Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
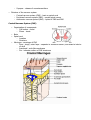
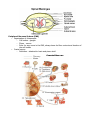
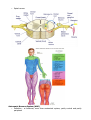
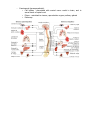
THE NERVOUS SYSTEM The individual neuron cannot do much alone, it must be interconnected with others in order to produce meaningful pattern of neural activity to regulate animal behaviours; this aggregate is the nervous system. This is a common term for a hugely rich and varying body of systems. In the main, there are two distinct groups of nervous system, the vertebrates and the invertebrates. Invertebrate nervous systems tend to be smaller and simpler than vertebrate ones. They vary widely in design and extent, from the sponge with no nervous system to the octopus which has almost vertebrate complexity. Many invertebrate nervous systems are distributed about the body of the animal with only a small, rudimentary brain in the head, the more complex the animal the larger the brain. In the vertebrate most of the complex neural behaviour is located in one place, the head. The number of neurons in invertebrates is usually limited and in some instances appears to be constant among individual animals of the same species. Yet between species the number varies widely. Neuronal organisation between these two type of animal is distinct also. Most neurons in the brains of vertebrates are usually bipolar or multipolar, with one or several dendrites extending from one side of the cell body and an axon from the other. Most invertebrate are monopolar, i.e. a single protrusion is made from the cell body and both axons and dendrites extend from this. An invertebrate neuron can have several axons within the dendritic tree with its own axon hillock. Invertebrates often develop more complex neural circuitry by making individual neurons more complex, unlike vertebrates who add on more neurons. There is evidence that the latter method seems to be less limiting than the prior. BEHAVIOUR Animal behaviour can be divided up into several broad classes, all of which are governed by different combinations of processes in the brain/nervous system. The simplest behaviour is the reflex, a stereotyped response triggered by stimuli in the environment, e.g. removing a part of the body from a pain inducing stimulus. The intensity and duration of the response is dependant on the force and duration of the stimulus. Reflexes are quick, instinct reactions which allow an animal to adjust its behaviour in response to sudden environmental changes. Taxes, also known as orientation behaviour, e.g. light following, is where the animal will turn itself towards, or away from an environmental agent. Fixed-action patterns are the most complex group of reactive behaviours, the stimulus involved is usually more complex than that involved in reflex actions, and it leads to an extended, usually stereotyped response to a sensory stimulus. There are also behaviours called motivated behaviours, which are governed primarily by the internal state of the animal. For example, feeding is not just based on the availability of food, but also on the animal's need for it, i.e. hunger. characterised motivational behaviour with six points: 1. grouping and sequencing of component behaviour in time 2. goal directedness: the sequence of component behaviours generated can only be understood by reference to some goal 3. spontaneity: the behaviour can occur in the absence of the eliciting stimuli 4. changes in responsiveness: the modularity effect of the motivational state varies depending upon its level of arousal or satiation 5. persistence: the behaviour can greatly outlast any initiating stimulus 6. associative learning. These behaviours are grouped together and combined in a hierarchy which elicit the total behaviour of an animal. Taxes, reflexes and fixed-action patterns are all found in most animals, governed by some type of motivational control. Some behaviours overrule others in the scheme of things, others taking over from each other depend on internal and external state. The hierarchy is dependant in a lot of cases on environmental context, it is not fixed and rigid. These combinations will be discussed in more detail in the next two sections with specific examples. INVERTEBRATE NERVOUS SYSTEMS Behaviour Mechanisms Some behaviours in invertebrates can be attributed to specialised physiological properties of certain neurons. For example, the sea snail Aplysia produces ink when a prolonged and strong tactile stimulus is applied, i.e. it tries to blind any predator. However, the ink is noxious to it as well, so unless the stimulus is prolonged, it does not release the ink. This delayed response can be linked to special voltage sensitive channels in the motor neurons involved in the behaviour, which oppose the normal depolarisation of the cell by the sensory inputs. The depolarisation for activation is therefore much more slow in activating. However other mechanisms underlie other behaviours. Neuronal circuits, more than one neuron working in tandem with each other, are responsible for, for example the lateral eye in Limulus, the horseshoe crab. The eye in Limulus is a compound eye made up of separate photoreceptive units. Each unit contains about fifteen photoreceptive cells. These cells are all coupled to one process, the eccentric cell where the action potentials are generated. Each eccentric cell is networked laterally to adjacent eccentric cell axons to form inhibitory synapses. An axon inhibited by its neighbour in turn inhibits that one. This creates a system where axons of eccentric cells along a bright side of a dark-light border are inhibited weakly by their neighbours in the dimmer light. Thus they generate more actions potentials than brightly lit cells away from the border which are inhibited strongly by all their neighbours. Conversely, the axons on the dimmer side of the border are strongly inhibited and fire less vigorously than other dark axons away from the border. Therefore, the perception of the border between light and dark is enhanced. The study of the lateral in the Limulus provides a model of perception based on neuronal circuits, collections of neurons working in tandem. However, there is also the specialized neuron model for the inking in Aplysia. These and other studies have lead to a conclusion that most information processing and behaviours are controlled at both levels, both cell and circuit. This function has been observed such that some neurons innervate a single patch of skin or muscle, but others can initiate complex behaviours involving many other neurons. These initiators are called command neurons. Rhythmic Motor Behaviour - Example Of Cell and Circuit Control The ability to move and carry out purposeful acts distinguishes animals from plants. Motor activity such as walking, swimming, breathing is rhythmic, i.e. it needs a stereotypical, repetitive sequence of neural outputs and muscle contractions. The neuronal circuit which controls rhythmic behaviour is termed a central pattern generator CPG. The Function of A CPG The Heart of The Leech The Heart Of The Leech A good example of rhythmic motor action is the heart beat, and one studied closely is the heart system of the leech. In the leech, blood is pumped around its body by two heart tubes running down each side of its body. The left and right tubes contract alternately. The heart tubes are innervated by motor neurons termed heart-excitatory, HE, neurons. The arrangement of these cells is an example of circuit level organisation. The leech has 21 ganglia, these motor neurons are found in ganglia 3 to 18 on both sides of each ganglion. For this reason they are called HE(R) and HE(L) cells, those which innervate the right and left tubes respectively. In addition, there are seven pairs of heart interneurons located in the first seven ganglia HN(1- 7) - these cells control and integrate the HE outputs and make up the CPG. The first four pairs HN cells HN(1-4) regulate the heart rhythm, they can reset and entrain the whole system. HN(5-7) cannot reset the system, but co-ordinate motor neuron inter-ganglia activity. All the HE cells are innervated by HN(3) and HE(5-18) have an extra innervation from HN(4). HN(1) and HN(2) do not directly innervate any motor neurons, but co-ordinate the activity if HN(3) and HN(4), which are the principle controllers of the heart rhythm. All synapse between interneurons and motor neurons are inhibitory, therefore activity in the interneurons is out of phase with the motor neurons. Without inhibition the motor neurons would fire continuously. Pairs of interneurons, i.e. the right and left of the same number inhibit each other reciprocally, so that when one is firing, the other is inhibited, and so the heart tubes contract alternately. However, this is not the whole picture, something is needed to terminate the inhibition after a certain time period, otherwise one neuron would be permanently inhibited and the other permanently on. This is where the cell level control comes in. Neurons possess a mechanism to oppose the hyperpolarisation which allows the cell to gradually depolarise. Therefore the inhibited interneuron will fire again even if the inhibition is constantly applied. Returning to circuit level control, HN(1) and HN(2) as previously mentioned, do not interact directly with the motor neurons, instead they are used to reciprocally inhibit HN(3) and HN(4). The Circuitry Of The First Four Heart Interneurons When HN(R3) and HN(R4) are active, HN(L3) and HN(L4) are inhibited, as are HN(R1) and HN(R2). When HN(L3) and HN(L4) are active, HN(R3), HN(R4), HN(L1) and HN(L2) are all inhibited. HN(4) is the orchestrator of the transitional activity, i.e. its time period to escape from inhibition controls the others. Figure 10 shows the cycle of inhibitions and activations in the leech. States A and C are quite long ~6 seconds, where as states B and D are short. The Inhibitory Cycle Of HN(1-4) The heart control of the leech is a prime example of the tandem working of circuit and cell level control. The inhibitory nature and cross connections of the interneuron synapses would be useless without the added speciality of the cell membranes to slowly depolarise and oppose the inhibition to allow the other states to exist. It is also important to recognise that it is not the nature of the cells in an isolated situation that produces the behaviour. There is a temporal aspect to the dynamics of the system, i.e. in the cylce shown above, teh working of the system is ocntrolled very much by the dischare rate of HN(4) and it is this cell changing in time that controls the oevrall behaviuor - a different discharge rate and the length and exact performance of the cycle could be changed. Neural Circuit Modulation Neuromodulation can redefine the circuitry of a CPG in a variety of ways: it can switch the circuit from one configuration to another so that quite different behaviours are mediated by the circuit; it can also fine tune a circuit gradually narrowing its behaviour to suit the situation. There are many different types of neuromodulator and a neural circuit can be affected by a large number of these chemicals and each has a specific affect. One neuromodulator can have multiple affects on a single neuron, or different effects on other cells. Neuromodulators can affect at a circuit and cell level. Both chemical and electrical synapses can be modulated, the neuromodulator can change membrane properties and synaptic strengths. The different types of neuromodulator give the neural circuitry adaptability are an important consideration in the modelling of neural networks. Memory and Learning Changes take place at synapses when those synapses are repeatedly activated, these changes can last for weeks, months, longer. These adaptations have been examined in detail, they will be discussed here using Aplysia as an example. The marine snail will withdraw its gills when a stimulus is applied. A gentle stimulus applied to the mantle/siphon causes rapid gill withdrawal. With repeated stimulation, the response diminishes, this is called habituation. If stimuli is repeated over several training sessions, habituation can last for weeks. A strong stimulus to the head or tail before a gentle mantle/siphon stimuli sensitises the reflex, i.e. the response is greater. Again, more training sessions mean longer lasting 'memory'. Neuronal Interaction Involved in Memory and Learning Habituation is localized to synapses between motor and sensory neurons, with repeated stimulus the sensory neurons release less and less neurotransmitter. Sensitisation is mediated via interneuronal synapses on sensory neuron terminals and action potentials from sensory neurons in the head and tail. Either head or tail related stimuli activate interneurons which release a neuromodulator at synapses with mantle sensory neurons. These cells then release more neurotransmitter onto the motor neurons than is normally the case, so the response is more pronounced. Long term habituation and sensitisation can create structural changes in the sensory neurons themselves. This kind of study add weight to the supposition that short term memory/learning is a chemical change in the neuronal circuit which will eventually die off, but with repeated stimulus, this can become a long term memory/learning as a structural change results from the neuromodulation. VERTEBRATE NERVOUS SYSTEMS The nervous system in the vertebrate consists of two parts, the central and peripheral nervous systems. The peripheral nervous system is made up of all the nerves and ganglia which lie outside the brain and spinal cord. The Central Nervous System A Diagrammatic View Of The Central Nervous System The Central nervous system consists of the spinal cord and the brain, and unlike invertebrate systems is highly centralised. All higher neural functions, perception, movement control, learning and memory are carried out in the brain. The spinal cord contains the CPG's for rhythmic motor behaviour, mediates reflexes and conducts sensory information from peripherals to the brain. Sensory neuron axons can do one of three things on entering the spinal cord. They can travel up the cord to its higher levels or the brain, they can connect with interneurons in the cord itself, or they can connect directly to motor neurons. The Brain The brain consists of three main structures, the hindbrain, midbrain and forebrain. hindbrain comes from the spinal cord and contains the medulla, cerebellum and pons. midbrain is a small region above the pons. The forebrain consists of two main units, containing the thalamus and hypothalamus and the other holding the basal ganglia cerebral cortex. The The one and Medulla and Pons Nerve tracts run through the medulla connecting the spinal cord and the higher brain centres. There are also many clusters of neurons which are involved in the control of the head, face, eyes, tongue and vital body functions. Also to be found distributed throughout the medulla and pons are groups of clusters of neurons called the reticular formation. Their axons extend widely throughout the rest of the brain and spinal cord. The formation regulates the level of activity in parts of the brain and is important in arousal and awareness levels. Destruction in this part of the brain can lead to coma from which there is no arousal. These formations also modulate pain pathways. Hypothalamus This is a regulatory centre of the brain - it is concerned with regulation of basic drives and acts, e.g. eating, sexual activity, heart rate. It can also effect emotional behaviour, so lesions can lead to aggressive behaviour. The hypothalamus also regulates hormone release - the hypothalamus controls hormone release from the pituitary gland which in turn controls the release of hormones from other glands throughout the body, such as thyroid. Some of the hormones released by the hypothalamus are released directly into circulation to their targets. Cerebellum The cerebellum has a highly regular structure and its main function is to co-ordinate and integrate motor behaviour. The cortex gives the commands for movement, but the cerebellum co-ordinates them with sensory input from the spinal cord and other sensory systems to produce a smooth output. Thalamus The thalamus consists of clusters of neurons which: convey sensory information to the cerebral cortex; or relay information about motor activity to the cortex. It is believed that the inputs to the thalamus modify the sensory and motor information before passing it on. Basal Ganglia These are five prominent clusters that are positioned above, below and either side of the thalamus. They are concerned with movement, its initiation and execution. They receive most of their information from the cortex and their output returns there via the thalamus. Cerebral Cortex This is where the higher central nervous system functions are collected. Skilled movements originate from here and it is considered the centre of consciousness, memory and intelligence. It is divided into two hemispheres, the left and the right, and each hemisphere has four lobes, frontal, parietal, occipital and temporal. The frontal lobes are concerned with movement and olfaction, the parietal lobes with somatic sensation, the occipital lobes with vision and the temporal lobes with audition and memory. Within each lobe are areas dedicated to the initial cortical analysis of specific sensory information or movements. Structures and Pathways in the vertebrate brain The Development of The Nervous System The nervous system is a remarkable phenomenon in nature, highly complex, consisting of almost incalculable numbers of nerves, neurons and their connections. How it develops is of prime concern when designing a model. The nervous systems cells all derive from neural plate cells Daughter cells produced after cell division one produced migrate to their final positions, passing through an intermediate zone on the way. Some neurons divide after migration and later form certain brain structures. There are two theories to explain cell differentiation in the nervous system: 1 - the lineage theory states that cells inherit developmental directives; 2 - the other suggests that cells differentiate as a consequence of environmental clues, chemical signals. Axon formation is another aspect of the nervous system development. Chemical affinity suggests that cells have specific chemical markers which allow them to recognise each other during development and regeneration. There is a certain amount of flexibility in the formation of connections during these stages. There is also evidence for competition for space during normal brain development, if axons do not compete successfully, their terminal arbors are restricted and they end up with fewer synaptic connections. Axonal formation is all very well, but the guidance of these structures to the correct places in the brain to make connections is also necessary. Chemical gradients are a likely mechanism, i.e. the axon growth follows the gradient, another is that certain cells act as guideposts along the way. the axon grows towards them due to chemical attractors, makes a connection and then continues, removing these guideposts can lead to stunting in axonal growth. Axons may also follow pioneer cells, cells which form connections early in development when distances between brain structures are short. One it has been developed, the nervous system has to mature in which time the synaptic connections and neuronal fields are subject to rearrangement and restriction. Also cells die during this development due to competition for synaptic sites and lack of innervation. An agent involved in this process, known as nerve growth factor, promotes: the survival of certain neurons; the growth of processes from specific neurons; guides axonal growth. Functional unit • Neuron - a cell Definitions o Cell body - soma or perikaryon o Fiber - neurites, may be afferent or efferent o • Synapse - release of neurotransmitters Divisions of the nervous system o Central nervous system (CNS) - brain or spinal cord o Peripheral nervous system (PNS) - cranial spinal nerves o Autonomic nervous system (ANS) - hybrid of CNS and PNS Central Nervous System (CNS) • • • • Organization of components o Cell bodies - nuclei o Fibers - tracts Brain Spinal cord o Location o Structure Meninges - coverings of CNS o Dura - tough, outer layer - separate in vertebral column, but fused to interior of skull o Arachnoid - web-like membrane o Pia - intimate surface of CNS structures Peripheral Nervous System (PNS) • Organization of components o Cell bodies - ganglia o Fibers - nerves o Rule: for any nerve in the PNS, always know its fiber content and location of the cell bodies • Cranial nerves o Definition - attached to brain and pierce skull • Spinal nerves Autonomic Nervous System (ANS) • Definition - a functional, more than anatomical system, partly central and partly peripheral • Rules o o o o • Efferent only 2 neurons from CNS to target 1st neuron is in CNS - preganglionic or presynaptic 2nd neuron is in PNS - postganglionic or postsynaptic Divisions o Thoracolumbar (sympathetic) Cell bodies - intermedio-lateral gray of spinal cord segments, T1 - L2 Fibers - distributed throughout the body Pathways o Craniosacral (parasympathetic) Cell bodies - associated with cranial nerve nuclei in brain, and in sacral levels of spinal cord Fibers - restricted to viscera, reproductive organs, salivary glands Pathways