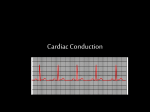
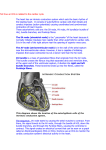
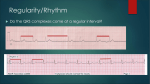
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Am J Physiol Heart Circ Physiol 286: H1853–H1871, 2004; 10.1152/ajpheart.01205.2003. The AV junction region of the heart: a comprehensive study correlating gross anatomy and direct three-dimensional analysis. Part II. Morphology and cytoarchitecture Darlene K. Racker Department of Medicine, Cardiology and The Feinberg Cardiovascular Research Institute, Northwestern University Feinberg Medical School, Chicago, Illinois 60611 Submitted 18 December 2003; accepted in final form 18 December 2003 atrial arrhythmia; atrioventricular node; atrioventricular node reentrant tachycardia; triangle of Koch; conduction; catheter ablation; electrophysiology; His bundle; dual atrioventricular node physiology; potentials THIS IS PART II OF A TWO-PART report on the anatomy of the atrioventricular (AV) junction region of the heart. Part I (55) detailed aspects of the gross anatomy and topography. In Part II, details of the histology are presented. An understanding of this anatomy is becoming increasingly more important as surgical and catheter ablation procedures are performed routinely in this region of the heart. Also, detailed knowledge of the anatomy is crucial for understanding electrical circuits and pathophysiology and for extending molecular biological inquiries. Address for reprint requests and other correspondence: D. K. Racker, Northwestern Univ. Feinberg Medical School, 303 E. Chicago Ave., CH-T 233, Chicago, IL 60611 (E-mail: [email protected]). http://www.ajpheart.org Because information on the AV junction region was incomplete, a correlated gross anatomic and histological study was undertaken utilizing protocols that preserve the interstitial and cytoplasmic integrity of paraffin-embedded tissues (see Fig. 1–8 to 1–11 in Ref. 48). For the AV junction region studies, the heart was naturally flattened to permit orthogonal plane sections, and a systematic comparison among photographs of whole hearts, subsequent tissue blocks, and photomicrographs of serial sections of the same hearts was made. The first series of studies demonstrated that a specialized myocardial network joins the AV node (i.e., three atrionodal bundles and the proximal AV bundle in two different longitudinal planes with respect to the AV junction region) and that the atrial septum is separated from the annulus by the medial atrial wall. The topography of the AV node was also presented (49, 51, 60). With the use of three planes of sections, it was demonstrated that ordinary myocardial tissues, the circumferential and perpendicular laminae, form the inferior-most region of the atrial wall around the valve. These tissues and the pectinate muscles form a muscular valvular apparatus (53, 61). The present study was undertaken as an essential part of a series of correlated electrophysiological investigations that showed that the atrionodal bundles and proximal AV bundle are part of a sinoventricular conduction system (49, 50, 53). The aim of ongoing investigations is to understand mechanisms underlying the role of the atrionodal bundles and proximal AV bundle in AV node function (57, 62). The purpose of this Part II study was to determine the relationship of the atrionodal and proximal AV bundles to the ordinary myocardium and intracardiac structures and the morphology, cytoarchitecture, and interrelationships of all structures within the AV junction region. A preliminary report of the results has been made (59). MATERIALS AND METHODS Twenty-one mongrel dogs weighing 9–15 kg were anesthetized with pentobarbital sodium (30 mg/kg iv). The Animal Care and Use Committee of Northwestern University approved the protocols for animal and tissue use. Preparation of the hearts was as previously described (55). In brief, the hearts were excised and the right atrium was exposed by using a ventrolateral incision through the AV groove, ventrolateral atrial wall, and superior vena cava. Each heart was flattened naturally by placing it on a paper towel, endocardial side down, in a dish to which Karnovsky’s fixative was added; 5% sucrose buffer was used for pre- and postfixation rinses. Photographs were The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 0363-6135/04 $5.00 Copyright © 2004 the American Physiological Society H1853 Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 Racker, Darlene K. The AV junction region of the heart: a comprehensive study correlating gross anatomy and direct threedimensional analysis. Part II. Morphology and cytoarchitecture. Am J Physiol Heart Circ Physiol 286: H1853–H1871, 2004; 10.1152/ ajpheart.01205.2003.—This “Part II morphology and cytoarchitecture” study is based on paraffin-embedded specimens in which the extracellular and intracellular matrix are preserved; single parallel, perpendicular, and transverse serial sections of the entire atrioventricular (AV) junction region (AVJR) and their correlation with photographs of the tissue blocks. As in Part I, the same major new findings are: 1) a coronary sinus fossa is formed by the superoposterior right medial atria wall (MAW), the left atrium, and the coronary sinus roof; 2) the posterior MAW forms two myocardial bridges and is isolated from the sinus venarum by the floor of the inferior vena cava; 3) the tendon of Todaro terminates in the superior lip of the coronary sinus ostium; 4) only ordinary myocardium contacts the annulus fibrosus, and there is little to no collagen separating its myofibers and tissues; 5) the ventricular septum shoulder is humped shaped, completely overlaid by annular myocardium, and joined by struts of papillary muscle; 6) the membranous septum joining the ventricular septum shoulder to the crista supraventricularis forms part of the aortic valve sinus walls; and 7) myocardium of the atrionodal bundles is aggregated into numerous small fascicles encased by collagen and is outside of the MAW as are the other specialized tissues. The proximal AV bundle and medial atrionodal bundle are aligned to the medial leg of Koch’s triangle and the tendon of Todaro. These data show, therefore, that the AVJR contains two overlapping atrial circuits. In the MAW, acivation of the posterior region is delayed because of the two myocardial bridges. Puncture of the AVJR can produce communication with an extracardiac space, posteriorly and medially, and with the aorta, anteriorly. H1854 AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE made from the endocardial and epicardial surfaces of the hearts during various stages of flattening and blocking. The AV junction region yielded one to three blocks for sectioning, depending on heart size and section plane. Eleven hearts were sectioned at a thickness of 10–15 m in either of three orthogonal planes (see bottom panel in Fig. 1), parallel to the endocardial surface (5 hearts), perpendicular to (4 hearts), or transverse to the long axis of the AV junction region (2 hearts) (see Table 1 in Ref. 55). All sections were mounted on glass microscope slides and selected sections were stained with one of the following: Goldner Trichrome, Mason Trichrome, Verhoff Elastica or hematoxylin-phloxine-safran. Photomicrographs of histological sections were taken at intervals of 0.5 mm for transverse and perpendicular sections and 0.25 mm for parallel sections. Additionally, every section was stained and photographed at selected sites. Photomicrographs of a 0.1-mm ruler, taken at the same magnification, were used for measurements within photomicrographs of the histological sections. Photographs of 10 additional hearts in various stages of dissection described in Part I (55) were studied compared with photographs of the gross hearts used for orthogonal sectioning and photomicrographs of the serial sections. Abbreviations of new terms are listed in Table 1. Data Analysis The size of structures was determined in millimeters by using section thickness and the number of serial sections in which each structure appeared, by direct measurements using an ocular micrometer, and from photomicrographs. The relationships between structures and their topology were determined by direct comparison of photomicrographs in the three planes of sections and photographs of the epicardial and endocardial surfaces of whole and dissected hearts and tissue blocks. RESULTS Figures 2, 3, and 4 (top inset) show representative endocardial and epicardial exposures of the AV junction region. The AV junction region extends from the superior and inferior borders of the coronary sinus ostium to the anterior edge of the medial leaflet. The posterior border of the coronary sinus ostium is the posterior atrial wall. For this systematic serial section study, the AV junction region was divided into three divisions according to endocardial topoAJP-Heart Circ Physiol • VOL graphic landmarks, each ⬃6 mm in length: 1) a posterior region that coincides with the length of the coronary sinus ostium and forms the right wall of the “coronary sinus fossa” superiorly and borders the posterior medial commissural leaflet inferiorly; 2) a midregion that coincides with the posterior half of the medial leaflet and is inferior to the fossa ovalis with atrial septum; and 3) an anterior region that coincides with the anterior half and edge of the medial leaflet (see Fig. 2E in Ref. 55). Figures 2–4 show serial sections of the AV junction region in the three orthogonal planes at regular intervals of ⬃0.25 mm for parallel (Fig. 2) and perpendicular (Fig. 3) planes, larger intervals especially for the transverse plane (Fig. 4), or smaller intervals when necessary to capture alterations in relationships or the morphology of specific tissues. Goldner Trichrome was found to be the most useful stain, because the tissues were stained differentially, as follows: 1) myocardium, brick red; 2) connective tissue, blue-green; 3) nerves and ganglia, magenta; 4) red blood cells, orange; and 5) nuclei, black. Primarily, a low magnification light microscope exploration of the serial sections was chosen for this study to establish the morphology or shape of the tissues and their interrelationships. Specialized Conduction Tissues of the AV Junction Region Atrionodal bundles. The medial, lateral, and superior atrionodal bundles are shown for the first time to be associated with epicardium of the medial atrial wall and the crest of the ventricular septum and ⬃1 cm away from the annulus fibrosus. The myofibers are packaged into small fascicles ensheathed by collagen, and the numerous fascicles forming each tissue are surrounded by collagen. Elastic fibers are restricted to the walls of large blood vessels. The myocytes are ⬃6 m in diameter, possess uniform cross striations, and are joined by intercalated disks that appear to be more prominent and more numerous compared with those in the proximal AV bundle, AV node, and distal AV bundle (58). No cell-to-cell connections were found with the ordinary atrial myocardium. 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 Fig. 1. Top: schematic of the hypothesized sinoventricular conduction system (from Fig. 1 in Ref. 55, modified with permission from Wiley-Liss, Inc.). The superior (SAB), medial (MAB), and lateral (LAB) atrionodal bundles, proximal atrioventricular (AV) bundle (PAVB), AV node (AVN), and distal AV bundle (DAVB) (solid lines) were traced from a single parallel section. The sinoatrial (SA) node (SAN) and its extensions (dashed-lines) were demonstrated in whole rabbit hearts stained for cholinesterase activity by Bojsen-Moller and Tranum-Jensen (10). Note that the LAB and MAB and the tail or caudal extensions of the SA node overlap. The fossa ovalis (FO) and orifices of the superior (SVC), inferior vena cavae (IVC), and coronary sinus (CS) are shown for orientation. Bottom: schematic diagram of the parallel, perpendicular, and transverse orthogonal section planes. BB, bundle branch. AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE Table 1. Abbreviations, and definitions of new terms and previous nomenclature Abbreviations A-IVS Groove Definitions Previous Nomenclature MAW im, sm MAW ip MAW sp MAW infero- superomida MAW inferoposteriora MAW superoposteriora n noncoronary posterior aortic valve sinus wall Proximal AV bundlea PAVB-AVN junctiona Posterior atrial walla posterior medial commissurral leaflet Perpendicular laminaa right aortic valve sinus wall Superior atrionodal bundle Superior vena cava Tendon of Todaro Ventriculoaortic septuma Ventricular septum papillary musclea Ventricular septal crista supraventricularis bridgea AAS ALFO Ao AVN AVN-DAVB Junct CFB CL CLn CS Fo CSO DAVB FO IVC LA LAB Ma PAVB PAVB-AVN Junct PAW PCML PL r SAB SVC TT VAS VS pap VS-CtSV AV groove None Same Same Tawara’s AV node None Same None None None Same His or AV bundle Same Same Same None Same Same None Atrial septum None Anterior septal region, muscular AV septum, medial atrial wall Atrial septum Septal isthmus Eustachian ridge, sinus septum, sinus band, posterior septal region Same Tawara’s atrial bundle None Caval isthmus Same None Same None Same Same None None None a New anatomic terms from Ref. 55, Table 2 (The Anatomical Record copyright 娀1999, Wiley-Liss, Inc.). MEDIAL ATRIONODAL BUNDLE. The medial atrionodal bundle is associated with the medial aspect and superior lip of the coronary sinus ostium, is subjacent to epicardium of the superoposterior medial atrial wall, and is opposed to the medial aspect of the tendon of Todaro (Fig. 4, C–F). Collagen separating individual fascicles is clearly seen in transverse (Fig. 4, C–F) and parallel (Fig. 2, J, K, L–M⬘) sections. The collagen AJP-Heart Circ Physiol • VOL sleeve enclosing the bundle is most apparent in parallel sections (Fig. 2M⬘, arrows). In transverse sections, stacked profiles of fascicles comprising the bundle are clearly seen adjacent to the medial aspect of the coronary sinus wall (Fig. 4, C–F). The trajectory of the medial atrionodal bundle along the epicardium of the superoposterior medial atrial wall is to the terminal right limb of the crista terminalis at the level of the superior lip of the coronary sinus ostium and the posterior atrial wall. LATERAL ATRIONODAL BUNDLE. The lateral atrionodal bundle is associated with the lateral aspect of the coronary sinus ostium, and, as shown here for the first time, is subjacent to and restricted to, the inferior lip and floor of the coronary sinus ostium (Fig. 4, E–F). In Fig. 4, E–F, the bundle is subjacent to the epicardium of the uppermost aspect of the inferoposterior medial atrial wall, is apposed to the crest of the ventricular septum, and is several millimeters away from the annulus fibrosus. A sinus that drains into the anteroinferior lip of the coronary sinus (Fig. 4, D–E) is seen in parallel sections (Fig. 2, G and H) and was found to be a common landmark that is helpful in locating the bundle in perpendicular (Fig. 3C) sections. Because this bundle takes a curvilinear path, its myofibers are often seen on their edges. More complete exposure of the lateral atrionodal bundle was made possible by partial dissection of the myocardium overlying the AV node (see Figs. 7–8 in Ref. 51). The trajectory of the lateral atrionodal bundle along the epicardium of the inferoposterior medial atrial wall is to the inferior terminal right limb of the crista terminalis at the level of the inferior lip of the coronary sinus ostium and the posterior atrial wall. SUPERIOR ATRIONODAL BUNDLE. The superior atrionodal bundle, as shown here for the first time, is subjacent to the base of the superoanterior medial atrial wall, deep to the narrowed region of the circumferential lamina as seen in sequential parallel sections (Fig. 2, I–J), and in single perpendicular sections (Fig. 3G). The bundle also is closely apposed to the crest of the ventricular septum (Fig. 4, I–K). In parallel sections, the bundle appears to join the AV node (Fig. 2J). However, perpendicular sections show clearly that it is separated by connective tissue from the AV node (Fig. 3F). The junction of the superior atrionodal bundle with the proximal AV bundle is shown in transverse sections (Fig. 4, I–K). The anterior extent of the superior atrionodal bundle runs in the superoanterior AV junction region, and is associated with the crest of the central fibrous body that separates it from the AV node (compare Figs. 2J and 3F). The collagen sleeve of the superior atrionodal bundle is seen in parallel sections (Fig. 2J⬘) but is more apparent in perpendicular (Fig. 3H) and transverse sections (Fig. 4K). Collagen separating single fascicles within the bundle is clearly visible in parallel sections (Fig. 2J⬘). The trajectory of the superior atrionodal bundle is toward the Bachmann bundle on the epicardial aspect of the medial atrial wall at the superior limit of the septal raphe. Here, the Bachmann bundle extends superiorly from the sinoatrial (SA) node and left limb of the sulcus terminalis en route to the left atrium. CYTOARCHITECTURE OF ATRIONODAL BUNDLES. Figure 5 is representative of the cytoarchitecture of the atrionodal bundles, as shown here for the first time, which contain multiple small fascicles with gently spiraling parallel small myofibers (Fig. 5, A and B). The myocytes contain delicate uniform cross striations and are joined by prominent intercalated disks (Fig. 5C). 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 ML MAB MAW MAW ia MAW sa Atrial interventricular septal groovea Atrioaortic septuma Anterior limbus of the fossa ovalis Aorta Atrioventricular node AV node-distal AV bundle junctiona Central fibrous body Circumferential laminaa Circumferential lamina, narrow regiona Coronary sinus fossa Coronary sinus ostium Distal AV bundlea Fossa ovalis Inferior vena cava Left atrium Lateral atrionodal bundlea Mitral valve anterior leaflet Medial leaflet Medial atrionodal bundlea Medial atrial walla MAW inferoanteriora MAW superoanteriora H1855 H1856 AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE central fibrous body (gray region encasing node in Figs. 2F; 3, G and H⬘–L; and 6, B and C3). Little fat and no large blood vessels are present (Figs. 2, F–L; 3, B–L; and 4, M–O). The interwoven nature of fascicles comprising the AV node as well as their collagen encasement and small size are apparent in parallel (Fig. 2J⬘) and perpendicular (Fig. 3, B⬘, E, F, and H⬘) sections. In transverse sections, the AV node is a dense knot of myofibers that appears to have little to no connective tissue between its myofibers at low power (Fig. 4, M–O). However, at medium and high power and in perpendicular sections it is clearly seen that continuous ribbons of collagen extends from the proximal AV bundle and through the AV node and the distal AV bundle (Fig. 3, F, H, and H⬘). Distal AV bundle. The distal AV bundle extends from the AV node anteriorly ⬃3 mm to its branch point at the anterior edge of the medial leaflet (Figs. 2, E–M and 4, P–Q). It is also delineated here for the first time to reside within the anterior part of central fibrous body in which it appears to penetrate the so-called septum fibrosum, the dense fibrous tissue floor of the central fibrous body (Figs. 2, E–M and 4, P–R). This denser region of the central fibrous body is seen to envelop the bundle along its full extent in parallel sections (Fig. 2, E–M). The distal AV bundle is also subjacent to the inferoanterior medial atrial wall and is apposed to the upper shoulder of the ventricular septum posteriorly (Fig. 4, P–Q) but attains the crest of the ventricular septum anteriorly as it branches (Fig. 4, Q–R). Right bundle branch. The right bundle branch continues from the distal AV bundle at the level of the atrioaortic septum (Figs. 3M and 4R). This septum is, in fact, part of the sinus walls of the aortic valve (Fig. 2M) and, as also shown here for the first time, is continuous with the ventriculoaortic septum, which is the wall of the right coronary sinus as shown in Part I (see Fig. 7B in Ref. 55). Superior Medial Atrial Wall in the AV Junction Region The superior division of the AV junction region is shown here for the first time to be part of the superior division of the medial atrial wall as displayed clearly in transverse sections. The superoposterior medial atrial wall forms the right wall of the “coronary sinus fossa,” and its epicardium abuts the medial atrionodal bundle (Fig. 4, C–F). The superomid medial atrial wall is narrow and runs inferior to the fossa ovalis (that Fig. 2. The AV junction region in orthogonal parallel plane serial sections. Top, representative tissue block with centimeter ruler. Endocardial aspect (left) displays orifices of the IVC and CS (arrows), the medial (ML) and posterior medial commissurral (PCML) leaflets, and location of the AV node (black dot). Epicardial aspect (right) displays the aortic valve right (r) and posterior noncoronary (n) sinus walls, a remnant of the left atrial endocardium (LA), IVC, and a string protruding from CS. Medial atrial wall (MAW)sp, -sm, and -sa indicate the superoposterior, superomid, and superoanterior regions, respectively, of the medial atrial wall. Sectioning commenced parallel to the endocardium. Selected serial sections are shown with interval spacing listed in millimeters (top, right, corner). Calibration bar for A–M is in A (top, center), J⬘ (bottom, left), M⬘ (top, right). A: medial leaflet. B: tendon of Todaro (TT) spans MAWsp and MAWsm. Atrial-interventricular septal (A-IVS) groove separates MAW from ventricular septum (VS) and roof of the central fibrous body (CFB). C: inferior circumferential lamina (CL) myofibers inserting into connective tissue. D: first of AVN and DAVB are deep to CFB roof. CL myofibers continue to terminate near AVN. E: first of PAVB. F: stack of CL myofibers insert into connective tissue along the VS hump-shaped shoulder; VS crest emerges, surrounded by collagen. G–H: small sinus (CS, black dot) drains into the inferoanterior CS wall, adjacent to LAB in H, and the first of the narrowed CL (CLn) region is superior to AVN. CLn is ⬍0.5 mm thick in H and I, but is absent in G and J. I: LAB. J: first of the MAB and SAB. CL, inferior to CS, is being replaced by connective tissue. J⬘: from box in J. SAB myofibers and its collagen sleeve (white arrows and arrowheads) pass over AVN en route to the PAVB. Parallel fascicles of PAVB-AVN and AVN-DAVB (Junct) junctions and tightly knotted AVN fascicles. K: SAB is absent. L: VS is a narrow bar of tissue several millimeters inferior to CS (see Fig. 4B). DAVB gives way to branch point (BP) and branching bundle at the anterior orifice of CFB. M: AVN has been replaced by the floor of CFB. M⬘: from box in M. MAB with collagen sleeve (black arrows). See Table 1 for other abbreviations. Goldner Trichrome stain. AJP-Heart Circ Physiol • VOL 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 Proximal AV bundle. The proximal AV bundle extends from the AV node posteriorly to the floor of the coronary sinus ostium and, as shown here for the first time, is associated with the superomid medial atrial wall epicardium, the medial aspect of the tendon of Todaro (Fig. 4, G–K), and the posterior half of the medial leaflet (Fig. 2, A, J–M and inset). Definitively, the transverse plane also shows for the first time that the bundle is ⬃1 cm away from the hinge point of the leaflet at the annulus fibrosus (Fig. 4G), is associated with only the crest and not the shoulder of the ventricular septum (Fig. 4, G–K), and runs at the level of the left atrium (Fig. 4G). A characteristic of the proximal AV bundle at its ventricular septal apposition is the presence of numerous fat cells (insets in Figs. 4, F, H, and J and 6A). The association of the proximal AV bundle with the medial aspect of the tendon of Todaro is not observable in parallel and perpendicular sections, because the tendon is sectioned through before the proximal AV bundle is attained (compare Fig. 2, B and C with F–M). The proximal AV bundle, the tendon of Todaro, and the crest of the ventricular septum appear to make almost a 45-degree angle with respect to the fibrous annulus, the AV node, and the coronary sinus ostium. This can be seen directly in parallel sections (Fig. 2J) but more clearly in successive transverse sections in which the AV node is seen near the hinge point of the medial leaflet (Fig. 4N), and the proximal AV bundle is at the crest of the ventricular septum (Fig. 4G). Proximal AV bundle-AV node junction. The acute angling of the proximal AV bundle forms an L-shape just before its junction with the superior atrionodal bundle (Fig. 4H, inset, and I–K). The short leg of the “L” is equivalent to the region of its acute angling and also the proximal AV bundle-AV node junction seen in parallel (Fig. 2, J⬘–K) and perpendicular (Fig. 3E) sections. Amazingly, parallel (Figs. 2J⬘ and 6A, inset), perpendicular (Figs. 3, E and H⬘ and 6B), and transverse sections (Fig. 4, H and J) show that all myofibers joining the atrial end of the AV node do so via this narrow bridge of tissue, ⬍0.5 mm in width. AV node. The AV node is shown topographically here for the first time to coincide with the inferoanterior region of the medial atrial wall (Fig. 4, M–O), to extend ⬃3 mm from the base of the middle medial leaflet anteriorly (Figs. 2, F–L and 3, B–L) to be closely apposed to the shoulder of the ventricular septum (Fig. 4, M–O) and to be in the posterior part of the AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE H1857 Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 AJP-Heart Circ Physiol • VOL 286 • MAY 2004 • www.ajpheart.org H1858 AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE coincides with the atrial septum) and is superior to the posterior half of the medial leaflet; its epicardial aspect forms the last vestige of the coronary sinus fossa and is tucked inside the heart where it abuts the proximal AV bundle (Fig. 4, G–L). The superoanterior medial atrial wall in the AV junction region contains terminal myofibers of the anterior limbus of the fossa ovalis, which is a prominent dense mass of myocardium (Fig. 4, M–O). or distal AV bundle. However, when these myocytes are cut lengthwise and on edge, as is the case of the perpendicular lamina myofibers in transverse sections (Fig. 4, N–O), the size of the myocytes is particularly misleading, because only the rectilinear long edge of the myocytes and myofibers may be seen. This same phenomenon holds true for the specialized myofibers, that is, the apparent size of the myocyte or myofiber depends on the plane of section. Inferior Medial Atrial Wall: Circumferential and Perpendicular Laminae Coronary Sinus Fossa The coronary sinus fossa, a dimple located at the base of the heart at the crux where the four chambers meet (see Fig. 3 in Ref. 55), is shown for the first time as it extends into the heart aligned with the superoposterior and the superomid medial atrial wall, with the tendon of Todaro, and with the medial atrionodal bundle and the proximal AV bundle (Fig. 4, C–L, fibrous remnant not labeled in I–L). Central Fibrous Body The central fibrous body was described in Part I (55) for the first time to have an endocardial exposure at its anterior extent, the so-called septum fibrosum that surrounds the distal AV bundle. The endocardial exposure can be seen in perpendicular sections (Fig. 3M) but to best advantage in transverse sections (Fig. 4R), and its anterior ostium coincides with the distal AV bundle at the branch point (Fig. 4R). Parallel sections show that the entire floor of the central fibrous body, as it underlies the AV node and distal AV bundle, is comprised of dense connective tissue, and the roof is comprised of less dense connective tissue (Figs. 2, E–M and 3I). The dense and thin regions of the central fibrous body are best seen in parallel sections (Fig. 2E). The posterior half of the central fibrous body envelops the AV node as seen in parallel and transverse sections (Figs. 3H⬘ and 4, M–0) and by examination of serial parallel sections (Fig. 2, B and M). Tendon of Todaro The tendon of Todaro is shown here for the first time to be aligned sequentially with the medial atrionodal bundle in the superoposterior medial atrial wall (Fig. 4, C–F) and with the proximal AV bundle in the superomid medial atrial wall (Fig. 4, G–I). As the tendon approaches the superoanterior medial atrial wall, it is covered by atrial myocardium and is several Fig. 3. The AV junction region in orthogonal perpendicular plane serial sections. Top: endocardial (left) and epicardial (right) aspects of a representative tissue block with the vertical edges ruled in millimeters and calibration bar (from Fig. 2). Sectioning was perpendicular to endocardium in the plane of the annulus and began from the atrial side of the tissue block (solid black line). The section level for G is shown (dashed line). Selected sequential serial sections with interval between sections given is in millimeters (top, right, corner) and the calibration bar in millimeters for A–D, F–M is in A (top, left), for E and H⬘ is in E (bottom, center), and for inset in D, F, G, H, and K, is in D, inset (bottom, left). A: The A-IVS groove. B: PAVB and AVN. B⬘: from dashed lines in B, fascicles of AVN separated by collagen. C: CL’s thick and narrowed regions (arrows). Note, a sinus (black dot) drains into CS ostium (see also Fig. 2, G–H and 4, D–E). D: PAVB. Inset: from dashed lines, displays parallel fine caliber myofibers separated by collagen. E: CLn is superior to AVN. PAVB-AVN and AVN-PAVB Juncts display parallel fascicles. CLn myofibers are large and dense compared with those of the specialized tissues. F: first of the SAB. Inset: from box; SAB joins the PAVB (large white arrow) and is separated from the AVN (white arrows) by the CFB. G: LAB is deep to the inferior lip of the CS. Inset: from dashed lines, fine-caliber PAVB fascicles in parallel array are divided by a blood vessel. H: SAB; inset is from dashed lines inside box and shows that the CFB connective tissue separates the SAB from the AVN (black arrows). H⬘: from box in H; myofibers of the PAVB-AVN and AVN-DAVB junctions are parallel, and those of the AVN are interwoven. Ribbons of collagen separate the myofibers. I–J: SAB is absent. DAVB at its branch point (BP). K: LAB is present and AVN is a thin bridge. Inset: vacuolated LAB fascicle (arrow). L: Perpendicular lamina (PL) is sandwiched between endocardium and epicardium M: the only specialized tissues remaining are the DAVB and the right bundle branch (RBB). Note the atrioaortic aeptum (AAS). See Table 1 for other abbreviations. Goldner Trichrome stain. AJP-Heart Circ Physiol • VOL 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 From various connective tissue stains, it is shown for the first time that little to no collagen separates either the circumferential or perpendicular laminae or myofibers within the laminae. This is particularly apparent in parallel serial sections that display the subendocardial perpendicular laminae and the subepicardial circumferential laminae as two sheets of singly arrayed parallel myofibers running at right angles to each other (Fig. 7). Transverse sections of the inferoposterior medial atrial wall show the laminae as they appear in the anterior, lateral, and posterior regions of the ring, i.e., a thin, subendocardial, perpendicular lamina closely apposed to a robust, subepicardial, circumferential lamina (Fig. 4, A–B). Also delineated here for the first time: 1) only the annular myocardium is adjacent to the annulus fibrosus (Fig. 4, A–L); 2) the circumferential lamina thins, narrows, and directly overlays both the crest of the central fibrous body (Fig. 3E) and the superior atrionodal bundle (Figs. 2, H–I and 3, C and E). The thin, narrowed region is admixed with myofibers of the anterior limbus of the fossa ovalis (Fig. 4M) and in transverse section is not distinguishable as a discrete tissue; 3) only myofibers of a much thicker perpendicular lamina directly overlay the posterior part of the central fibrous body and are seen in the long axis in transverse (Fig. 4, M–O) and in cross section in perpendicular (Fig. 3L) sections; and 4) only myofibers of the perpendicular lamina contact the hinge point of the middle medial leaflet (Fig. 4N). Myocytes of both laminae exemplify cytologic characteristics of ordinary atrial myocardium. The parallel array and lack of collagen encasement of single myofibers as well as infrequent myocyte branching can be appreciated in parallel sections (Fig. 7D). The ordinary atrial myocyte is 15 to 30 m in diameter (Fig. 7D), more than twice that of myocytes in the atrionodal bundles (Fig. 5C), proximal AV bundle, AV node, AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE millimeters away from both the L-shaped proximal AV bundle-AV node junction (Fig. 4L) and the AV node (Fig. 4, M–O). Figure 4O shows the tendon near its origin from the central fibrous body. Figure 2, B and C shows the terminus and the midregion of the tendon in the superoposterior and the superomid medial atrial wall in single sections. Thus the tendon shields the proximal AV bundle (Fig. 4, G–K), the medial atrionodal bundle (Fig. 4, C–F), and the superior atrionodal bundle at its junction with the proximal AV bundle (Fig. 4, I–K) but does not shield the proximal AV bundle-AV node junction (Fig. 4L). H1859 Ventricular Septum and Papillary Muscles The crest and shoulder of the ventricular septum and the papillary muscle attachments were described in Part I (55). Here, it is shown for the first time that posteriorly the ventricular septum is a narrow bar of tissue (Fig. 2, K–M) as it joins the right ventricular free wall and is inferior to the coronary sinus ostium. Its hump shape rises abruptly to abut the coronary sinus ostium as seen in parallel (Fig. 2, J–M) and transverse (Fig. 4, A–B) sections. The transverse sections show that the bulk of the myocardium in the AV junction region is that Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 AJP-Heart Circ Physiol • VOL 286 • MAY 2004 • www.ajpheart.org H1860 AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE of the ventricular septum, that the AV node and part of the distal AV bundle (both within the central fibrous body) are apposed to the shoulder of the ventricular septum (Fig. 4, M–P), that the distal AV bundle gradually moves to the crest of the ventricular septum (Fig. 4Q) where it bifurcates (Fig. 4R), and that the medial and superior atrionodal bundles and the proximal AV bundle are apposed to the crest of the ventricular septum (Fig. 4, C–J). Direct Three-Dimensional Analysis of the AV Junction Region Certain details and interrelationships are apparent only by direct comparison of the three planes of sections as seen in Fig. 6 and delineated here for the first time as follows. A change in the axis of the conduction tissues occurs at the proximal AV bundle as clearly seen in serial transverse section Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 Fig. 4. The AV junction region in orthogonal transverse plane serial sections. Top: 2 tissue blocks used for sectioning with the endocardial (left) and epicardial (right) aspects, horizontal edges ruled in millimeters, and a calibration bar. Vertical lines indicate the location of sections in C, F, K, M, O, P, and R. The epicardial exposure shows left atrium endocardium and attached mitral valve leaflet (Ma) overlying the aortic valve. Selected serial sections are shown with interval between sections given in millimeters (top, right). The calibration bar for A–R is in A (top, left) and insets for F (lower right), H (upper right), J (lower right). C–F: shows the posterior MAWsp, -ip. G–L: the midMAWsm, -im, and the anterior M–Q. MAWsa, -ia in the AV junction region. A–B: PL myofibers are seen along the long axis. CL myofibers are in cross section. B: PL is thicker compared with PL in A. Ventricular septum papillary muscles (VS pap) are also seen. C: first of the MAB is seen subjacent to MAWsp and in the floor of the CS fossa (CSFo). MAB is associated with the superior lip of the CS ostium and TT. The A-IVS groove is traced by white arrows. CL is no longer apparent. D–E: first of the LAB is seen subjacent to the inferior lip of CS. A sinus draining into the inferoanterior lip of the CSO (black dot) is adjacent to LAB. F: the LAB, MAB, and PAVB convergence is set hard against the VS crest and is several millimeters away from the annulus fibrosus (AF). Inset: from dashed lines; shows finecaliber and vacuolated PAVB fascicles. G: PAVB is set hard against the VS crest. H–I: PAVB is L-shaped. MAWim is comprised of only PL near AF. Inset in H shows L-shaped PAVB (arrows) at high magnification. J–K: PAVB the SAB junction. Inset: J, from white arrowhead-hash marks; shows PAVB at high magnification. L: SAB is absent. M–O: PL myofibers are separated from AVN myofibers by the CFB. CFB is robust in O but a thin connective sleeve in M and N (white or small black arrow). PL myofibers extend into the leaflet. Myofibers from the anterior limbus of the fossa ovalis (ALFO) are in MAWsa. P–Q: DAVB within the CFB at VS crest. R: DAVB at BP and AAS are seen. Ao, aorta. See Table 1 for other abbreviations. Goldner Trichrome stain. AJP-Heart Circ Physiol • VOL 286 • MAY 2004 • www.ajpheart.org AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE (Fig. 6, C1–C4) but is not apparent in either parallel (Fig. 6A) or perpendicular (Fig. 6B) sections. Whereas evidence of the central fibrous body location of the AV node may be deduced from comparison of sections in the parallel (Fig. 6A) and perpendicular (Fig. 6B) planes, single transverse (Fig. 6C4) sections show that a halo of connective tissue completely encircles the node. The narrowed region of the circumferential lamina is superior to the roof of the central fibrous body with enclosed AV node (Fig. 6B) and only the perpendicular lamina myofibers cascade over the central fibrous body (Fig. 6C4). The superior atrionodal bundle may appear in transverse sections to be part of the perpendicular lamina as it cascades over the central fibrous body and AV node (Fig. 6C4). How- ever, parallel (Fig. 6A) and perpendicular (Fig. 6B) sections show clearly that these myofibers form a discrete bundle that is confluent with only myofibers of the proximal AV bundle. Comparison of the disposition of the specialized tissues in Fig. 6, A–C, to the right medial atrial wall confirms that the specialized tissues: 1) are all outside the right atrium and do not traverse the medial atrial wall; 2) make no myofiber-to-myofiber contact with the atrial or the ventricular myocardium; and 3) are isolated from the atrial and ventricular myocardium by collagen. The specialized tissues extending from the medial atrionodal bundle to the right bundle branch (Fig. 6A) run parallel to the upper or medial leg of the triangle of Koch (see Fig. 2 in Ref. 55), and only annular myocardium is associated with the lower leg of the triangle (Fig. 6, C1–C4). 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 Fig. 4 —Continued AJP-Heart Circ Physiol • VOL H1861 H1862 AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE DISCUSSION Systematic correlation of the gross anatomy in whole and dissected hearts and tissue blocks to the histology in three orthogonal planes of serial sections in Part I (55) and Part II studies demonstrated 1) the entirety of the medial atrial wall with its myocardial bridges and sheets of myofibers inferiorly forming the circumferential lamina, perpendicular lamina, and terminals of the anterior limbus of the fossa ovalis; 2) the fascicular and collagen composition of the continuum of AV junction region specialized conduction tissues; 3) the crest and shoulder of the ventricular septum and its papillary muscles; 4) the tricuspid medial and posterior medial commissurral leaflets; 5) the fossa ovalis-atrial septum relationship with the medial atrial wall; 6) the atrial-interventricular septal groove; AJP-Heart Circ Physiol • VOL 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 Fig. 5. Medial atrionodal bundle in parallel orthogonal plane sections. A: MAB parallel fascicles separated by collagen. B: from box in A, showing small myofibers comprising the fascicle. C: from box in B; myocytes possess evenly spaced cross striations and are joined by prominent intercalated disks. Calibration bar top, right of each panel. Other abbreviations are in Table 1. Goldner Trichrome stain. 7) the coronary sinus fossa; 8) termination of the tendon of Todaro in the superior lip of the coronary sinus ostium and its passage in the superomedial atrial wall; 9) the anterior and posterior central fibrous body regions; 10) the inferior vena cava isolation of the posterior AV junction region; 11) the anterior AV junction region association with the atrioaortic, ventriculoaortic, and ventricular septum crista supraventricularis bridge components of the membranous septum; and 12) the posterior AV junction region association with the posterior atrial wall. These features previously escaped detection because 1) a physiologic fixative protocol was needed to conserve delicate features of the intracellular and extracellular matrix; 2) flattening the heart was needed to expose structures caught in the curvatures of the AV junction region in histological sections; 3) three orthogonal planes of serial sections were needed to reveal specific details (e.g., see table in Ref. 58). Three planes of sections also confirmed the existence of the tissues. Some features of the above structures have been described by other investigators, but the significance and nature of their observations can only be made clear when the entire AV junction region is exposed. The following is a listing of the major contributors and their discoveries. Tawara (69, 70) described the proximal AV bundle and AV node with the AV node and AV bundle encased within the central fibrous body, the aggregation and connective tissue encasement of the myocardium into numerous fascicles, and the uniform cross striations. However, neither the proximal AV bundle, the atrionodal bundles, the arrangement of myofibers within the fascicles, nor the nerve supply for the AV node and distal AV bundle are demonstrated in Tawara’s data or noted in the text. Keith and Flack (33) and Bojsen-Moller and Tranum-Jensen (10, 72) described the AV junction region extensions of the SA node, the collagen encasement of these myofibers, and the uniform cross striations of the myocytes with the use of transmission electron microscopy (72). However, connections between the SA node extensions that are actually the atrionodal bundles, and that of the proximal AV bundle were not detected, nor was the proximal AV bundle distinguished from the AV node. Keith and Flack (33), Papez (46), and Baird and Robb (6) described circular myofibers with perpendicular myofibers offshoots but did not realize that the myofibers actually form distinct tissues (the circumferential and the perpendicular laminae) and constitute the inferior-most region of the atrial wall. Nor did they realize that the circumferential lamina myofibers merge with the pectinates, and thereby with the perpendicular lamina form the muscular valvular apparatus. Sweeney and Rosenquist (68) demonstrated the atrial septum as a blade-shaped structure incorporating the annulus inferiorly but did not realize that the atrial septum is limited to the inferior border of the fossa ovalis and therefore has no connection with the annulus. Walmsley and Watson (75) described only the anterosuperior region of the medial atrial wall but did not detect the inferior, middle, and posterior regions. Zimmerman and Bailey (78) described the tendon of Todaro as joining the cardiac skeleton but did not realize that the tendon terminates in the superior lip of the coronary sinus ostium. AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE H1863 McAlpine (38) demonstrated the supravalvular lamina of the anterior, lateral, and posterior atrial walls but did not realize that the lamina extends into the medial atrial wall and is actually comprised of the circumferential and perpendicular laminae or that the circumferential myofibers merge with the pectinates. James (29) showed that the crista supraventricularis joins the aortic wall but did not realize that it joins the membranous septum and thus forms the ventricular septum crista supraventricularis bridge. Sonnenblick et al. (66) demonstrated the muscular basis for valve function, i.e., the sphincter-like closure of the valve and contracture of the leaflets, but did not know that the structural basis for these actions were the circumferential and perpendicular laminae, respectively. AJP-Heart Circ Physiol • VOL Hoffman and colleagues (12, 20, 21) created atrial flutter in the right atrium using a Y-shaped incision model in canine heart, not knowing the existence of the muscular valvular apparatus or why transection of the valve terminated and precluded inducement of the arrhythmia. Hoffman and colleagues (16, 73) demonstrated electrical activity in the SA and AV nodes and the crista terminalis during atrial standstill in high potassium, and Dinari and Aygen (17) documented sinoventricular transmission during atrial standstill with hyperkalemia, not knowing the existence of the sinoventricular conduction system. Because an accurate knowledge of the anatomy is key to understanding the function, pathophysiology, and development of safer treatment modalities, etc. (the list continues), perhaps we should rethink what has been considered as dogma. Also, 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 Fig. 6. AV junction region by direct threedimensional analysis of selected orthogonal plane serial sections (from Fig. 6 in Ref. 55, The Anatomical Record copyright 娀1999 Wiley-Liss, Inc.). A: parallel plane shows the continuum of specialized tissues that spans the AV junction. Inset: from box, the PAVBAVN Junct is ⬍1 mm in width. B: perpendicular section made at the level of the CSO shows by the gray hues that specialized tissues are heavily invested by collagen and are within the A-IVS groove. Thick and thin regions of the circumferential lamina are traced by arrows. C1–C5: transverse serial sections at intervals are indicated by millimeters at top of each panel. Note that the A-IVS groove (black two-way arrows in C1) is quite extensive, because it represents the region of direct apposition of the MAW with the shoulder and crest of VS in C2–C5. Calibration bar spans bottom of A and B in inset of A, and for C1–C5 is in C1. See Table 1 for other abbreviations. Goldner Trichrome stain. H1864 AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE because the new structures are not seen in conventional data, the anatomy does not change, and every aspect of the anatomy is critical to functioning of the heart, it is of utmost importance to consider differences between the new and conventional data. Topography of AV Junction Region Components Results of the present study clearly show that structures within the AV junction region share a regular relationship with each other and with gross anatomic landmarks that do not vary, regardless of the size of the heart. Thus there is a dedication of structures within the superior, inferior, posterior, and middle and anterior AV junction regions. The topographic dedication of intracardiac structures is exemplified by the common appearance of a small sinus in the anteroinferior, lower lip of the coronary sinus ostium in all three planes of section (Figs. 2, G and H; 3C; and 4, D and E) in three different hearts. This small sinus has provided great utility in localizing the lateral atrionodal bundle in the in vitro whole heart for correlated functional studies (57, 62). The base of the medial leaflet of the tricuspid valve was also found to be useful in identifying the AJP-Heart Circ Physiol • VOL location of the specialized tissues for in vitro studies, because the anterior half is associated with the AV node and distal AV bundle, whereas the posterior half is associated with the proximal AV bundle (Figs. 2–4). The conserved position of the aortic valve cusps, membranous septum, and fibrous tissues and their relationships to the AV node were noted by Tawara (Ref. 67, p. 191), and demonstrated schematically (see Figs. A and B in Ref. 70) for different species and in young and old hearts. The length of the AV node [3 mm in the dog (see p. 18 in Ref. 67)] is directly confirmed by parallel and perpendicular sections using the scale in Fig. 6, A and B. The current beliefs that “Thus, the AV junction varies considerably from heart to heart, and no two AV junction regions are identical, especially in humans.” (Ref. 8, p. 228) are at odds with data of Tawara (67, 70) and the data herein. Medial Atrial Wall and the AV Junction Region Excision of the left atrium showed the limits of the medial atrial wall (49, 51, 60). The three orthogonal planes of serial sections herein demonstrated that in the AV junction region, 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 Fig. 7. Bilaminated annular myocardium in parallel orthogonal plane serial sections of the inferoposterior medial atrial wall. A: radially oriented myofibers of PL are interrupted due to curvature of the heart. B: from a section deep to A, PL myofibers (white arrows) appears to merge with CL myofibers (black arrows). C: from a section deep to B, CL myofibers extend from the inferior lip of CSO to the annulus. D: high magnification from a region similar to box in C shows little or no collagen separating myofibers and myofibers are singly arrayed. Calibration bar in B for A–C. See Table 1 for other abbreviations. Goldner Trichrome stain. AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE AJP-Heart Circ Physiol • VOL extends the extracardiac approach of the medial atrionodal to the proximal AV bundle (Fig. 6, C1 and C2) and has found utility in recording extracellular potentials using the extracardiac approach (5). Superoanterior AV junction region and superoanterior medial atrial wall. The superoanterior medial atrial wall of the superoanterior AV junction region runs in tandem with the anterior half of the medial leaflet and contains the origin of the tendon of Todaro (Figs. 4, M–R; 2, C and E; and 5 in Ref. 55). The superoanterior medial atrial wall provides an extracardiac approach to the superior atrionodal bundle and the AV node. The latter is marked topographically by an epicardial fat pad as demonstrated in gross specimen definitively using an AV node marker pin and serial histological sections (see Figs. 1, and 3, A–C in Ref. 51). These findings have also proven to be of great utility in localizing these tissues in functional studies by Hirao et al. (22). It is surprising that although the superoanterior medial atrial wall had been described by Walmsley and Watson (75) as the medial atrial wall in 1966, it was still designated as part of the atrial septum in histological sections and schematics by Anderson et al. in 1974 (see Fig. 3 in Ref. 4), Anderson et al. in 1975 (see Fig. 5 in Ref 2), and more recently by James et al. in 1996 (see Figs. 4, 6A, and 10 in Ref. 31), Mazgalev et al. in 2001 (see Fig. 7 in Ref. 37), Medkour et al. in 1998 (see Fig. 1 in Ref. 42), Waki et al. in 2000 (see Figs. 5–6 in Ref. 74), and Zhang et al. in 2001 (see Fig. 2B in Ref. 77) and in mapping studies of whole hearts (see Fig. 6 in Ref. 4, Fig. 4 in Ref. 5, and Figs. 5 and 6 in Ref. 45). Tendon of Todaro The tendon is an important marker for the proximal AV bundle in transverse section (Fig. 4). This landmark makes clear that the proximal AV bundle is not delineated in current histological studies, because the relationship with the tendon is directly apparent only in the transverse, frontal plane section, which is the plane of choice of conventional studies; yet histological sections of the middle and posterior regions of the tendon are seldom, if ever, included in the works cited herein. The tendon is also a useful marker for the atrionodal bundles junctions with the proximal AV bundle (midregion of the tendon) and the lateral and medial atrionodal bundles junction with the proximal AV bundle (anterior border of the coronary sinus region of the tendon). Finally, the likely function of the tendon is to maintain patency of the coronary sinus ostium, because the tendon terminates in the superior lip of the coronary sinus ostium (Figs. 2A and 4C). Central Fibrous Body The entirety of the central fibrous body was shown herein in parallel sections to form a “channel” or “tunnel” that encases the distal AV bundle, anteriorly, and the AV node, posteriorly (Fig. 2, F–M). The point of transition between the AV node and distal AV bundle within the central fibrous body is marked by a thick connective tissue collar for the distal AV bundle, as seen in parallel sections (Fig. 2J), but as a thick roof for the distal AV bundle in transverse sections (Fig. 4, P and Q) as sketched by Tawara in 1906 (see Figs. 4 and 5 in Ref. 70). The posterior extent of the central fibrous body, which spans the mitral and tricuspid valve annuli (the right fibrous trigone) 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 only ordinary atrial myocardium forms the medial atrial wall (Figs. 2–4). However, the various medial atrial wall divisions are still referred to as atrial septum. It is important that the atrial septum has two endocardial aspects, whereas a wall has an epicardial and endocardial aspect. It may also be surprising to find that the bulk of the AV junction region is ventricular myocardium. It is important, therefore, to consider that cells harvested from an excised AV junction region tissue block will have a preponderance of myocytes other than the specialized tissues as intended, e.g., Yuill and Hancox (76). It is of note that the mass of ventricular septum is greater than that of the ordinary atrial and papillary muscle, and the specialized tissues are of a minute population. It will also be important to distinguish anatomic markers for the various myocyte types, or alternatively, single cells could be isolated from specific tissues, because it is known that ordinary atrial myocardium can be perturbed to elicit electrical potentials with characteristics of specialized tissues. Superoposterior AV junction region and superoposterior medial atrial wall. Histological data showed that atrial myocardium of the posterior AV junction region forms the “posterior” medial atrial wall, and the superoposterior medial atrial wall contains the termination of the tendon of Todaro in the upper lip of the coronary sinus ostium (Fig. 4). The superoposterior medial atrial wall was also shown to run in tandem with the coronary sinus ostium, to be the right wall of the coronary sinus fossa by transection of the fossa, and to be isolated from the sinus venarum and midcrista terminalis by the inferior vena cava (Fig. 2, A–F, left, upper corner) (see Figs. 3 and 5 in Ref. 55). Thus because the inferior vena cava borders the full extent of the superoposterior medial atrial wall (see Fig. 3, E–G, in Ref. 55), this region of the wall has no contact with the atrial septum or, importantly, with the sinus venarum. The superoposterior wall in dog heart is similar to that of human heart, as viewed epicardially by McAlpine (see Fig. 68–3 in Ref. 38) and endocardially by Anderson and Becker (see Figs. 2.8 and 2.11 in Ref. 1) and by McAlpine (see Figs. 97–3 and 99–3 in Ref. 38). The superoposterior region, however, continues to be referred to as the sinus septum, the Eustachian ridge, the septal band, and the atrial septum. The inferior vena cava ostium is also depicted as part of the atrial septum in mapping studies reviewed by Janse and Anderson (see Fig. 5 in Ref. 32) and by Boyden (for review, see Fig. 6 in Ref. 11) via isochronal activation lines assigned to this region. Correlated anatomic electrophysiological studies are needed to resolve this issue, because it is known that cardiac myocardium does not reside within the inferior vena cava orifice. In addition, the inferior vena cava was shown to form a complete line of block along the superoposterior medial atrial wall (see Fig. 3, E and F in Ref. 55). Superomid AV junction region and superomid medial atrial wall. The superomid medial atrial wall and superomid AV junction region run inferior to the fossa ovalis (see Figs. 2, C and E and 5 in Ref. 55) that isolates it from the sinus venarum, and also contain the midregion of the tendon of Todaro. The epicardial expanse is the last vestige of the coronary sinus fossa (Fig. 4, G–L). Demonstration of the physical limits of the atrial septum (see Fig. 1 in Ref. 51) is key in understanding the extent of the medial atrial wall, and discovery of the coronary sinus fossa was key to delineating the middle medial atrial wall epicardium (see Fig. 3 in Ref. 55). This region of the wall H1865 H1866 AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE Topographic Criteria for Specialized Tissues of the AV Junctional Region There are three topographic criteria for the AV junction region-specialized continuum. First, it is located outside the medial atrial wall. Second, each tissue has a dedicated location. The medial and lateral atrionodal bundles are located at the superior and inferior lips of the coronary sinus ostium. The proximal AV bundle extends from the anterior border of the coronary sinus ostium to the posterior half of the medial leaflet, inferior to the fossa ovalis. The AV node and distal AV bundle, each measuring ⬃3 mm in length (Fig. 6, A–C), extend from the anterior half to the anterior edge of the medial leaflet. The superior atrionodal bundle runs inferior to the narrowed circumferential lamina and superior to the proximal AV bundle-AV node junction (Fig. 6B). Third, the specialized tissues are aligned with the medial or atrial leg of the triangle of Koch (see Fig. 2C in Ref. 55, and Fig. 2A in Ref. 58). Histological Criteria for Specialized Tissues of the AV Junctional Region The specialized tissues are shown herein to have four histological characteristics: 1) a grouping of a few parallel myofibers into small fascicles, 2) an encasement of the fascicles in collagen, 3) numerous small fascicles forming each tissue, and AJP-Heart Circ Physiol • VOL 4) myofibers containing uniform cross striations. Characteristics 1–3 distinguish the specialized tissues from the working atrial myocardium. However, each specialized tissue exhibits distinguished features: 1) myocytes with finger-like processes and thin branches were not found in the atrionodal bundles and are most pronounced in the AV node; 2) fascicles of nerve endings are found in the proximal AV bundle and AV node; 3) myofibers and fascicles exhibit distinct arrangement in each tissue such that a) atrionodal bundle myofibers and fascicles run parallel, b) proximal AV bundle myofibers are tightly coiled and form fascicles that run parallel, c) AV node myofibers run parallel, but the fascicles are wildly intertwining, d) distal AV bundle myofibers and fascicles run parallel as in the atrionodal bundles, but the myofibers are larger; 4) there is an abrupt ordering of both fascicles and myofibers at junctions between the specialized tissues, thereby distinguishing one tissue from the other at the high-power light microscope level; 5) myocytes have clear perinuclear regions and are largest in the distal AV bundle; and 6) intercalated disks are more prominent in the atrionodal bundles than in the proximal AV bundle and node (Ref. 56, see Figs. 3D and 4D in Ref. 58). The foregoing corroborate criteria established by Tawara for hallmarks of specialized tissues (Ref. 67, p. 152), i.e., collagen encasement of small clusters of myofibers with uniform cross striations, clear perinuclear regions (see Figs. 1–3 in Ref. 67). The collagen envestment can be discerned in parallel and transverse sections but is most conspicuous in perpendicular sections in which the tissues appear pale compared with the overlaying atrial and underlying ventricular myocardium (Fig. 3B, E–G). To make this distinction clear, Tawara indicated in the text (see p. 22 in Ref. 67) and by carefully colored figures that the specialized tissues were orange because of the mixture of red connective tissue and yellow myocardium staining by the Van Geison method. By contrast, intracellular and extracellular components of the tissues are not discernible in conventional high-power light microscope data, e.g., Becker and Anderson (see Fig. 10 in Ref. 7), James (see Figs. 3 and 12 in Ref. 27), Lev and Bharati (see Fig. 5–5B in Ref. 34), and Anderson and Ho (see Fig. 3 in Ref. 3). This loss of anatomic data accounts for differences reported for tissues between the various reports and the account herein. Atrial Approaches to the AV Node: Atrionodal and Proximal AV Bundles From the three orthogonal planes of sections, the atrial approaches to the AV node were shown to be restricted to fascicular extensions of the atrionodal bundles that approach the AV node via the proximal AV bundle (Figs. 2–5). Thus the atrial approaches provide a multilimb input to the AV node that is similar to its multilimb output, the distal AV bundle, and bundle branches. The atrionodal bundles are superior to and ⬃10 mm away from the annulus fibrosus as they join the proximal AV bundle, due to the acute angling of the proximal AV bundle (Figs. 4, D and K and 6). The proximal AV bundle is likewise ⬃10 mm away from the annulus (Fig. 6, C2 and C3). The elevated position and acute angling of the proximal AV bundle away from the fibrous annulus and the AV node and the minute size of the atrionodal bundles (Fig. 6C) and the PAVB-AVN junction (inset in Fig. 6, A and B), most likely 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 (Fig. 4, M–O), is joined medially by the tendon of Todaro (see Fig. 8 in Ref. 78), encases the AV node of Tawara as depicted in the 3-day-old dog heart by Tawara (see Fig. 3, Dog 120 in Ref. 70), and is analogous to the halo of connective tissue seen in transverse sections (Fig. 4O) and in Tawara’s monograph (see Fig. 3, Dog 120 in Ref. 70). The central fibrous body encasement for the AV node and distal AV bundle is clearly described by Tawara in the English translation by Suma and Shimada with a preface by Anderson (67). For the node, Tawara stated, “. . .the isolated muscle groups are now situated in the middle of the right half of the fibrous septum, between the ventricular and atrial septal musculature, surrounded by connective tissues of the atrioventricular septum” (see p. 10 in Ref. 67), and again, “. . .the node. . .is in the middle of the noncoronary aortic leaflet” (see p. 21 in Ref. 67). “For simplicity, I will call this portion of the atrial segment, i.e., the network, the ‘node.’” (see p. 139 in Ref. 67). For the distal AV bundle, “The anterior continuation of the node forms the ventricular segment of the connecting system. . . .I make the border at the place where the system penetrates the atrioventricular fibrous septum, because this place is easily determined anatomically.” (see p. 141 in Ref. 67). Nevertheless, the AV node and the distal AV bundle are termed the penetrating bundle by many investigators, e.g., Anderson and Ho (see Fig. 6 in Ref. 3) and Tohsida et al., (see Fig. 7 in Ref. 71) because of the central fibrous body encasement. This is surprising, because many of these same investigators depicted the AV node within the central fibrous body in their early studies, i.e., James in human (see Fig. 2 in Ref. 27) and dog (see Fig. 3 in Ref. 28), and Anderson et al. in rabbit hearts (see Fig. 4 in Ref. 4). The fibrous collar of Anderson et al. (4) is, in fact, the posterior orifice of the central fibrous body. Thus the AV node of many current-day conventional workers is not the AV node of Tawara depicted herein. AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE AJP-Heart Circ Physiol • VOL ping (12) and serial histological sections of the entire tricuspid orifice (61) and by the restriction of cholinesterase activity in annular myocardium of whole hearts to the AV junction region in studies by Bojsen-Moller and Tranum-Jensen (10). Are atrionodal bundles terminals of SA node extensions? Orthogonal sections corroborate that the atrionodal bundles are analogous to terminals of the SA node extensions, because the tail bifurcates and surrounds the coronary sinus and the head extension runs on a plane in line with the superior atrionodal bundle (reviewed in Ref. 54). The SA node extensions first described by Keith and Flack in 1907 (33), using fixatives that preserve the natural color of the tissues and fine dissections, were later demonstrated in whole rabbit heart stained for cholinesterase activity (10). It is of note that the SA node extensions should not be confused with the sinoatrial ring bundle fibers of the endocardium in the latter report. TranumJensen and Bojsen-Moller (72) showed by electron microscopy of thin sections made from the stained tissue that the SA node extensions are different from the adjacent myocardium in displaying small branching myocytes containing glycogen deposits and grouped in clusters. These attributes are also similar to those for the proximal AV bundle, AV node, and distal AV bundle (56, 58). The fact that the atrionodal bundles are associated with epicardium and terminate in the atrial-interventricular septal groove likely accounts for failure to detect connections of the extensions with the proximal AV bundle, the specialized AV ring tissues of Tranum-Jensen and BojsenMoller (72). References to the SA node extensions could not be found in studies by other investigators. Ordinary Myocardium of the AV Junction Region The anatomy herein demonstrates that in contrast to the specialized tissues of the AV junction region, the ordinary atrial myocardium is arrayed as sheets of myofibers with little to no collagen separating the myofibers or tissues (Fig. 7) and only ordinary atrial myocardium contact the annulus (Fig. 6, B and C). Significantly, the ordinary atrial myofibers are larger than myofibers of the specialized tissues (compare Figs. 5 and 7). The ordinary myofibers may appear to be encased in collagen when sandwiched by endocardial and epicardial connective tissue, and depending on the plane of section, yet the myofibers remain singly arrayed (Figs. 3L, 4N, and 6C4). Muscular valvular apparatus. Annular myocardium has long been known to be critical for valve function (19, 65) i.e., the sphincter-like contraction of the tricuspid orifice via the circumferential lamina that completely encircles the ring; and contraction of the valve leaflets via the perpendicular lamina that are offshoots of the circumferential myofibers and run radially to the ring and into only the major leaflets but are absent from the anterior medial leaflet. The perpendicular lamina augments competency of the leaflets in conjunction with the papillary muscle chordae tendoneae. Motion pictures have captured this action that occurred in the absence of a pressure gradient but in the presence of stretching with filling (65). Thus the perpendicular lamina myofibers are offshoots of the circumferential lamina and the latter merges with the pectinates, thereby forming the muscular valvular apparatus (54, 61). The circumferential lamina was shown to be the substrate for reentry in the Y-shaped canine model of atrial flutter, because it is the only myocardial continuum to encircle 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 account for failure to detect these tissues in histological sections of hearts fixed in the curved, natural state. Data herein are in keeping with the description of the topology, morphology, and cytoarchitecture for the proximal AV bundle as the atrial bundle component of the bridging fibers of Tawara (67, 69, 70) . The atrial bundle likewise has an elevated position under the fossa ovalis in its passage to the elevated valvula Thebessi and entry of the coronary sinus ostium (69). Tawara (70) described the atrial bundle (parallel fibers), and as part of the atrial segment, as translated by Suma and Shimada (67), “They do not make a complicated network, but run more parallel and posteriorly, usually not isolated but arranged in many small bundles. The fibers are separated from each other here by abundant connective tissue. . . .This portion of the atrial segment, consisting of muscle fibers running in parallel fashion, connects with the ordinary atrial muscle fibers.” (see p. 115 in Ref. 67). Thus Tawara did not detect the atrionodal bundles. The monograph contained sketches of histological sections that did not include the coronary sinus ostium nor the elevated entry edge, and drawings of the conduction system on photographs of whole hearts depicted only the Knoten and/or short parallel fibers of the proximal AV bundle at the PAVB-AVN junction. Tawara also did not demonstrate the full extent of the atrial (proximal AV) bundle and believed that the bundle, isolated by its connective tissue investment, was connected with ordinary atrial myocardium near the coronary sinus. It may be surprising that neither the atrionodal bundles nor the proximal AV bundle had been demonstrated in studies previously to those by Racker (49, 51). It is even more surprising that conventional studies still do not show structures compatible with the proximal AV bundle or the atrionodal bundles. This fact is clear by examination of the actual anatomic data that show in schematics that regions in which the atrionodal bundles and proximal AV bundle are found are either completely excluded by Hudson (see Fig. 2–11 in Ref. 25) and Smith et al. (see Fig. 2 in Ref. 64) or partially excluded by Lev et al. (see Fig. 1 in Ref. 35). When these regions are included in the schematics [e.g., Becker and Anderson (see Fig. 1 in Ref. 7), Inoue and Becker (see Fig. 1, in Ref. 26), Medkour et al. (see Fig. 1 in Ref. 42), Anderson and Ho (see Fig. 6 in Ref. 3), Waki et al. (see Figs. 3, 5, and 6 in Ref. 74), Zhang et al. (see Figs. 1 and 2 in Ref. 77), and Mazgalev et al. (see Fig. 7 in Ref. 37)] none of the features of the tissues appears in the published histological data. It is significant that the tendon of Todaro is shown well removed from the conduction tissues. Moreover, neither the specialized tissues nor the laminae are distinguishable in endocardial denuded hearts by SanchezQuintana et al. (see Fig. 2 in Ref. 63) and by Cabrera et al. (see Fig. 4 in Ref. 13). Thus structures compatible with the proximal AV bundle and atrionodal bundles are not seen in the above studies. Are there other atrionodal bundles? It appears unlikely that there are other specialized conduction tissues within the AV junction region, with the exception of remnants of bundles heading toward the left atrium in serial sections of the three orthogonal planes. Studies of the left atrium are thus needed to further access these remnants. Moreover, it appears unlikely that there are other specialized conduction tissues elsewhere within the tricuspid orifice, based on studies by Hoffman and colleagues using extracellular (20, 21) and intracellular map- H1867 H1868 AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE Physiological Significance for Transmission Correlated electrophysiological studies have shown that each of the specialized (including the SA node) and the ordinary atrial tissues evoke unique transmembrane and extracellular potentials and have unique transmission properties. Preferential transmission to the atrionodal bundles was shown as these tissues depolarize before other tissues in the AV junction region at the same site. Definitively, the specialized tissues maintain its original SA node rhythm with high extracellular potassium concentrations, whereas transmission ceases in the atrial myocardium across the atrium and at the SA node (49, 50, 53). The specialized tissues were thus deemed to be part of a sinoventricular conduction system. More recent observations corroborate this designation since early activation of the atrionodal bundles have been shown by transmembrane potential recordings (see Table in Ref. 62) and the potentials are significantly different from those of the ordinary atrial myocardium (see Table in Ref. 57). AJP-Heart Circ Physiol • VOL Overlapping myocardial circuits. Because cell-to-cell connections are required for electrotonic transmission, the anatomy herein shows that there are two overlapping independent atrial myocardial circuits in the AV junction region, i.e., the muscular valvular apparatus and the sinoventricular conduction system. Extracellular potentials from both the specialized and ordinary atrial myocardium appear in the same electrogram trace under normal conditions and result in the so-called low-high and high-low electrogram configurations seen clinically (for review see Ref. 54). Myocardial circuits of the AV junction region were identified as independent circuits using conditions that interrupted transmission in one circuit but not the other, such as high potassium, transections, pacing (49, 50, 53), and photolysis of the iontophoresed fluorescent probe Lucifer Yellow (unpublished observations). Termination of atrial flutter. Termination of long cycle length reentry pathways has been accomplished via small linear surgical (20, 21) or radiofrequency (43) lesions in the posterior atrial wall and or inferoposterior medial atrial wall. Importantly, lesions made in the inferoposterior medial atrial wall may affect the lateral atrionodal bundle (Fig. 6). Knowledge of electrical potentials evoked by atrionodal bundles will be helpful in avoiding these important tissues. Juxtaposition of the PAVB-AVN junction, the superior atrionodal bundle, and the narrow circumferential lamina region. Juxtaposition of the PAVB-AVN junction, the superior atrionodal bundle, and the narrow circumferential lamina region must be considered during manipulation of the AV junction region and likewise explains responses to certain procedures. These tissues occupy an area ⬍2 mm square (Fig. 6B). Thus catheter ablation of the so-called “fast pathway” for the cure of AV node reentry arrhythmias has the potential to 1) induce AV node block via interruption of the PAVB-AVN junction, which is not shielded by the tendon of Todaro or the central fibrous body, and 2) concomitantly terminate or preclude the occurrence of atrial flutter due to circus movement in the tricuspid ring, by dividing the circumferential lamina circus pathway at its narrowest point (Figs. 2 H–J, 3 E–F, and 6B). Thus recognition of the topography as well as discrete potentials evoked by the various tissues will be of great utility in avoiding untoward effects. Ordinary atrial high-resolution activation maps and late activation versus AV node activation. Anatomic data herein for the medial atrial wall are consistent with instantaneous activating wavefronts of depolarization for the atrial wall but not AV node activation as commonly believed. The maps were obtained from multiple unipolar electrodes in humans by McGuire et al. (39), in pig by deBakker et al. (15) and Hocini et al. (24), and in dog by Hocini et al. (24), with multiple micropipette electrodes by Billette (9), and with optical probes and voltage sensitive dyes by Choi and Salama (14) and Efimov and Mazgalev (18). Significantly, electrical potentials used to construct the maps have hallmark characteristics of ordinary atrial myocardium, i.e., spiked high-amplitude extracellular and triangular-shaped transmembrane potentials. Moreover, areas of late activation, which are taken as transitional zones to the AV node, [also see Antz et al. (5)] correlate instead with the atrial bridges and ordinary atrial wall myocardium. In corroboration with the anatomic view of later activation of the two atrial bridges, the bridges have been shown to be activated 20 to 30 ms, respectively, later than the superoan286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 the ring, and its division anywhere around the ring can terminate and preclude reentry. The atrial limb of the Y-shaped incision divides pectinates and enables reentry. Normally impulses from the pectinates activate the laminae but collide within the circumferential lamina, preventing sustained reentry (12, 20, 21, 54). Annular myocardium as the AV node and AV node inputs. Surprisingly, neither the circumferential nor the perpendicular laminae are delineated as discrete tissues nor is their function in valve closure considered by current day investigators. Instead, comparison of the actual anatomic data shows that annular myocardium and medial atrial wall myocardium have been and continue to be taken as inputs to the AV node and the AV node itself by Loh et al. (see Fig. 2 in Ref. 36) and by Nikolski et al. (see Fig. 4 in Ref. 44) as terminal twigs of internodal tracts by James (see Figs. 2B, 6, 7, and 9 in Ref. 27), as AV node transitional cells by Becker and Anderson (see Fig. 7 in Ref. 7), by Ho et al. (see Figs. 2, A–B, and 3A in Ref. 23), or as AV node approaches by McGuire et al. (see Figs. 11–12 in Ref. 40). The circumferential lamina myofibers are taken in part for the rightward extension of the AV node by Inoue and Becker (see Fig. 2B and Figs. 5–7 in Ref. 26) and the posterior nodal extensions by Medkour et al. (see Fig. 2 in Ref. 42) that supposedly are attenuated in the newborn but grow with age according to Waki et al. (see Figs. 5–6 in Ref. 74). The perpendicular lamina myofibers are taken as the transitional posterior zone of the AV node by Anderson et al. (see. Figs. 8–9A in Ref. 2), as transitional overlay fibers by Ho et al. (see Fig. 2 in Ref 23), and as anterior and posterior atrionodal connections by McGuire et al. (see Fig. 1 in Ref. 41). The entire medial atrial wall was interpreted as AV node inputs by Becker and Anderson in 1976 (see Fig. 14 in Ref. 7) and continued to be demonstrated as such by Sanchez-Quintana et al. in 1997 (see Fig. 2 in Ref. 63). Because the perpendicular and circumferential laminae are important for valve function and the circumferential lamina is implicated as a substrate for certain atrial arrhythmias, a more detailed knowledge of their electrical properties is needed. It will be important to compare genetic markers in these tissues. Distinguishing atrial from specialized tissues is simplified by the finding that only ordinary myocardium is singly arrayed as individual myofibers. AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE AJP-Heart Circ Physiol • VOL cell-to-cell connections, because negatively charged Lucifer Yellow dye diffusion was restricted to intracellular compartments of the dye-injected myocyte (49, 52). Moreover, after a total of 1 h of dye injection and 6 h diffusion time, the myocytes and fascicular pathway remained tightly coupled to the spontaneous SA node rate (see Figs. 1 and 4 and Table in Ref. 52). Fascicular transmission was seen by temporal differences in activation of myocytes within the same tissue. Temporal differences correlated with the appearance of asynchronous catheter and micropipette electrode potentials and under direct observation at the PAVB-AVN junction, AV node, and distal AV bundle (49, 50; see Fig. 5 in Ref. 53). Activation of a selected fascicle can be obtained by using coincident catheter, wire, and the micropipette electrode recording method (57, 62). Such recordings are important for accessing injury to a single myocyte and the response of its electrotonic pathway as well as nearby tissues. Clearly, the death of a single myocyte has no effect on transmission within a tissue, because a dead myocyte can be closed off from healthy myocytes by intercalated disks (49; see Fig. 6 in Ref. 52). On the other hand, disruption of a fascicular pathway can result in alterations in transmission to the targeted tissue. Disruption of a fascicular pathway was shown following photoablation of Lucifer Yellow containing fascicles by death of the myocyte (indicated by loss of the wire potential), anisotropy in transmission in the tissue (indicated by bizarre catheter potential in the wire electrode recording), and alterations in transmission downstream (indicated by skipped AV nodal beats; unpublished findings). Significantly, the photoablation recordings also corroborate that the atrial myocytes do not share connections with the specialized tissues, because side-by-side extracellular potentials appeared unchanged in the same traces showing cell death of the atrionodal bundle fascicles. Fascicular transmission by multiple elements of the same tissue is taken to provide redundancy of signal and the three atrionodal bundles to function in optimizing and regulating efficiency of ventricular pumping under various conditions of normal activity, sleep, or exercise. Studies are needed to determine how the atrionodal bundles effect AV node transmission in the beat-to-beat functioning of the heart, whether specific regions of the ventricles are targeted by a specific atrionodal bundle input and how these tissues and the laminae of the annular myocardium might participate in cardiac arrhythmias. Transmission at the cellular level. The anatomy herein demonstrates that current concepts for physiology of myocardial transmission will have to be revised as a hallmark of transmission links increasing speed of propagation with the increasing size of a fiber. However, the specialized myofibers are, in fact, an order of magnitude smaller than the working ordinary myofibers (Figs. 5 and 7), yet transmission is faster in the smaller atrionodal bundle myofiber as explicitly shown by the low-high double potentials of extracellular electrograms, as noted earlier. In conclusion, although facts of the anatomy cannot be changed, our ability to appreciate the anatomy is determined by the protocols that are used to preserve it. With respect to nomenclature, new structures (e.g., coronary sinus fossa) were so named for the sake of anatomic utility. However, the term atrionodal bundle nevertheless remains unwieldy, because 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 terior medial atrial wall (see Table in Ref. 62). It is significant that the specialized tissues all possess low-amplitude extracellular potentials and transmembrane potentials with plateaus (atrionodal bundles), domes (proximal AV bundle), or phase 4 depolarizing upstrokes (AV node). The atrionodal bundles are activated before the atrial myocardium at each site, as explicitly shown by the low amplitude waveform of the so-called low-high double potentials (49, 50, see SAB trace in Fig. 6 in Ref. 53 and Ref. 54 for review). Whereas, slow transmission occurs in the proximal AV bundle, which is implicit in the low-amplitude waveform of long duration in the so-called high-low double potentials (49, 50, and see Figs. 5–9 in Ref. 53), activation of the proximal AV bundle and punctate junctions with the atrionodal bundles occurs ⬎100 ms later than in the atrionodal bundles and the ordinary atrial myocardium. Thus it seems more likely that the atrial maps show activation of the ordinary atrial myocardium only, as is the case for the high-resolution ventricular maps and not the AV node as intended. Sinoventricular conduction system high-resolution activation maps. Topographic markers for the SA node, atrionodal bundles, proximal AV bundle, AV node, and distal AV bundle, and delineation of the unique low-amplitude potentials of the specialized tissues permitted simultaneous mapping of beat-tobeat activity in the sinoventricular conduction system tissues by Racker and colleagues (49, 50, 53, 57, 62). The sinoventricular conduction system maps show that each of the tissues transmit impulses differently with transmission being very rapid in the atrionodal bundles, occurring tens of milliseconds before the ordinary atrial myocardium; impulses are slowest in the proximal AV bundle but speed up in the AV node and are rapid again in the distal AV bundle. Evidence for slowing of transmission at punctate junctions between tissues has been demonstrated (see Fig. 10 in Ref. 53). Pronounced coiling of the proximal AV bundle intrafascicular myofibers appears to be the most probable anatomic basis for transmission being slowest in this tissue (see Figs. 4 and 5 in Ref. 58), whereas rapid transmission in the atrionodal bundles and the distal AV bundle correlates with the parallel ordering of their myofibers and fascicles and with the rapid upstroke of their transmembrane potentials. Finger-like, serpentine myocyte extensions of the AV node (see Fig. 7 in Ref. 58) and proximal AV bundle (see Fig. 3D in Ref. 58) correlate with the slow upstroke of AV node transmembrane potentials during orthograde transmission and the slow upstroke of proximal AV bundle transmembrane potentials during retrograde transmission. The presence of unusual fascicles of nerve endings in the proximal AV bundle and AV node shows autonomic control of transmission in these tissues (see Figs. 3 B–D and 6, B and D, inset in Ref. 58). The collagen encasement of fascicles comprising the tissues most probably function as insulation, and that of the central fibrous location of the AV node and distal AV bundle as protection from compression, as adult-like ECG patterns are recorded in the bent single primordial heart tube before the appearance of collagen or the central fibrous body by Patten (47). Transmission at the fascicular level. Organization of myofibers into multiple fascicles, as shown herein for the atrionodal bundles, was shown to have a surprising consequence as the basic mechanism of transmission by the specialized tissues. Specifically, fascicles within a tissues were shown to share no H1869 H1870 AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE these bundles, which come from the atrium, join the proximal AV bundle and not the AV node. To term the His bundle as distal AV bundle is likewise unfortunate and may appear to be disrespectful. Finally, it appears irrational to consider that impulse transmission pathways exist between tissues that share no gap junction channels. In the AV junction region, the anatomy shows that the specialized and ordinary tissues, although overlapping, are independent circuits. ACKNOWLEDGMENTS The author thanks Dr. Brian F. Hoffman for leading the way and Dr. Alan H. Kadish for constant encouragement and support. The photographic assistance of Vanessa Jones and Gail Alfred is gratefully acknowledged. The author is indebted to Dr. Hsueh-Hwa Wang for carefully reading this manuscript and for helpful discussion and suggestions. This work was supported by the Northwestern Memorial Foundation, Jadwiga Roguska-Kytes, Grant NMH 572; National Heart, Lung, and Blood Institute Grants HL-40667 and HL-30557 (to Dr. Brian F. Hoffman); American Heart Association Grant 9951464Z; Dr. Scholl Foundation Grant 990841. REFERENCES 1. Anderson RH and Becker AE. Cardiac Anatomy: an Integrated Text and Color Atlas. London: Gower Medical, 1980. 2. Anderson RH, Becker AE, Brechenmacher C, Davies MJ, and Rossi L. The human atrioventricular junctional area: a morphological study of the AV node and bundle. Euro J Cardiol 3: 11–25, 1975. 3. Anderson RH and Ho SY. Anatomic criteria for identifying the components of the axis responsible for atrioventricular conduction. J Cardiovasc Electrophysiol 12: 1265–1268, 2001. 4. Anderson RH, Janse MJ, Van Capelle FJ, Billette J, and Becker AE. A combined morphological and electrophysiological study of the AV node in rabbit heart. Circ Res 35: 909–922, 1974. 5. Antz M, Scherlag BJ, Otomo K, Pitha J, Tondo C, Patterson E, Jackman WM, and Lazzarra R. Evidence for multiple atrio-AV nodal inputs in the normal dog heart. J Cardiovasc Electrophysiol 9: 395–408, 1998. 6. Baird J and Robb J. Study, reconstruction, and gross dissection of the atrioventricular conducting system of the dog heart. Anat Rec 108: 747– 762, 1950. 7. Becker AE and Anderson RH. Morphology of the human atrioventricular junctional area. In: Conduction System of the Heart, edited by Wellens HJ, Lee KI, and Janse MJ. Philadelphia: Lea and Febiger, 1976, p. 263–286. 8. Bharati S. Anatomic boundaries between the atrioventricular node and the atrioventricular bundle. J Cardiovasc Electrophysiol 9: 228, 1998. 9. Billette J. Atrioventricular nodal activation during premature stimulation of the atrium. Am J Physiol Heart Circ Physiol 252: H163–H177, 1987. 10. Bojsen-Moller F and Tranum-Jensen J. Rabbit heart nodal tissue, sinuatrial ring bundle and atrioventricular connections identified as a neuromuscular system. J Anat 112: 367–382, 1972. 11. Boyden P. Models of atrial reentry. J Cardiovasc Electrophysiol 6: 313–324, 1995. 12. Boyden PA, Frame LH, and Hoffman BF. Activation mapping of reentry an anatomic barrier in the canine atrium: observations during entrainment and termination. Circulation 79: 406–416, 1989. 13. Cabrera JA, Sanchez-Quintana D, Ho SY, Medina A, and Anderson RH. The architecture of the atrial musculature between the orifice of the inferior caval vein and the tricuspid valve: The anatomy of the isthmus. J Cardiovasc Electrophysiol 9: 1186–1195, 1998. 14. Choi BR and Salama G. Optical mapping of atrioventricular node reveals a conduction barrier between atrial and nodal cells. Am J Physiol Heart Circ Physiol 274: H829–H845, 1998. 15. De Bakker J, Coronel R, McGuire MA, Vermeulen JT, Opthof T, Tasseron S, Van Hemel NM, and Defauw JJ. Slow potentials in the atrioventricular junctional area of patients operated on for atrioventricular node tachycardias and in isolated porcine hearts. J Am Coll Cardiol 23: 709–715, 1994. 16. De Mello WC and Hoffman BF. Potassium ions and electrical activity of specialized cardiac fibers. Am J Physiol 199: 1125–1130, 1960. AJP-Heart Circ Physiol • VOL 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 GRANTS 17. Dinari G and Aygen M. Sinoventricular conduction. New Eng J Med 289: 1238–1239, 1973. 18. Efimov IR and Mazgalev TN. High-resolution, three-dimensional fluorescent imaging reveals multilayer conduction pattern in the atrioventricular node. Circulation 98: 54–57, 1998. 19. Ellison JP and Hibbs RG. The atrioventricular valves of the guinea-pig: a light microscopy study. Am J Anat 138: 331–346, 1973. 20. Frame LH, Page RL, Boyden PA, Fenoglio JJ, and Hoffman BF. Circus movement in the canine atrium around the tricuspid ring during experimental atrial flutter and during reentry in vitro. Circulation 76: 1155–1175, 1987. 21. Frame LH, Page RL, and Hoffman BF. Circus movement in the canine atrium around the tricuspid ring during experimental atrial flutter and during reentry in vitro. Circ Res 58: 495–511, 1986. 22. Hirao K, Scherlag BJ, Poty H, Otomo K, Tondo C, Antz M, Patterson E, Jackman WM, and Lazzara R. Electrophysiology of the atrio-AV nodal inputs and exits in the normal dog heart: radiofrequency ablation using an epicardial approach. J Cardiovasc Electrophysiol 8: 904–915, 1997. 23. Ho SY, Kilpatrick L, Kanai T, Germroth PG, Thompson RP, and Anderson RH. The architecture of the atrioventricular conduction axis in dog compared to man: its significance to ablation of the atrioventricular nodal approaches. J Cardiovasc Electrophysiol 6: 26–39, 1995. 24. Hocini M, Loh P, Ho SY, Sanchez-Quintana D, Thibault B, deBakker JMT, and Janse MJ. Anisotropic conduction in the triangle of Koch of mammalian hearts: electrophysiologic and anatomic correlations. J Am Coll Cardiol 31: 629–636, 1998. 25. Hudson RE. The conduction system. In: Cardiovascular Pathology. Baltimore, MD: Williams & Wilkins, 1965, vol. 1, p. 53–145. 26. Inoue S and Becker AE. Posterior extensions of the human compact atrioventricular node. Circulation 97: 188–193, 1998. 27. James TN. Morphology of the human atrioventricular node with remarks pertinent to its electrophysiology. Am Heart J 62: 756–771, 1961. 28. James TN. Anatomy of the A-V node of the dog. Anat Rec 148: 15–27, 1964. 29. James TN. Anatomy of the crista supraventricularis: its importance for understanding right ventricular function, right ventricular infarction, and related conditions. J Am Coll Cardiol 6: 1083–1095, 1985. 30. James TN and Sherf L. Fine structure of the His bundle. Circulation 64: 9–28, 1971. 31. James TN, St. Martin E, Willis PW, and Lohr TO. Apoptosis as a possible cause of gradual development of complete heart block and fatal arrhythmias associated with absence of the AV node, sinus node, and internodal pathways. Circulation 93: 1424–1438, 1996. 32. Janse MJ and Anderson RH. Specialized internodal atrial pathways-fact or fiction? Eur J Cardiol 2: 117–136, 1974. 33. Keith A and Flack M. The form and nature of the muscular connections between the primary divisions of the vertebrate heart. J Anat Physiol 41: 172–189, 1907. 34. Lev M and Bharati S. The anatomy of the conduction system. In: Physiology of Atrial Pacemakers and Conductile Tissues. New York: Futura, 1980, p. 21–36. 35. Lev M, Widran J, and Erickson EE. A method for the histopathologic study of the atrioventricular node, bundle and branches. Arch Path 52: 73–83, 1951. 36. Loh P, Ho SY, Kawara T, Hauer RNW, Janse MJ, Breithardt G, and deBakker JMT. Reentrant circuits in the canine atrioventricular node during atrial and ventricular echoes electrophysiological and histological correlations. Circulation 108: 231–238, 2003. 37. Mazgalev TN, Ho SY, and Anderson RH. Anatomic-electrophysiological correlations concerning the pathways for atrioventricular conduction. Circulation 103: 2660–2667, 2001. 38. McAlpine WA. Heart and Coronary Arteries. New York: SpringerVerlag, 1975. 39. McGuire Bourke JP MA, Robotin MC, Johnson DC, Meldrum-Hanna W, Nunn GR, Uther J, and Ross DL. High resolution mapping of Koch’s Triangle using sixty electrodes in humans with atrioventricular junctional (AV nodal) reentrant tachycardia. Circulation 88: 2315–2328, 1993. 40. McGuire MA, deBakker JMT, Vermeulen JT, Opthof T, Becker AE, and Janse MJ. Origin and significance of double potentials near the atrioventricular node correlation of extracellular potentials, intracellular potentials, and histology. Circulation 89: 2351–2360, 1994. 41. McGuire MA, Robotin M, Yip ASB, Bourke JP, Johnson DC, Dewsnap BI, Grant P, Uther JB, and Ross DL. Electrophysiologic and AV JUNCTION REGION MORPHOLOGY AND CYTOARCHITECTURE 42. 43. 44. 45. 46. 47. 49. 50. 51. 52. 53. 54. 55. 56. 57. 58. 59. AJP-Heart Circ Physiol • VOL 60. Racker DK and Tse WW. A correlated gross anatomical and histological study of the canine atrioventricular (AV) junctional region (Abstract). Anat Rec 211: 157A, 1985. 61. Racker DK, Ursell PC, and Hoffman BF. Anatomy of the tricuspid annulus circumferential myofibers as the structural basis for atrial flutter in a canine model. Circulation 84: 841–851, 1991. 62. Racker DK, Wahler GM, and Kadish AH. Atrionodal bundles have unique intracellular and extracellular properties confirming the presence of a specialized conduction system in the right atrium. PACE 21: 920, 1998. 63. Sanchez-Quintana D, Davies W, Ho SY, Oslizlok PO, and Anderson RH. Architecture of the atrial musculature in and around the triangle of Koch: its potential relevance to atrioventricular nodal reentry. J Cardiovasc Electrophysiol 8: 1396–1407, 1997. 64. Smith A, Ho SY, and Anderson RH. Histological study of the cardiac conducting system as a routine procedure. Med Lab Sci 34: 223–229, 1977. 65. Smith HI, Essex HE, and Badles EJ. A study of the movements of heart valves and of heart sounds. Ann Int Med 33: 1357–1359, 1950. 66. Sonnenblick EH, Napolitano LM, Daggett WM, and Cooper T. An intrinsic neuromuscular basis for mitral valve motion in the dog. Circ Res 21: 9–15, 1967. 67. Suma K and Shimada M. The Conduction System of the Mammalian Heart. London: Imperial College Press, 2000. 68. Sweeney LJ and Rosenquist GC. The normal anatomy of the atrial septum in the human heart. Am Heart J 98: 194–199, 1979. 69. Tawara S. Die topographie und histologie der bruckenfasern. Ein beitrag zur lehre von der bedeutung der Purkinjeschen faden. Zentralbl f Physiol 19: 70–76, 1906. 70. Tawara S. DAS Reizletungssystem Des Saugetierherzens. Ein anatomisch-histologische studie uber das atriovertrikularbundle und die Purkinjeschen faden. Jena: Verlag Von Gustav Fischer, 1906. 71. Toshida N, Hirao K, Yamamoto N, Tanaka M, Suzuki F, and Isobe M. Ventricular echo beats and retrograde atrioventricular nodal exits in the dog heart: multiplicity in their electrophysiology and anatomic characteristics. J Cardiovasc Electrophysiol 12: 1256–1264, 2001. 72. Tranum-Jensen J and Bojsen-Moller F. The ultrastructure of the sinuatrial ring bundle and of the caudal extension of the sinus node in the right atrium of the rabbit heart. Zellforsch 138: 97–111, 1973. 73. Vassalle M and Hoffman BF. The spread of sinus activation during potassium administration. Circ Res 17: 285–295, 1965. 74. Waki K, Kim JS, and Becker AE. Morphology of the human atrioventricular node is age dependent: a feature of potential clinical significance. J Cardiovasc Electrophysiol 11: 1144–1151, 2000. 75. Walmsley R and Watson H. The medial atrial wall of the right atrium. Circulation 34: 400–411, 1966. 76. Yuill KH and Hancox JC. Characteristics of single cells isolated from the atrioventricular node of the adult guinea-pig heart. Eur J Physiol 445: 311–320, 2002. 77. Zhang Y, Bharati S, Mowrey KA, Zhuang S, Tchou PJ, and Mazgalev TN. His electrogram alternans reveal dual-wavefront inputs into and longitudinal dissociation within the bundle of His. Circulation 104: 832–838, 2001. 78. Zimmerman J and Bailey CP. The surgical significance of the fibrous skeleton of the heart. J Thor Cardiovasc Surg 44: 701–712, 1962. 286 • MAY 2004 • www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.32.246 on May 8, 2017 48. histologic effects of dissection of the connections between the atrium and posterior part of the atrioventricular node. J Am Coll Cardiol 23: 693–701, 1994. Medkour D, Becker AE, Khalife K, and Billette J. Anatomic and functional characteristics of a slow posterior AV nodal pathway role in dual-pathway physiology and reentry. Circulation 98: 164–174, 1998. Moulton K, Miller B, Scott J, and Woods WT. Radiofrequency catheter ablation for AV nodal reentry: a technique for rapid transition of the slow AV nodal pathway. PACE 16: 760–768, 1993. Nikolski VP, Jones SA, Lancaster MK, Boyett MR, and Efimov IR CX43 and dual-pathway electrophysiology of the atrioventricular node and atrioventricular nodal reentry. Circ Res 92: 469–475, 2003. Olgin JE, Ursell PC, Kao AK, and Lesh MD. Pathological findings following slow pathway ablation for AV nodal reentrant tachycardia. J Cardiovasc Electrophysiol 7: 625–631, 1996. Papez JW. Heart musculature of the atria. Am J Anat 27: 255–285, 1920. Patten BM. The development of the sinoventricular conduction system. Univ Mich Med Bull 22: 1–21, 1956. Racker DK. Transmission Electron Microscopy: Methods of Application. Springfield, IL: CC Thomas, 1983. Racker DK. Gross Anatomical, Histological and Electrophysiological Definitions of AV Nodal Inputs in the Canine Heart (PhD thesis). Chicago, IL: University of Health Sciences/The Chicago Medical School, 1987. Racker DK. The effects of 10 mM K⫹ on electrical activity within the superior atrionodal (SAB) and proximal AV bundles (PAVB) in the canine heart (Abstract). FASEB J 2: 3696, 1988. Racker DK. The atrioventricular node and input pathways: a correlated gross anatomical and histological study of the canine AV junctional region. Anat Rec 224: 336–354, 1989. Racker DK. The demonstration of intercellular coupling between cells of the canine distal AV bundle using lucifer yellow-CH and histological sections. J Mol Cell Cardiol 21: 479–493, 1989. Racker DK. Sinoventricular transmission in 10 mM K⫹ by canine atrioventricular nodal inputs: superior atrionodal bundle and proximal atrioventricular bundle. Circulation 83: 1738–1753, 1991. Racker DK. Transmission and reentrant activity in the sinoventricular conducting system and in the circumferential lamina of the tricuspid valve. J Cardiovasc Electrophysiol 4: 513–525, 1993. Racker DK. The AV junction region of the heart: a comprehensive study correlating gross anatomy and direct three-dimensional analysis. Part I. Architecture and topography. Anat Rec 256: 49–63, 1999. Racker DK and Kadish AH. Morphology of the proximal AV bundle, the AV node and the distal AV bundle by three dimensional analysis. PACE 20: 1116, 1997. Racker DK and Kadish AH. The canine atrionodal bundle multilimb input to the AV node possess unique intracellular and extracellular potentials (Abstract). FASEB J 13: A105, 1999. Racker DK and Kadish AH. Proximal atrioventricular bundle, atrioventricular node and the distal atrioventricular bundle are distinct anatomic structures with unique histologic characteristics and innervation. Circulation 101: 1049–1059, 2000. Racker DK, Kadish AH, and Hoffman BF. Atrioventricular node input pathways by direct three dimensional analysis (Abstract). PACE 19: 732, 1996. H1871