Survey
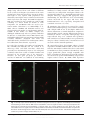
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
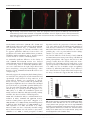
Journal of General Virology (2007), 88, 2899–2904 Short Communication DOI 10.1099/vir.0.82907-0 Enteroglial and neuronal involvement without apparent neuron loss in ileal enteric nervous system plexuses from scrapie-affected sheep Giuseppe Marruchella,1 Ciriaco Ligios,2 Valeria Albanese,1 Maria Giovanna Cancedda,2 Laura Madau,2 Giovanna Lalatta-Costerbosa,3 Maurizio Mazzoni,3 Paolo Clavenzani,3 Roberto Chiocchetti,3 Giuseppe Sarli,4 Luigi De Grossi,5 Umberto Agrimi,6 Adriano Aguzzi7 and Giovanni Di Guardo1 Correspondence Giovanni Di Guardo [email protected] 1 Department of Comparative Biomedical Sciences, Faculty of Veterinary Medicine, University of Teramo, Teramo, Italy 2 Istituto Zooprofilattico Sperimentale della Sardegna, Sassari, Italy 3 Department of Veterinary Morphophysiology and Animal Productions, Faculty of Veterinary Medicine, University of Bologna, Ozzano Emilia (Bologna), Italy 4 Department of Veterinary Public Health and Animal Pathology, Division of Veterinary Pathology, Faculty of Veterinary Medicine, University of Bologna, Ozzano Emilia (Bologna), Italy 5 Istituto Zooprofilattico Sperimentale delle Regioni Lazio e Toscana, Viterbo, Italy 6 Istituto Superiore di Sanità, Department of Food Safety and Veterinary Public Health, Rome, Italy 7 Institute of Neuropathology, University Hospital of Zurich, Zurich, Switzerland Received 6 February 2007 Accepted 29 June 2007 The enteric nervous system (ENS) probably plays a dominant role in sheep scrapie pathogenesis, but little is known about the cell types involved. We investigated the ileal myenteric and submucosal plexuses of four naturally and four orally experimentally scrapie-affected ARQ/ARQ Sarda sheep, as well as those of 12 healthy-control Sarda sheep carrying different PrP genotypes. All scrapie-affected animals, euthanized at clinical-disease end stage, showed PrPd deposition within enteric glial cells (EGCs) and calbindin-immunoreactive (CALB-IR) and neuronal nitric oxide synthase (nNOS)-IR neurons. Whole-mount investigations revealed no significant differences between the densities of total, CALB-IR and nNOS-IR neurons in scrapieaffected versus healthy sheep, irrespective of PrP genotype. Our results suggest that EGCs and CALB-IR and nNOS-IR neurons are probably involved in the pathogenesis of natural and oral experimental sheep scrapie. Furthermore, the infectious agent may be less pathogenic towards ENS neurons than it is towards central nervous system neurons. Sheep scrapie is the ‘prototype’ of transmissible spongiform encephalopathies (TSEs) or prion diseases, a group of neurodegenerative disorders affecting humans and animals. The key pathogenetic event in TSEs is the accumulation within the central nervous system (CNS) and peripheral tissues of an abnormal isoform (disease-specific PrP, PrPd) of the host-encoded cellular prion protein, PrPC (Aguzzi & Polymenidou, 2004). throughout the PrP gene (PRNP) (Prusiner, 1998). In sheep, PRNP codons 136, 154 and 171 are of greatest importance in modulating susceptibility/resistance to scrapie (Goldmann et al., 1994). In Suffolk and Sarda breeds (Westaway et al., 1994; Vaccari et al., 2001), codon 171 plays a major role in disease-susceptibility control, whereas in Cheviot sheep, the additional influence of codon 136 has been reported (Goldmann et al., 1994). Genotype of the host is known to modulate susceptibility/ resistance to TSEs, which is dependent on polymorphisms The gastrointestinal tract is probably the natural prion entry site, with the enteric nervous system (ENS) playing, along with palatine tonsils and Peyer’s patches (PPs), a crucial role in the early pathogenesis of animal and human TSEs (Mabbott & MacPherson, 2006). It has been suggested that ENS plexuses could act as the initial site A supplementary table showing tissue antigens and details of the monoclonal and polyclonal antibodies utilized in the study is available with the online version of this paper. 0008-2907 G 2007 SGM Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sun, 07 May 2017 13:45:51 Printed in Great Britain 2899 G. Marruchella and others of neuroinvasion for prions, which may subsequently gain access to the CNS through sympathetic and parasympathetic efferent fibres (Aguzzi & Heikenwalder, 2006). Nevertheless, no definitive information is available on the cellular basis for prion transmigration from gut to ENS plexuses, nor on the ENS cell types colonized by TSE agents. Likewise, the morphofunctional changes affecting the ENS cell populations during infection are unknown, despite the existence of studies addressing CNS neuron damage and targeting on behalf of TSE agents (Guentchev et al., 1999). This study was aimed at characterizing the ENS cells that are targeted during natural and oral experimental scrapie infection in Sarda sheep. Major emphasis was placed upon enteric glial cells (EGCs) and two neuron populations expressing calbindin (CALB) and neuronal nitric oxide synthase (nNOS), respectively. In sheep, CALB-immunoreactive (IR) cells account for 20–25 % of myenteric plexus (MP) and 65–75 % of submucosal plexus (SMP) neurons, most of them corresponding to cholinergic Dogiel type II neurons and probably acting as intrinsic primary afferent neurons (Chiocchetti et al., 2004, 2006). Enteric nNOS-IR cells, accounting for 31–36 % of MP and 22–24 % of SMP neurons (Lalatta-Costerbosa et al., 2007), correspond to Dogiel type I cells, probably acting as inhibitory motoneurons (Pfannkuche et al., 2002). Another objective was to evaluate whether neuron loss occurs within ileal ENS plexuses from naturally and orally experimentally scrapie-affected Sarda sheep. Indeed, whilst CNS neuron loss is constantly observed in TSEs (Pocchiari, 1994), no data concerning ENS are available. Ileal MPs and SMPs were obtained from 20 Sarda sheep carrying different PrP genotypes, which were characterized as reported elsewhere (Vaccari et al., 2001). Twelve sheep (three ARQ/ARQ, seven ARR/ARQ and two ARR/ARR animals), aged 2–4 years and originating from a scrapiefree flock, acted as healthy controls and were slaughtered according to standard procedures, with tissue samples (cerebral obex, palatine tonsils and distal ileum) being collected from them. The remaining animals included four ARQ/ARQ naturally scrapie-affected and four additional ARQ/ARQ sheep that had been dosed orally at 20 days of age with 25 ml 20 % scrapie brain homogenate. These animals were euthanized, in accordance with approved protocols, at the terminal stage of disease (between 2 and 5 years), with the same tissues being collected. The diagnosis of scrapie was confirmed or excluded by immunohistochemistry (IHC) and Western blot (WB) analysis, which were carried out on the obex, tonsils and distal ileum, in agreement with published protocols (Ligios et al., 2006). Ileal samples were fixed in 10 % neutral-buffered formalin, embedded in paraffin and cut into 5 mm thick sections, both transverse and tangential, which were placed on glass slides coated with (3-aminopropyl)triethoxy-silane (Sigma-Aldrich). 2900 PrPd IHC was performed with a mouse monoclonal antibody (mAb) (F99/97.6.1; VMRD, Inc.). A pre-treatment protocol was applied to abolish PrPC immunoreactivity without damaging other epitopes and preserving tissue morphology (Kovács et al., 2005). Sections were rinsed in 96 % formic acid for 2 min at room temperature and autoclaved at 100 uC for 20 min in 0.01 M citrate buffer (pH 6.0). Antibody binding was detected by using a biotin–avidin method and immune reactions were visualized by 3-39-diaminobenzidine (DAB) chromogen solution (both from Vector Laboratories, Inc.). Control sections included obex, tonsil and distal ileum from scrapie-infected and uninfected Sarda sheep. Additional negative controls were set up by omitting the primary antibody. Double-labelling indirect immunofluorescence (DLIIF) was carried out with a panel of commercial mAbs and polyclonal (Po)Abs against PrPC (F99/97.6.1 mAb), glial fibrillary acidic protein, an EGC marker (anti-GFAP PoAb), neuron-specific enolase, a neuron cell marker (anti-NSE PoAb), nNOS and CALB (both PoAbs) (see Supplementary Table S1, available in JGV Online, for details). The same protocol to abolish PrPC immunoreactivity was employed in DLIIF and then two primary antibodies were added to sections and incubated overnight at room temperature. PrPd was detected by an enhanced biotin–avidin protocol (Vector Laboratories, Inc.) and sections were incubated with goat anti-mouse and antirabbit biotinylated secondary antibodies for 30 min. Immune reactions were visualized by Texas red avidin DCS (D cell sorter) and fluorescein avidin DCS fluorochromes (Vector Laboratories, Inc.). Control sections included obex, tonsil and distal ileum from scrapieinfected and uninfected Sarda sheep. Additional negative controls were set up by omitting primary antibodies. Sections were examined under a Nikon Eclipse 800 microscope equipped with fluorescence and images were collected with a Nikon DXM 1200 digital camera. Colocalization analyses were performed by ImageJ software (National Institutes of Health). To estimate the density (cells mm22) of total (HuC/D-IR), nNOS-IR and CALB-IR neurons within ileal MPs, wholemount (WM) preparations were made. We utilized a protocol originally developed in the rat (Phillips et al., 2004), adapting it to the sheep ileum, where MPs show a more variable and less geometric texture (Gabella, 1987) and MP neurons are often gathered in large ganglia exhibiting a polygonal or ring-like morphology, with no apparent directionality (Lalatta-Costerbosa et al., 2007). Beginning 2 cm oral to the ileocaecal junction, we collected from each animal 8–10 cm long segments, which were cut along the mesenteric border, pinned on balsa board and fixed in 2 % paraformaldehyde plus 0.2 % picric acid in PBS (pH 7.0) at 4 uC overnight, before being washed three times in DMSO (10 min each) and stored at 4 uC in PBS containing sodium azide (0.1 %). Segments were subdivided into six transverse columns, with a 161 cm Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sun, 07 May 2017 13:45:51 Journal of General Virology 88 Role of the enteric nervous system in scrapie sample being collected from each column at different levels, to achieve an adequate representation of the entire circumference (from anti-mesenteric to mesenteric border) and length of every segment. WM preparations were obtained by removing the mucosa, submucosa and circular muscle layer from each sample. For DLIIF investigations, MPs were incubated in 10 % normal goat serum in PBS containing 1 % Triton X-100 for 30 min at room temperature. An anti-HuC/D mAb was used as panneuron marker (Chiocchetti et al., 2004; Phillips et al., 2004), whereas CALB-IR and nNOS-IR neurons were detected with specific PoAbs (see Supplementary Table S1, available in JGV Online, for details). Samples were challenged simultaneously with two primary antibodies for 40 h at 4 uC, washed three times in PBS (10 min each) and finally incubated with goat anti-mouse and anti-rabbit secondary antibodies conjugated with Alexa 594 (Molecular Probes) and fluorescein isothiocyanate (FITC) 492 (Calbiochem) fluorochromes, respectively. small-sized or faintly stained cells, MP neurons were counted by two independent investigators blind to the experimental conditions at 640 magnification with a Zeiss Axioplan microscope equipped with appropriate filters discriminating the fluorochromes used. Conventionally, neurons intersected by the upper and lower fields’ hemicircumferences were, respectively, disregarded and considered (De Souza et al., 1993). For each of the six samples, the number of total (HuC/DIR), CALB-IR and nNOS-IR cells was counted in 12 microscope fields (each field, 0.28 mm2), which had previously been determined by means of two orthogonal coordinates taken from a table of random numbers and measured on the movable stage of the microscope. Therefore, for each stain per plexus per animal, a total area of 10.08 mm2 was evaluated. To avoid missing All scrapie-affected sheep showed IHC evidence of PrPd deposition in ileal MPs, SMPs and PPs, as well as in tonsils and obex. WB analysis confirmed the above results. PrPd accumulation was more prominent at the MP level, with a ‘pepper granule-like’ pattern strongly compatible with EGCs. A simultaneous involvement of neuronal perikarya was observed. No specific immunolabelling was present in negative-control sections. By DLIIF, PrPd deposition All quantitative data collected, not grouped for control/ infected status and PrP genotype, were tested for normality with a Shapiro–Wilk W test. As the Shapiro–Wilk W test did not demonstrate a normal distribution, comparisons among healthy controls carrying different PrP genotypes and between controls – irrespective of PrP genotype – and scrapie-affected sheep were evaluated with Spearman’s rank correlation test. Statistical analyses were performed by using CSS software (StatSoft) and a conventional 5 % level was used to define statistical significance. (a) (b) (c) (d) (e) (f) Fig. 1. Naturally scrapie-affected Sarda sheep. Distal ileum (tangential section). (a–c) Specific anti-CALB (a) and anti-PrPd (b) immunostaining is shown within an SMP neuron. Colocalization of the two signals is shown in white (c). (d–f) Specific antinNOS (d) and anti-PrPd (e) immunolabelling is observed within two neuronal perikarya and nerve fibres from an MP. Colocalization of the two signals is shown in white (f). DLIIF with anti-CALB, anti-nNOS and anti-PrP antibodies was performed. Fluorescein avidin DCS (green) and Texas red avidin DCS (red) were used as fluorochromes. Colocalization analyses were carried out by means of ImageJ software. Bars, 50 mm. http://vir.sgmjournals.org Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sun, 07 May 2017 13:45:51 2901 G. Marruchella and others (a) (b) (c) Fig. 2. Naturally scrapie-affected Sarda sheep. Distal ileum (tangential section). Specific anti-GFAP (a) and anti-PrPd (b) immunostaining is shown within the EGCs of a large MP. Colocalization of the two signals is shown in white (c). Fluorescein avidin DCS (green) and Texas red avidin DCS (red) were used as fluorochromes. Colocalization analyses were carried out by means of ImageJ software. Bars, 50 mm. involved EGCs and neurons (NSE-IR cells) of MPs and SMPs from the eight scrapie-affected sheep. Both CALB-IR and nNOS-IR neurons (Fig. 1) harboured discrete, granular PrPd aggregates in cell bodies and fibres, with no apparent quantitative differences between these cell populations. In contrast, EGCs exhibited more prominent and diffuse PrPd accumulations (Fig. 2). Negative-control sections showed no evidence of PrPd deposition. No statistically significant differences in the density of total, CALB-IR and nNOS-IR neurons were observed within ileal MPs among the different PrP genotypes of the 12 control sheep. Also, no statistically significant differences were found between the eight scrapie-affected and the 12 control sheep – irrespective of PrP genotype – when densities of total, CALB-IR and nNOS-IR neurons were compared (Fig. 3). deposition and for the progression of infection (Blattler et al., 1997). EGCs are the morphofunctional equivalent of CNS astrocytes (Cabarrocas et al., 2003), which harbour PrPd earlier than CNS neurons (Diedrich et al., 1991) and probably play a role in prion-induced neuron damage (Raeber et al., 1997; Jeffrey et al., 2004). We found no differences in the density of total, CALB-IR and nNOS-IR neurons among control sheep with different PRNP polymorphisms. This suggests that the host’s PrP genotype neither affects the density and/or the neurochemical code of ileal MP neurons in Sarda sheep, as was reported previously for nNOS-IR cells (Lalatta-Costerbosa et al., 2007), nor does it modulate scrapie susceptibility/ resistance by influencing MP features. As no differences were detected between scrapie-affected and scrapie-negative sheep, our data also suggest that ENS neurons may This study supports the assumption that ileal ENS plexuses are involved in the pathogenesis of both natural and oral experimental scrapie infection in sheep. However, as all scrapie-affected animals were investigated at clinicaldisease end stage, we cannot exclude the possibility that ENS plexuses might have also been colonized by centrifugal spread, from CNS to gut. By IHC, granular PrPd deposits were detected within ileal MPs and SMPs from scrapieaffected sheep. As reported previously (Heggebø et al., 2003; Jeffrey et al., 2006), the accumulation pattern was mainly consistent with EGCs, although neuronal perikarya and fibres also showed evidence of PrPd deposition. DLIIF and colocalization analyses confirmed PrPd accumulation in EGCs, CALB-IR and nNOS-IR neurons. Neurons of ENS plexuses express PrPC (Heggebø et al., 2000; Shmakov et al., 2000) and may show PrPd deposition in different TSEs (Andréoletti et al., 2000; McBride et al., 2001; Sigurdson et al., 2001; van Keulen et al., 2002). However, PrPd was more consistently detectable within EGCs, which in humans were also shown to express PrPC (Shmakov et al., 2000), a crucial prerequisite for PrPd 2902 Fig. 3. Mean densities of total (HuC/D-IR), CALB-IR and nNOSIR neurons within ileal MPs from the 12 control Sarda sheep carrying different PrP genotypes and from the eight ARQ/ARQ (naturally and experimentally) scrapie-affected sheep. No statistically significant differences (P¡0.05) were detected between healthy-control (filled bars) and scrapie-affected (empty bars) animals (Spearman’s rank correlation test). Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sun, 07 May 2017 13:45:51 Journal of General Virology 88 Role of the enteric nervous system in scrapie interact with the infectious agent in a unique way. Indeed, it is widely accepted that CNS neuron loss represents a dominant feature in TSEs (Pocchiari, 1994). Evidence exists that microglial cells are involved in the pathogenesis of TSE-associated neuron damage at brain level (Peyrin et al., 1999; Pasquali et al., 2006; Priller et al., 2006), but ENS plexuses do not physiologically host a microglial component, a plausible explanation for the apparent lack of neuron loss in our scrapie-affected sheep. We cannot, however, exclude the possibility that qualitative changes were still affecting ENS neurons. In conclusion, this study showed that EGCs and neurons, particularly CALB-IR and nNOS-IR cells, within ileal ENS plexuses accumulate PrPd during natural and oral experimental scrapie infection in Sarda sheep, without apparent neuron loss. This intriguing observation warrants further study on the complex biological interplay between prions and neurons. Diedrich, J. F., Bendheim, P. E., Kim, Y. S., Carps, R. I. & Haase, A. T. (1991). Scrapie-associated prion protein accumulates in astrocytes during scrapie infection. Proc Natl Acad Sci U S A 88, 375–379. Gabella, G. (1987). The number of neurons in the small intestine of mice, guinea-pigs and sheep. Neuroscience 22, 737–752. Goldmann, W., Hunter, N., Smith, G., Foster, J. & Hope, J. (1994). PrP genotype and agent effects in scrapie: change in allelic interaction with different isolates of agent in sheep, a natural host of scrapie. J Gen Virol 75, 989–995. Guentchev, M., Wanschitz, J., Voigtländer, T., Flicker, H. & Budka, H. (1999). Selective neuronal vulnerability in human prion diseases. Fatal familial insomnia differs from other types of prion diseases. Am J Pathol 155, 1453–1457. Heggebø, R., Press, C., Gunnes, G., Lie, K. I., Tranulis, M. A., Ulvund, M., Groschup, M. H. & Landsverk, T. (2000). Distribution of prion protein in the ileal Peyer’s patch of scrapie-free lambs and lambs naturally and experimentally exposed to the scrapie agent. J Gen Virol 81, 2327–2337. Heggebø, R., González, L., Press, C., Gunnes, G., Espenes, A. & Jeffrey, M. (2003). Disease-associated PrP in the enteric nervous system of scrapie-affected Suffolk sheep. J Gen Virol 84, 1327–1338. Jeffrey, M., Goodsir, C. M., Race, R. E. & Chesebro, B. (2004). Acknowledgements Scrapie-specific neuronal lesions are independent of neuronal PrP expression. Ann Neurol 55, 781–792. The experimentally scrapie-affected sheep were kindly provided by Dr Ciriaco Ligios, being part of another research project funded by the European Union. We also thank Mrs Marina Baffoni for outstanding technical assistance. This work was carried out with research grants (PRIN 2004, 2006) from Ministero per l’Istruzione, l’Università e la Ricerca, the Universities of Teramo and Bologna and Fondazione del Monte di Bologna e Ravenna. Jeffrey, M., Gonzalez, L., Espenes, A., Press, C., Martin, S., Chaplin, M., Davis, L., Landsverk, T., MacAldowie, C. & other authors (2006). Transportation of prion protein across the intestinal mucosa of scrapiesusceptible and scrapie-resistant sheep. J Pathol 209, 4–14. Kovács, G. G., Preusser, M., Strohschneider, M. & Budka, H. (2005). Subcellular localization of disease-associated prion protein in the human brain. Am J Pathol 166, 287–294. Lalatta-Costerbosa, G., Mazzoni, M., Clavenzani, P., Di Guardo, G., Mazzuoli, G., Marruchella, G., De Grossi, L., Agrimi, U. & Chiocchetti, R. (2007). Nos-immunoreactivity and NADPH-d References Aguzzi, A. & Heikenwalder, M. (2006). Pathogenesis of prion diseases: current status and future outlook. Nat Rev Microbiol 4, 765–775. Aguzzi, A. & Polymenidou, M. (2004). Mammalian prion biology: one century of evolving concepts. Cell 116, 313–327. Andréoletti, O., Berthon, P., Marc, D., Sarradin, P., Grosclaude, J., van Keulen, L., Schelcher, F., Elsen, J. M. & Lantier, F. (2000). Early accumulation of PrPSc in gut-associated lymphoid and nervous tissues of susceptible sheep from a Romanov flock with natural scrapie. J Gen Virol 81, 3115–3126. Blattler, T., Brandner, S., Raeber, A. J., Klein, M. A., Voigtlander, T., Weissman, C. & Aguzzi, A. (1997). PrP-expressing tissue required for transfer of scrapie infectivity from spleen to brain. Nature 389, 69–73. histochemistry in the enteric nervous system of Sarda breed sheep with different PrP genotypes in wholemount and cryostat preparations. J Histochem Cytochem 55, 387–401. Ligios, C., Cancedda, M. G., Madau, L., Santucciu, C., Maestrale, C., Agrimi, U., Ru, G. & Di Guardo, G. (2006). PrPSc deposition in nervous tissues without lymphoid tissue involvement is frequently found in ARQ/ARQ Sarda breed sheep preclinically affected with natural scrapie. Arch Virol 151, 2007–2020. Mabbott, N. A. & MacPherson, G. (2006). Prions and their lethal journey to the brain. Nat Rev Microbiol 4, 201–211. McBride, P. A., Schulz-Schaeffer, W. J., Donaldson, M., Bruce, M., Diringer, H., Kretzschmar, H. A. & Beekes, M. (2001). Early spread of glial cells in inflammatory bowel disease. Glia 41, 81–93. scrapie from the gastrointestinal tract to the central nervous system involves autonomic fibers of the splanchnic and vagus nerves. J Virol 75, 9320–9327. Chiocchetti, R., Grandis, A., Bombardi, C., Clavenzani, P., Lalatta Costerbosa, G., Lucchi, M. L. & Furness, J. B. (2004). Pasquali, P., Nonno, R., Mandara, M. T., Di Bari, M. A., Ricci, G., Petrucci, P., Cappuccini, S., Cartoni, C., Macrì, A. & Agrimi, U. (2006). Cabarrocas, J., Savidge, T. R. & Liblau, R. S. (2003). Role of enteric Characterisation of neurons expressing calbindin immunoreactivity in the ileum of the unweaned and mature sheep. Cell Tissue Res 318, 289–303. Chiocchetti, R., Grandis, A., Bombardi, C., Lucchi, M. L., Tadini Dal Lago, D., Bortolami, R. & Furness, J. B. (2006). Extrinsic and intrinsic sources of calcitonin gene-related peptide immunoreactivity in the lamb ileum: a morphometric and neurochemical investigation. Cell Tissue Res 323, 183–196. Intracerebral administration of interleukin-12 (IL-12) and IL-18 modifies the course of mouse scrapie. BMC Vet Res 2, 37. Peyrin, J. M., Lasmezas, C. I., Haik, S., Tagliavini, F., Salmona, M., Williams, A., Richie, D., Deslys, J. P. & Dormont, D. (1999). Microglial cells respond to amyloidogenic PrP peptide by the production of inflammatory cytokines. Neuroreport 10, 723–729. Pfannkuche, H., Schemann, M. & Gäbel, G. (2002). Ruminal muscle De Souza, R. R., Moratelli, H. B., Borges, N. & Liberti, E. A. (1993). of sheep is innervated by non-polarized pathways of cholinergic and nitrergic myenteric neurones. Cell Tissue Res 309, 347–354. Age-induced nerve loss in the myenteric plexus of the small intestine in man. Gerontology 39, 183–188. Phillips, R. J., Hargrave, S. L., Rhodes, B. S., Zopf, D. A. & Powley, T. L. (2004). Quantification of neurons in the myenteric plexus: an http://vir.sgmjournals.org Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sun, 07 May 2017 13:45:51 2903 G. Marruchella and others evaluation of putative pan-neuronal markers. J Neurosci Methods 133, 99–107. is expressed in the human enteric nervous system. Nat Med 6, 840–841. Pocchiari, M. (1994). Prions and related neurological diseases. Mol Aspects Med 15, 195–291. Sigurdson, C. J., Spraker, T. R., Miller, M. W., Oesch, B. & Hoover, E. A. (2001). PrPCWD in the myenteric plexus, vagosympathetic trunk Priller, J., Prinz, M., Heikenwalder, M., Zeller, N., Schwarz, P., Heppner, F. L. & Aguzzi, A. (2006). Early and rapid engraftment and endocrine glands of deer with chronic wasting disease. J Gen Virol 82, 2327–2334. of bone-marrow-derived microglia in scrapie. J Neurosci 26, 11753–11762. Vaccari, G., Petraroli, R., Agrimi, U., Eleni, C., Perfetti, M. G., Di Bari, M. A., Morelli, L., Ligios, C., Busani, L. & other authors (2001). PrP Prusiner, S. B. (1998). Prions. Proc Natl Acad Sci U S A 95, genotype in Sarda breed sheep and its relevance to scrapie. Brief report. Arch Virol 146, 2029–2037. 13363–13383. Raeber, A. J., Race, R. E., Brandner, S., Priola, S. A., Sailer, A., Bessen, R. A., Mucke, L., Manson, J., Aguzzi, A. & other authors (1997). Astrocyte-specific expression of hamster prion protein (PrP) van Keulen, L. J. M., Vromans, M. E. W. & van Zijderveld, F. G. (2002). renders PrP knockout mice susceptible to hamster scrapie. EMBO J 16, 6057–6065. Westaway, D., Zuliani, V., Cooper, C. M., Da Costa, M., Neuman, S., Jenny, A. L., Detwiler, L. & Prusiner, S. B. (1994). Homozygosity for Shmakov, A. N., McLennan, N. F., McBride, P., Farquhar, C. F., Bode, J., Renninson, K. A. & Ghosh, S. (2000). Cellular prion protein prion protein alleles encoding glutamine-171 renders sheep susceptible to natural scrapie. Genes Dev 8, 959–969. 2904 Early and late pathogenesis of natural scrapie infection in sheep. APMIS 110, 23–32. Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sun, 07 May 2017 13:45:51 Journal of General Virology 88