Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
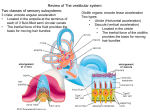
The Vestibular System: Or, why we don’t fall in the dark! The vestibular organs sense head motion (anterior) (lateral) Semicircular canals sense head rotations - Angular accelerations Otoliths sense linear accelerations - Gravity and other linear accelerations The function of the vestibular system can be simplified by remembering some basic terminology of classical mechanics. All bodies moving in a three-dimensional framework have six degrees of freedom: three of these are translational and three are rotational. The translational elements refer to linear movements in the x, y, and z axes (the horizontal and vertical planes). Translational motion in these planes (linear acceleration and static displacement of the head) is the primary concern of the otolith organs. The three degrees of rotational freedom refer to a body's rotation relative to the x, y, and z axes and are commonly referred to as roll, pitch, and yaw. The semicircular canals are primarily responsible for sensing rotational accelerations around these three axes. Vestibular outputs very rapidly influence eye, head, and postural reflexes • Vestibulo-ocular reflex – Eye velocity compensates for head velocity • Vestibulo-colic – Head position maintained despite body movements • Vestibulospinal – Postural changes in response to vestibular signals Scanning electron micrograph of hair cells from the bullfrog inner ear. The mechanisms underlying the depolarization and hyperpolarization of vestibular hair cells depend, respectively, on the potassium-rich character of endo lymph and the potassium-poor character of the perilymph that bathes the basal and lateral portions of the hair cells. Deflection of the stereocilia toward the kino cilium causes potassium channels in the apical portions of the stereocilia and kinocilium to open. K+ flows into the cell from the endolymph, depolarizing the cell membrane (see Fig. 7). This depolarization in turn causes voltage-gated calcium channels at the base of the hair cells to open, allowing Ca++ to enter the cell. The influx of Ca++ causes synaptic vesicles to release their transmitter (aspartate or glutamate) into the syn aptic clefts, and the afferent fibers respond by under going depolarization and increasing their rate of firing. When the stimulus subsides, the stereocilia and kinocilium return to their resting position, allowing most calcium channels to close and voltagegated potassium channels at the base of the cell to open. K+ efflux returns the hair cell membrane to its resting potential (see Fig. 7). Deflection of the stereocilia away from the kino cilium causes potassium channels in the basolateral portions of the hair cell to open, allowing K+ to flow out from the cell into the interstitial space. The result ing hyperpolarization of the cell membrane decreases the rate at which the neurotransmitter is released by the hair cells and consequently, decreases the firing rate of afferent fibers. Almost all vestibular primary afferent fibers have a moderate spontaneous firing rate at rest (approximately 90 spikes per second). Therefore, it is likely that some hair cell calcium channels are open at all times, causing a slow, constant release of neurotransmitter. The ototoxic effects of some aminoglycoside antibiotics (e.g., streptomycin, gentamicin) may be due to direct reduction of the transduction currents of hair cells. From: Dickman at http://vestibular.wustl.edu/vestibular2.html Linear acceleration Figure 14.2. The morphological polarization of vestibular hair cells and the polarization maps of the vestibular organs. (A) A cross section of hair cells shows that the kinocilia of a group of hair cells are all located on the same side of the hair cell. The arrow indicates the direction of deflection that depolarizes the hair cell. (B) View looking down on the hair bundles. (C) In the ampulla located at the base of each semicircular canal, the hair bundles are oriented in the same direction. In the sacculus and utricle, the striola divides the hair cells into populations with opposing hair bundle polarities. Scanning electron micrograph of calcium carbonate crystals (otoconia) in the utricular macula of the cat. Each crystal is about 50 µm long. (From Lindeman, 1973.) In humans, they are 3 - 30 µm long. Figure 14.4. Morphological polarization of hair cells in the utricular and saccular maculae. (A) Cross section of the utricular macula showing hair bundles projecting into the gelatinous layer when the head is level. (B) Cross section of the utricular macula when the head is tilted. (C) Orientation of the utricular and saccular maculae in the head; arrows show orientation of the kinocilia, as in Figure 14.2. The saccules on either side are oriented more or less vertically, and the utricles more or less horizontally. The striola is a structural landmark consisting of small otoconia arranged in a narrow trench that divides each otolith organ. In the utricular macula, the kinocilia are directed toward the striola. In the saccular macula, the kinocilia point away from the striola. Note that, given the utricle and sacculus on both sides of the body, there is a continuous representation of all directions of body movement. Figure 14.5. Forces acting on the head and the resulting displacement of the otolithic membrane of the utricular macula. For each of the positions and accelerations due to translational movements, some set of hair cells will be maximally excited, whereas another set will be maximally inhibited. Note that head tilts produce displacements similar to certain accelerations. Figure 14.6. Response of a vestibular nerve axon from an otolith organ (the utricle in this example). (A) The stimulus (top) is a change that causes the head to tilt. The histogram shows the neuron's response to tilting in one direction. (B) A response of the same fiber to tilting in the opposite direction. (After Goldberg and Fernandez, 1976.) Angular acceleration Figure 14.7. The ampulla of the posterior semicircular canal showing the crista, hair bundles, and cupula. The cupula is distorted by the fluid in the membranous canal when the head rotates. Figure 14.8. Functional organization of the semicircular canals. (A) The position of the cupula without angular acceleration. (B) Distortion of the cupula during angular acceleration. When the head is rotated in the plane of the canal (arrow outside canal), the inertia of the endolymph creates a force (arrow inside the canal) that displaces the cupula. (C) Arrangement of the canals in pairs. The two horizontal canals form a pair; the right anterior canal (AC) and the left posterior canal (PC) form a pair; the left AC and the right PC form a pair. RALP LARP http://vestibular.wustl.edu/vestibular.html Vestibular nerves signal head velocity The viscous damping of the semicircular canals result in the vestibular nerves carrying signals proportional to head velocity. Responses of a left AC afferent (MFR 76 spikes/s, CV* = 0.04) to earth-horizontal axis rotation. The bottom trace represents stimulus head velocity, 0.5 Hz, with a peak amplitude of 31.2°/s. From Haque A, Angelaki DE, Dickman Exp Brain Res. 2004 155:81-90. Vestibulo-ocular reflex Figure 14.10. Connections underlying the vestibulo-ocular reflex. Projections of the vestibular nucleus to the nuclei of cranial nerves III (oculomotor) and VI (abducens). The connections to the oculomotor nucleus and to the contralateral abducens nucleus are excitatory (red), whereas the connections to ipsilateral abducens nucleus are inhibitory (black). There are connections from the oculomotor nucleus to the medial rectus of the left eye and from the adbucens nucleus to the lateral rectus of the right eye. This circuit moves the eyes to the right, that is, in the direction away from the left horizontal canal, when the head rotates to the left. Turning to the right, which causes increased activity in the right horizontal canal, has the opposite effect on eye movements. The projections from the right vestibular nucleus are omitted for clarity. VOR gain is low at low frequencies Cupula signaling ability decreases with a 5 second time constant Vestibular projections to the spinal cord Figure 14.11. Descending projections from the medial and lateral vestibular nuclei to the spinal cord. The medial vestibular nuclei project bilaterally in the medial longitudinal fasciculus to reach the medial part of the ventral horns and mediate head reflexes in response to activation of semicircular canals. The lateral vestibular nucleus sends axons via the lateral vestibular tract to contact anterior horn cells innervating the axial and proximal limb muscles. Neurons in the lateral vestibular nucleus receive input from the cerebellum, allowing the cerebellum to influence posture and equilibrium. Lateral Vestibulospinal Tract Lateral Vestibular nucleus Lateral Vestibulospinal Tract: • The inputs from the otolith organs project mainly to the lateral vestibular nucleus, which in turn sends axons in the lateral vestibulospinal tract to the spinal cord. • The input from this tract exerts a powerful excitatory influence on the extensor (antigravity) muscles. When hair cells in the otolith organs are activated, signals reach the medial part of the ventral horn. By activating the ipsilateral pool of motor neurons innervating extensor muscles in the trunk and limbs, this pathway mediates balance and the maintenance of upright posture. • Decerebrate rigidity, which is characterized by rigid extension of the limbs, arises when the brainstem is transected above the level of the vestibular nucleus. The tonic activation of extensor muscles in this instance suggests that the vestibulospinal pathway is normally strongly suppressed by descending projections from higher levels of the brain, especially the cerebral cortex. © 2001 by Sinauer Associates, Inc. Medial Vestibulospinal Tract Lateral Vestibular nucleus Medial Vestibulospinal Tract: • Axons from the medial vestibular nucleus descend in the medial longitudinal fasciculus to reach the upper cervical levels of the spinal cord. • This pathway regulates head position by reflex activity of neck muscles in response to stimulation of the semicircular canals from rotational accelerations of the head. • For example, during a downward pitch of the body (e.g., tripping), the superior canals are activated and the head muscles reflexively pull the head up. The dorsal flexion of the head initiates other reflexes, such as forelimb extension and hindlimb flexion, to stabilize the body and protect against a fall. © 2001 by Sinauer Associates, Inc. Vestibular signals are sent to cortex Figure 14.12. Thalamocortical pathways carrying vestibular information. The lateral and superior vestibular nuclei project to the thalamus. From the thalamus, the vestibular neurons project to the vicinity of the central sulcus near the face representation. Sensory inputs from the muscles and skin also converge on thalamic neurons receiving vestibular input. The superior and lateral vestibular nuclei send axons to the ventral posterior nuclear complex of the thalamus, which projects to two cortical areas relevant to vestibular sensations. One cortical target is just posterior to the primary somatosensory cortex, near the representation of the face; another (not shown) is at the transition between the somatic sensory cortex and the motor cortex (Brodmann's area 3a). There are also projections to posterior parietal cortex (Brodmann's area 5). Electrophysiological studies of individual neurons in these areas show that the relevant cells respond to proprioceptive and visual stimuli as well as to vestibular stimuli. Many of these neurons are activated by moving visual stimuli as well as by rotation of the body (even with the eyes closed), suggesting that these cortical regions are involved in the perception of body orientation in extrapersonal space.