muscimol in the dorsal hypothalamus completely blocked the daily increase of plasma melatonin during darkness and experiments in SCN-lesioned animals showed that also the elevated plasma melatonin levels in these animals were decreased by the muscimol administration 19. On the other hand, infusion of the GABA-antagonist bicuculline had no effect on the nocturnal melatonin increase. Muscimol infusions elicited a decrease of extracellular noradrenaline levels in the pineal concomittant with decreased melatonin release showing clearly that the inhibitory influence of the SCN is mediated via the sympathetic input to the pineal gland 19. In conjunction with the strong resemblance between the acute inhibitory effect of muscimol and light on melatonin release, these data suggested to us that the GABA-containing projections from the SCN might serve to tr?ÿsmit the inhibitory effect of light onto the melatonin-rhythm-generatingsystem in the dorsal hypothalamus. Therefore, we set out to test the hypothesis that the light-induced inhibition of melatonin is mediated by the endogenous release of GABA within the dorsal hypothalamus. Animals were confronted with a 1-min nocturnal light exposure in the middle of a bilateral 2-h perfusion of the dorsal hypothalamus with either Ringer or Ringer + Bicuculline. It is clear from Figure 3 that bicuculline completely prevented the inhibitory effect of light on melatonin release. This experiment thus documents that retina-mediated photic activation of SCN neurons induces the release of GABA from efferent SCN nerve terminals, resulting in an inhibition of melatonin release by the pineal gland 18. At present it is not clear yet if GABA release from SCN terminals is also involved in the circadian control of the daily melatonin rhythm. In this aspect the reported co-localization of GABA and vasopressin in SCN neurons and their efferent projections is intriguing5, since the circadian rhythm in synthesis and release of vasopressin, with peak activity found during (subjective) day time, is well-known16. 2.5 Insulin The daily rhythm of insulin release seems to be mainly driven by a rise in blood glucose levels as a consequence of a circadian pattern in feeding behaviour, and a direct circadian modulation of insulin release is not evident. After SCN lesions glucose and insulin responses are similar during day and night, but also the distribution of the meals across the light/dark cycle is equal now 28. Therefore, it might still be so that the distribution of feeding activity is the primary determinant of daily variations in insulin and glucose responses, and not the circadian oscillator located in the SCN. Lesion experiments did indicate, however, that the SCN exerts an important inhibitory control on insulin release 28, as previously found for corticosterone and melatonin. To further investigate SCN control of insulin release we designed an experimental setup 30 that would enable us to unmask a possible direct control of the endogenous pacemaker on the circadian aspects of insulin release. Rats were subjected to a Figure 3: Inhibitory effect of a 1-min light exposure at ZT17 on melatonin levels (n=6). During light exposure (stippled line) hypothalamic microdialysis probes aimed at the PVN/DMH area were perfused with either Ringer alone (ɂ) or Ringer + Bicuculline (ɀ). The gray box indicates the 2-h hypothalamic perfusion period. feeding regimen of 6 meals a day, spaced 4 hours apart. A significant daily variation in feeding responses was present despite the fact that feeding conditions, such as prior fasting period, amount of food intake and intake rate, were very much similar now for all six meals 21. It is evident that the most prominent (inhibitory) effects of the circadian timing system on feeding-induced insulin release are present at the end of the light period (Figure 4). With regard to the basal insulin levels, however, a daily rhythm was not evident. On the other hand basal glucose levels did show a significant circadian variation with highest glucose levels being reached at the end of the light period 21, i.e. shortly before the onset of the dark period and the main period of (feeding) activity of the rat. Together these data indicate that the SCN controls glucose homeostasis at two levels. First basal glucose levels are controlled to ensure sufficient glucose supply to active tissues at times when the animal awakens but is not feeding yet. Secondly, feeding-induced, but not basal, insulin release is stimulated (or disinhibited) only at the animals usual feeding time to prevent hypoglycemia due to increased insulin release before the animal has been able to find food. At present we have no indications yet which SCN-projections and/or -transmit- 31 ters might be involved in the control of basal glucose and feeding-induced insulin responses. As described in the foregoing we think SCN control over sympathetic inputs to the adrenal is an important mechanism to control circadian corticosterone release. A similar mechanism may account for the control on glucose homeostasis. Increased sympathetic input to the liver and fat tissue enhances glucose output and stimulates lipolysis, thus promoting hyperglycemia. Electrical stimulation of the SCN too results in hyperglycemia 8,26. Therefore, increased SCN activity at the end of the light period may stimulate sympathetic activity in different tissues resulting in both a corticosterone and a glucose peak. At the same time, as becomes clear from Figure4, the stimulatory effect of the SCN on corticosterone secretion and plasma glucose levels coincides exactly with its inhibitory effect on the feeding-induced insulin responses. Our previous experiments have shown that the pronounced daily rhythmicity Figure 4: Maximal feeding-induced insulin increments (bars) during 6 meals equally dispersed over the 24- hour L/D-cycle and maximal corticosterone increases (open symbols) due to a 1-hour blockade of the VPergic SCN output to the DMH (by hypothalamic administration of VP-antagonist) at five different times along the L/D-cycle. in insulin responses is most likely effectuated via an inhibitory action of the SCN on the parasympathetic input to the pancreas 21,28. Thus, congruent with a stimulatory effect of SCN output on sympathetic activity, parasympathetic activity is inhibited. Indeed, a considerable proportion of PVN neurons with 32 descending projections send divergent axon collaterals to both the spinal cord and the dorsal vagal complex in the brainstem 29. Therefore, an important aspect of SCN control on daily rhythms may involve its inputs to PVN neurons with descending projections and its effect on the sympathovagal balance of the autonomic nervous system 30,31. 3. Summary The results obtained so far show that the rhythmic release of VP and GABA from SCN terminals are important signals for the control of hormonal rhythms such as corticosterone, melatonin and LH. The ultimate circadian waveform of a hormonal rhythm seems to be determined by an alternation of inhibitory and stimulatory signals from the SCN. For a number of rhythms evidence has been obtained that an important part of the circadian message from the SCN is transferred by modulating the sympathovagal balance of the autonomic input to hormonal glands or other peripheral organs. The presently proposed mechanism for the implementation of circadian rhythms may hold not only for hormonal rhythms, but also for SCN control on behavioral (i.e. sleep-wake) and physiological rhythms, such as body temperature and cardiovascular regulation. Acknowledgements We gratefully acknowledge support from IRIS (grant No. PHA-614-NLD) and the superb technical assistence from J. Van der Vliet, J.J. van Heerikhuize and J. Wortel. References 1 Bos N.P.A. and Mirmiran M. (1990) Circadian rhythms in spontaneous neuronal discharges of the cultured suprachiasmatic nucleus. Brain Res. 511, 158-162. 2 Buijs, R.M. The anatomical basis for the expression of circadian rhythms: the efferent projections of the suprachiasmatic nucleus. In: Progress in Brain Research, Vol.111, Hypothalamic Integration of Circadian Rhythms, edited by Buijs, R.M., Kalsbeek, A., Romijn, H.J., Pennartz, C.M.A. and Mirmiran, M. Amsterdam: Elsevier Science BV., 1996, p. 229-240. 3 Buijs R.M., Hou Y.X., Shinn S. and Renaud L.P. (1994) Ultrastructural evidence for intra- and extranuclear projections of GABAergic neurons of the suprachiasmatic nucleus. J. Comp. Neurol. 340, 381-391. 4 Buijs R.M., Markman M., Nunes-Cardoso B., Hou Y.-X. and Shinn S. (1993) Projections of the suprachiasmatic nucleus to stress-related areas in the rat hypothalamus: A light and electron microscopic study. J. Comp. Neurol. 335, 42-54. 5 Buijs R.M., Wortel J. and Hou Y.X. (1995) Colocalization of gamma-aminobutyric acid with vasopressin, vasoactive intestinal peptide, and somatostatin in the rat 33 suprachiasmatic nucleus. J. Comp. Neurol. 358, 343-352. 6 Buijs R.M., Wortel J., Van Heerikhuize J.J., Feenstra M.G.P., Ter Horst G.J., Romijn H.J. and Kalsbeek A. (1999) Anatomical and functional demonstration of a multisynaptic suprachiasmatic nucleus adrenal (cortex) pathway. Eur. J. Neurosci. 7 Buijs R.M., Wortel J., VanHeerikhuize J.J. and Kalsbeek A. (1997) Novel environment induced inhibition of corticosterone secretion: Physiological evidence for a suprachiasmatic nucleus mediated neuronal hypothalamo-adrenal cortex pathway. Brain Res. 758, 229-236. 8 Fujii T., Inoue S., Nagai K. and Nakagawa H. (1989) Involvement of adrenergic mechanism in hyperglycemia due to SCN stimulation. Horm. Metab. Res. 21, 643-645. 9 Gillette M.U. and Reppert S.M. (1987) The hypothalamic suprachiasmatic nuclei: circadian patterns of vasopressin secretion and neuronal activity in vitro. Brain Res. Bull. 19, 135-139. 10 Hermes M.L.H.J., Coderre E.M., Buijs R.M. and Renaud L.P. (1996) GABA and glutamate mediate rapid neurotransmission from suprachiasmatic nucleus to hypothalamic paraventricular nucleus in rat. J. Physiol-London. 496, 749-757. 11 Hoorneman E.M.D. and Buijs R.M. (1982) Vasopressin fiber pathways in the rat brain following suprachiasmatic nucleus lesioning. Brain Res. 243, 235-241. 12 Inouye, S.I.T. Circadian rhythms of neuropeptides in the suprachiasmatic nucleus. In: Progress in Brain Research, Vol.111, Hypothalamic Integration of Circadian Rhythms, edited by Buijs, R.M., Kalsbeek, A., Romijn, H.J., Pennartz, C.M.A. and Mirmiran, M. Amsterdam: Elsevier Science BV., 1996, p. 75-90. 13 Jasper M.S. and Engeland W.C. (1994) Splanchnic neural activity modulates ultradian and circadian rhythms in adrenocortical secretion in awake rats. Neuroendocrinology 59, 97-109. 14 Kalsbeek, A. and Buijs, R.M. Peptidergic transmitters of the suprachiasmatic nuclei and the control of circadian rhythmicity. In: Progress in Brain Research, Vol.92, The Peptidergic Neuron, edited by Joosse, J., Buijs, R.M. and Tilders, F.J.H. Amsterdam: Elsevier Science BV, 1992, p. 321-333. 15 Kalsbeek, A. and Buijs, R.M. Rhythms of inhibitory and excitatory output from the circadian timing system as revealed by in vivo microdialysis. In: Progress in Brain Research, Vol.111, Hypothalamic Integration of Circadian Rhythms, edited by Buijs, R.M., Kalsbeek, A., Romijn, H.J., Pennartz, C.M.A. and Mirmiran, M. Amsterdam: Elsevier Science BV, 1996, p. 271-291. 16 Kalsbeek A., Buijs R.M., Engelmann M., Wotjak C.T. and Landgraf R. (1995) In vivo measurement of a diurnal variation in vasopressin release in the rat suprachiasmatic nucleus. Brain Res. 682, 75-82. 17Kalsbeek A., Buijs R.M., Van Heerikhuize J.J., Arts M. and Van Der Woude T.P. (1992) Vasopressin-containing neurons of the suprachiasmatic nuclei inhibit corticosterone release. Brain Res. 580, 62-67. 18 Kalsbeek A., Cutrera R.A., Van Heerikhuize J.J., Van Der Vliet J. and Buijs R.M. 34 (1998) GABA release from SCN terminals is necessary for the light-induced inhibition of nocturnal melatonin release in the rat. Neuroscience 19 Kalsbeek A., Drijfhout W.J., Westerink B.H.C., Van Heerikhuize J.J., Van Der Woude T., Van Der Vliet J. and Buijs R.M. (1996) GABA receptors in the region of the dorsomedial hypothalamus of rats are implicated in the control of melatonin. Neuroendocrinology 63, 69-78. 20 Kalsbeek A., Rikkers M., Vivien-Roels B. and Pévet P. (1993) Vasopressin and Vasoactive Intestinal Peptide Infused in the Paraventricular Nucleus of the Hypothalamus Elevate Plasma Melatonin Levels. J. Pineal Res. 15, 46-52. 21 Kalsbeek A. and Strubbe J.H. (1998) Circadian control of insulin secretion is independent of the temporal distribution of feeding. Physiol. Behav. 63, 553-560. 22 Kalsbeek A., Teclemariam-Mesbah R. and Pévet P. (1993) Efferent projections of the suprachiasmatic nucleus in the golden hamster (Mesocricetus auratus). J. Comp. Neurol. 332, 293-314. 23 Kalsbeek A., Van Der Vliet J. and Buijs R.M. (1996) Decrease of endogenous vasopressin release necessary for expression of the circadian rise in plasma corticosterone: a reverse microdialysis study. J. Neuroendocrinol. 8, 299-307. 24 Kalsbeek A., Van Heerikhuize J.J., Wortel J. and Buijs R.M. (1996) A diurnal rhythm of stimulatory input to the hypothalamo-pituitary-adrenal system as revealed by timed intrahypothalamic administration of the vasopressin V1 antagonist. J. Neurosci. 16, 5555-5565. 25 Larsen P.J., Enquist L.W. and Card J.P. (1998) Characterization of the multisynaptic neuronal control of the rat pineal gland using viral transneuronal tracing. Eur. J. Neurosci. 10, 128-145. 26 Nagai K., Fujii T., Inoue S., Takamura Y. and Nakagawa H. (1988) Electrical stimulation of the suprachiasmatic nucleus of the hypothalamus causes hyperglycemia. Horm. Metab. Res. 20, 37-39. 27 Palm I.F., Van Der Beek E.M., Wiegant V.M., Buijs R.M. and Kalsbeek A. (1999) Vasopressin induces an LH surge in ovariectomized, estradiol-treated rats with lesion of the suprachiasmatic nucleus. Neuroscience 28 Strubbe J.H., Alingh Prins A.J., Bruggink J. and Steffens A.B. (1987) Daily variation of food-induced changes in blood glucose and insulin in the rat and the control by the suprachiasmatic nucleus and the vagus nerve. J. Auton. Nerv. Syst. 20, 113-119. 29 Swanson L.W. and Kuypers H.G.J.M. (1980) The paraventricular nucleus of the hypothalamus: Cytoarchitectonic subdivisions and organization of projections to the pituitary, dorsal vagal complex, and spinal cord as demonstrated by retrograde fluorescence double-labeling methods. J. Comp. Neurol. 194, 555-570. 30 Teclemariam-Mesbah R., Kalsbeek A., Buijs R.M. and Pévet P. (1997) Oxytocin innervation of spinal preganglionic neurons projecting to the superior cervical ganglion in the rat. Cell Tiss. Res. 287, 481-486. 31 Teclemariam-Mesbah R., Kalsbeek A., Pévet P. and Buijs R.M. (1997) Direct vasoac- 35 tive intestinal polypeptide-containing projection from the suprachiasmatic nucleus to spinal projecting hypothalamic paraventricular neurons. Brain Res. 748, 71-76. 32 Teclemariam-Mesbah R., Ter Horst G.J., Postema F., Wortel J. and Buijs R.M. (1999) Anatomical demonstration of the suprachiasmatic nucleus - pineal pathway. J. Comp. Neurol. 33 Van Der Beek, E.M. Circadian control of reproduction in the female rat. In: Progress in Brain Research, Vol.111, Hypothalamic Integration of Circadian Rhythms, edited by Buijs, R.M., Kalsbeek, A., Romijn, H.J., Pennartz, C.M.A. and Mirmiran, M. Amsterdam: Elesevier Science BV, 1996, p. 295-320. 34 Van Der Beek E.M., Horvath T.L., Wiegant V.M., Van Den Hurk R. and Buijs R.M. (1997) Evidence for a direct neuronal pathway from the suprachiasmatic nucleus to the gonadotropin-releasing hormone system: Combined tracing and light and electron microscopic immunocytochemical studies. J. Comp. Neurol. 384, 569-579. 35 Vrang N., Larsen P.J. and Mikkelsen J.D. (1995) Direct projection from the suprachiasmatic nucleus to hypophysiotrophic corticotropin-releasing factor immunoreactive cells in the paraventricular nucleus of the hypothalamus demonstrated by means of Phaseolus vulgaris-leucoagglutinin tract tracing. Brain Res. 684, 61-69. 36 Watson R.E., Langub M.C., Engle M.G. and Maley B.E. (1995) Estrogen-receptive neurons in the anteroventral periventricular nucleus are synaptic targets of the suprachiasmatic nucleus and peri-suprachiasmatic region. Brain Res. 689, 254-264. 36 Body temperature and sleep wake rhythms torn apart: implications for the regulation of mood DOMIEN G.M. BEERSMA DEPARTMENT OF BIOLOGICAL PSYCHIATRY AND ZOOLOGICAL LABORATORY, UNIVERSITY OF GRONINGEN, GRONINGEN, THE NETHERLANDS The prior history of sleep and wakefulness and the current phase position of the circadian pacemaker (biological clock) both have a major impact on all kinds of physiological and psychological processes, including the course of body temperature and many aspects of performance and well being. Under normal circumstances all sleep episodes occur within a restricted range of circadian phases. Therefore, it is difficult to discriminate between those two major influences. Experimentally, the range of circadian phases at which sleep occurs can be increased, for instance by scheduling sleep at 20 hour intervals. At low light levels it turns out that the circadian pacemaker will no longer run in synchrony with the 20 hour sleep-wake cycle. Instead it will free run with a period slightly longer than 24 hours. Under such conditions of forced desynchrony sleep systematically scans through all circadian phases. Forced desynchrony experiments revealed that mood in healthy subjects varies systematically with circadian phase and with the time since wake-up. The circadian component shows maximum feelings of depression around the minimum in the endogenous rhythm in core body temperature. The component related to the timing of sleep shows a progressive increase in feelings of depression in the course of prolongued wakefulness. Inspired by these findings we embarked to investigate the circadian regulation of mood in seasonal affective disorder (SAD) patients, since it has been suggested that the mood disturbances in SAD are due to circadian abnormalities. The results of the very first forced desynchrony studies in an SAD patient will be presented. 37