Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
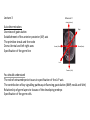
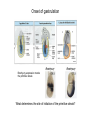
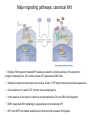
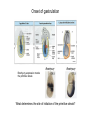
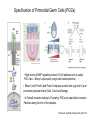
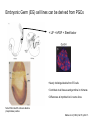
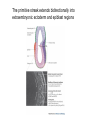
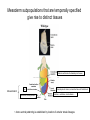
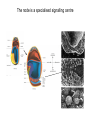
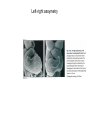
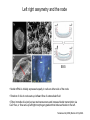
Lecture 3 Where am I? Anterior (Head) Axis determination. Overview of gastrulation Establishment of the anterior-posterior (AP) axis The primitive streak and the node Dorso-Ventral and left-right axes Specification of the germ line Right Dorsal (Back) Ventral (Front) Left Posterior (Tail) You should understand The role of extraembryonic tissues in specification of the A-P axis The contribution of key signalling pathways influencing gastrulation (BMP, nodal and Wnt) Relationship of germ layers to tissues of the developing embryo Specification of the germ cells. Germ layers, Ectoderm, Mesoderm, and Endoderm, give rise to all tissues of the developing embryo Blastocyst Development of the egg cylinder Onset of gastrulation Brachyury expression marks the primitive streak What determines the site of initiation of the primitive streak? Major signalling pathways; BMP and Nodal/Activin • BMPs and Nodal signal by binding type I/II receptor and activating ser/thr kinase to phosphorylate Smads • BMP signal transduced by phosphorylating Smad 1,5, or 8 and Nodal through Smad 2 or 3 • Phospho Smads bind Smad4, translocate to the nucleus and activate target genes Major signalling pathways; Nodal fine tuning • Nodal can be regulated at the level of conversion of pro-nodal to nodal by Furin/PACE4 • Cer1 and Lefty 1 are diffusible antagonists of nodal. Major signalling pathways; canonical Wnt • Binding of Wnt ligand to frizzled/LRP stabilises b-catenin by blocking activity of the destruction complex comprising Axin, Dvl, and the kinases CK1 gamma and GSK beta. • Stabilised b-catenin translocates to the nucleus, binds to TCF family proteins and activates expression • In the absence of b-catenin TCF proteins repress target genes. • In the absence of wnt ligand, b-catenin is phosphorylated by CK1 and GSK3 and degraded • DKK1 anagonises Wnt signalling by sequestering and internalising LRP • WIF1 and sFRP are frizzled related proteins that bind and sequester Wnt ligands Onset of gastrulation Brachyury expression marks the primitive streak What determines the site of initiation of the primitive streak? Establishment of proximal-distal assymetry The distal visceral endoderm (dve) ? • At E5.5 Nodal is expressed in epiblast cells of early egg cylinder embryos. • At the proximal rim the ExE provides two positive feedback loops (Bmp4 and pro-nodal cleavage) that enhance nodal signalling in the proximal epiblast. • Visceral endoderm cells at the distal tip of the egg cylinder (the dve) respond to Nodal signalling from the epiblast by producing a cocktail of Nodal and Wnt inhibitors, further enhancing the nodal gradient • Unknown inhibitory signals from ExE block nodal responsiveness in other regions of the VE. Rotation of VE moves DVE cells to anterior position (AVE) thus converting P-D axis into A-P axis. Courtesy of Shankar Srinivas, DPAG Signalling from the AVE determines A-P axis orientation. • DVE and then AVE cells secrete diffusible antagonists of Nodal (Lefty1 and Cer-l) and Wnt (DKK1) signalling • Confers anterior characteristics to underlying epiblast (expresses neuroectoderm markers), confirmed by analysis of AVE transplants. • Conversely, epiblast cells retaining high Nodal and Wnt signalling are specified to become mesoderm, marking the initiation of gastrulation. • Nodal and Wnt3 target genes include Fgf8 that triggers epitheilial to mesenchymal transition (EMT) in nascent mesoderm. EMT leads to delamination of nascent mesoderm required for cell movements during primitive streak migration. Thomas and Beddington (1996) Curr Biol. 6, 1487-1496; Brennan et al (2001) Nature 411, p965-969 Evidence from mutants • Nodal mutants fail to specify mesoderm and neuroectoderm • Smad2 in VE required for anterior specification. Mutants specify mesoderm only, eg express brachyury throughout epiblast Development of the germline Specification of Primordial Germ Cells (PGCs) • High levels of BMP signalling instruct 30-40 epiblast cells to adopt PGC fate – Blimp1 expression (single cell transcriptomics). • Blimp1 (with Prmt5) and Prdm14 repress somatic fate (eg hoxb1) and promotes pluripotent fate (Oct4, Sox2 and Nanog). • In Smad2 mutants lacking A-P polarity, PGCs are specified in random Patches along the rim of the epiblast. Ohinata et al (2005), Nature 436, p207-213 Embryonic Germ (EG) cell lines can be derived from PGCs + LIF + bFGF + Steel factor • Nearly indistinguishable from ES cells • Contribute to all tissues and germline in chimeras • Differences at imprinted loci in some lines Note PGCs like ES cells are alkaline phosphatase positive Matsui et al (1992) Cell 70, p841-7. Gastrulation The primitive streak extends bidirectionally into extraembryonic ectoderm and epiblast regions Mesoderm contributes to extraembryonic tissues The primitive streak extends laterally as well as proximo-distally Mesoderm subpopulations that are temporally specified give rise to distinct tissues Muscle and blood of extraembryonic tissues gut definitive endoderm Mesendoderm Notochord, head process Mesenchyme of viscera, connective tissue of limbs,blood. Somites – vertebrae, muscle blocks …. • dorso-ventral patterning is established by location of anterior streak lineages The node is a specialised signalling centre Left-right assymetry Left right assymetry and the node E8.5 • Nodal mRNA is initially expressed equally in cells on either side of the node • Rotation of cilia in node sets up leftward flow of extracellular fluid • Either immotile cilia (red) act as mechanosensors and increase Nodal transcription via Ca2+ flux, or flow sets up left/right morphogen gradient that induces Nodal on the left. Nonaka et al (2002) Nature 418, p96-9. End Lecture 3