Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
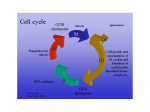
APC/C Cyc B CDK1 Cyc B WEE1/MIT1 APC/C WEE1/MIT1 M CDK1 Cyc D1 Cyc A CDK4 p16INK4A, INK4 proteins CDK2 G2 G1 Cyc D1 CDK4 Cyc A WEE1/MIT1 p16INK4A, INK4 proteins CDK2 S Cyc A CDK2 Cyc E Cyc E CDK2 Cyc D1 SCFFbx4 CDK4 p21CIP1 p27KIP1 p57KIP2 WEE1/MIT1 CDK2 p21CIP1 p27KIP1 p57KIP2 WEE1/MIT1 SCFFbw7 Figure 1.1. Temporal CDK activity controls cell cycle progression. Mitogen-dependent expression of D-type cyclins facilitates activation of G1-phase CDK4/6. The mammalian family of INK4 proteins specifically inhibit CDK4/6 activity by direct binding to the CDK, antagonizing CDK activity. Nuclear cyclin D/CDK4/6 kinase activity drives cell cycle progression beyond the restriction point, thereby committing the cell to one round of division. Following the G1/S transition, nuclear CDK4/6 activity is terminated by ubiquitin mediated proteolysis of cyclin D1 by the SCFFbx4 E3 ubiquitin ligase. Cyclin E expression in late G1 facilitates CDK2 activation, and association with the CIP/KIP family of CDK inhibitors inhibits its kinase activity. CDC25-dependent removal of inhibitory threonine/tyrosine phosphorylation on CDK2 is also essential for activation. Cyclin E/CDK2 activity drives E2F-dependent gene transcription, replication origin firing and S-phase progression, and this activity is attenuated in early-S phase by SCFFbw7-mediated ubiquitylation and degradation of cyclin E. Cyclin A expression also activates CDK2 in S-phase, facilitating DNA replication and inhibiting origin re-licensing. Cyclin A/CDK2 activity persists through G2 phase, functioning in centrosome duplication. Cyclin B is the sole mitotic cyclin, as cyclin B/CDK1 activity is essential for both centrosome duplication and mitotic processes. Both cyclin A and cyclin B are substrates of the G2/M phase E3 ligase, the anaphase promoting complex/cyclosome (APC/C). 24 B. A. G1 G1/S CycD1 CycD1 CDK4 Cyc E CDK4 CDK2 PPP Rb CycD1 Late G1 p21 CDK4 p27 GSK3 S12 P P Fbx4 dimerization Ligase activation E2F E2F Target Genes E2F DP Nucleus Cytoplasm D. C. G1/S S Cyc E GSK3 CDK2 T286 P Cdc45 Cdc6 Cdt1 ORC MCM 2-7 MCM Licensed Replication Origin 2-7 CycD1 CDK4 Ub K48-Ub Nucleus Cytoplasm 26S Proteasome Figure 1.2. Cell cycle-dependent cyclin D1 regulation. (A) Mitogen-dependent cyclin D1 expression is required for CDK4/6 activation during G1 phase. Active, nuclear cyclin D1/CDK4 kinase promotes cell cycle progression in two ways. First, cyclin D1/CDK4 catalyzes Rb phosphorylation, thereby triggering release of E2F transcription factors. Second, cyclin D1/CDK4 complexes titrate the CKIs p21CIP1 and p27KIP1 away from cyclin E/CDK2, facilitating CDK2 activation, full Rb inactivation, and gene transcription required for S-phase entry. (B) In late G1 phase, AKT-dependent inhibitory phosphorylation of GSK3β is alleviated, allowing GSK3β kinase activation. In the cytoplasm, GSK3β phosphorylates Fbx4 on serine 12, creating a consensus 14-33ε docking site. 14-3-3ε binding promotes Fbx4 dimerization and ligase activation. (C) Cyclin E/CDK2 activity facilitates entry into S-phase and DNA replication. Following the G1/S transition, nuclear cyclin D1/CDK4 kinase activity is no longer required. At this time, GSK3β enters the nucleus and phosphorylates cyclin D1 on T286, which triggers CRM1-mediated nuclear export. (D) Once in the cytoplasm, phosphorylated cyclin D1 is recognized by the SCFFbx4-αB crystallin E3 ubiquitin ligase. Cyclin D1 is polyubiquitylated and targeted for degradation by the 26S proteasome. 25 A. SCF E3 Ligases Ub Substrate B. SCF-like E3 Ligases Ub Ub Substrate C. S-phase CDT1 ORC PCNA Ub P MCM 2-7 MCM 2Replication 7 Origin Ub 26S Proteasome Figure 1.3. SCF and SCF-like E3 ubiquitin ligases. (A) Prototypical SKP-CULLIN-F-BOX (SCF) complex composition. The core cullin scaffold (CUL1) associates with the RING-finger protein RBX1 which is required for E2 recruitment and transfer of charged ubiquitin to the substrate. SKP1 also associates directly with CUL1 and facilitates substrate recruitment by binding the F-box domain within an F-box protein, an adapter for substrate recognition. Interaction of substrates with the SCF permits transfer of ubiquitin onto the substrate molecule by the E2 enzyme. (B) SCF-like E3 ligases contain RBX1, but harbor different cullin scaffold proteins and substrate adapters. The CUL7-based E3 ligase binds SKP1 and the FBW8 F-box protein, specifically, thereby functioning much like the canonical SCF. The CUL4-based ligase utilizes a different set of substrate adapters, including DDB1 and a DCAF protein that coordinates substrate recognition. (C) The CUL4DDB1/CDT2 E3 ligase maintains DNA replication fidelity. Following origin firing in S-phase, cyclin A/CDK2dependent phosphorylation of pre-RC components prevents origin re-licensing. Importantly, chromatin-bound CDT1 is recognized by the CDT2 WD-repeat DCAF adapter, which is targeted to chromatin through its PCNA interacting PIP box domain. CDT1 is subsequently polyubiquitylated by the CUL4 E3 ubiquitin ligase and targeted for proteasomal degradation. 26 Endometrial Cancer Esophageal, Endometrial Cancer Breast Cancer GSK3β P T286 CycD1 Ub Nuclear Export CDK4 Nucleus Cytoplasm 26S Proteasome Esophageal Cancer 14-3-3ε S12 P GSK3β P Dimerization Active Ligase Figure 1.4. Cyclin D1 regulatory pathways are targeted in human cancer. Cyclin D1 protein accumulation is tightly controlled via phosphorylation-dependent proteolysis, and mutations targeting cyclin D1 phosphorylation or degradation contribute to neoplastic transformation. Specific disruption of T286 phosphorylation occurs in endometrial cancer, and mutations targeting Fbx4 have been indentified in esophageal cancer. Furthermore, αB crystallin loss occurs in tumor-derived breast carcinoma cell lines. Ultimately, impaired cyclin D1 proteolysis promotes accumulation of active cyclin D1/CDK4 kinase, triggering DNA re-replication and subsequent genomic instability necessary for neoplastic transformation. 27 Protein Targeted Mutation Consequence Tumor Type Cyclin D1 T286R Constitutively Nuclear Esophageal Cyclin D1 Δ266-295 Constitutively Nuclear Cyclin D1 P287A Constitutively Nuclear Esophageal Tumor-derived esophageal carcinoma cell lines TE3, TE7, and TE12 Cyclin D1 P287S/T Constitutively Nuclear Endometrial Cyclin D1 Δ289-292 Constitutively Nuclear Endometrial αB crystallin Δ chr. 11 Tumor-derived breast cancer cell lines (MCF-7, MDA-MB 231) Fbx4 S8R Impaired ligase activity Esophageal Fbx4 S12L Disrupts phosphorylation “ Fbx4 P13S Disrupts phosphorylation “ Fbx4 L23Q Dimerization-deficient “ Fbx4 G30N “ Fbx4 P76T “ Table 1. Summary of mutations targeting cyclin D1 phosphorylation or Ub-ligase function. Mutations disrupting GSK3β-dependent cyclin D1 T286 phosphorylation and nuclear export include mutation of T286, P287, and deletion of residues corresponding to the CRM1 binding site. Mutations targeting the SCFFbx4-αB crystallin E3 ubiquitin ligase result in impaired ligase activity toward cyclin D1 and subsequent cyclin D1/CDK4 accumulation in the nucleus. 28 A. IR Replication Fork Collapse Intra-S Checkpoint P ATM CHK2 CDC25 p53 p21 H2AX CDK 2, CDK1 Signal Amplification, DNA Repair DSB UV Light Nucleotide Depletion Replication Stress Rad9/Rad1/Hus1 Rad17 DNA Replication Checkpoint P ATR ATRIP CHK1 CDC25 p53 p21 CDK 2, CDK1 Stalled DNA Replication B. DNA damage PhosphorylationIndependent G1 PhosphorylationDependent ATM G1 P APC/C CycD1 CDK4 Ub-mediated proteolysis (APC/C) G1 arrest PhosphorylationDependent CycD1 S-phase P ERK GSK3 P CycD1 P GSK3 CDK4 CDK4 Ub-mediated proteolysis (SCF) G1 arrest Ub-mediated proteolysis (SCF) Accelerated Proteolysis Intra S-Checkpoint Figure 1.5. DNA damage checkpoint responses prevent genomic instability. (A) S-phase DNA damage checkpoint activation. The intra-S-phase checkpoint response is mediated by ATM activation following DSB induction in genomic regions outside of active DNA replication. Genotoxic insults such as IR trigger DSBs and subsequent ATM activation. DSBs also result when stalled replication forks collapse, leading to ATM activation. ATM auto-phosphorylation catalyzes its monomerization from an inactive dimeric conformation, thereby facilitating downstream effector phosphorylation events necessary for cell cycle arrest and DNA repair. Replication stress, on the other hand, triggers ATR activation following accumulation of single-strand DNA (ssDNA). ssDNA is rapidly coated with replication protein A (RPA), which promotes recruitment of the ATR kinase and its cofactor ATRIP. Molecular adapters TopBP1 and claspin facilitate ATR-dependent Chk1 activation. Additional ATR substrates are required for cell cycle arrest and DNA repair, as in the ATM-dependent DSB response. (B) Cyclin D1 regulation following DNA damage. Previous reports established regulatory mechanisms for cyclin D1 destruction following G1-phase DSB induction. Left panel: proposed phosphorylation-independent cyclin D1 destruction by APC/C. Middle panel: proposed ERK-mediated cyclin D1 phosphorylation and subsequent recognition by the SCFFbxo31 ubiquitin ligase. Right panel: proposed mechanism for SCFFbx4-mediated cyclin D1 destruction following S-phase DNA damage. This model is tested and confirmed in chapter 2 of this thesis. 29