Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
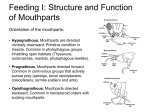
EXTERNAL MORPHOLOGY OF AN ORTHOPTEROID INSECT Morphology is inherent to an understanding of how insects live and function. In contrast to anatomy which is the naming and description of the structural features of an animal, morphology includes the manner in which the parts have been derived, how they function and how they interrelate with similar structures in closely-related animals. A thorough understanding of the morphology of the animal might enable the economic entomologist to predict the manner of egg deposition of a potential pest species, the taxonomist to distinguish between taxonomic groupings, the medical entomologist to predict the method of feeding of some insect vector upon its host, or the phylogenist to ascertain something about the evolution of the group. Orthopteroid insects are good choices for this study because gross morphological features are readily observed. Orthopteroids are considered primitive and demonstrate the basic body plan of most insects. Locate the Bold and underlined words in the following descriptive material. Make certain that you are able to locate each described structure on the specimen itself and avoid copying from a text because the morphological features vary from one species to another and textual material may not necessarily agree fully with the animal before you. As you hold the grasshopper (or other Orthopteroid insect) between your fingers you will note that it is “hard” to the touch, due to the presence of a rigid, leathery EXOSKELETON. If the entire body wall of the animal were heavily sclerotized it could present a distinct problem to the animal for it would not allow freedom of movement. Closer examination of the grasshopper, or any other insect, will reveal many impressed lines and areas of membranes between large sclerotized plates and particularly lines and areas of membranes between large sclerotized plates or around joints of appendages where a great deal of movement is necessary. These impressed lines or grooves in the exoskeleton are termed SUTURES. The term ‘”suture” is a collective term used for any impressed line in the exoskeleton visible externally. The more common type of suture is produced by invagination of the body wall, externally evident as a distinct longitudinal impressed line and internally evident as a ridge. This internal ridges may be used as a strengthening device for the body wall increasing the rigidity or may increase the internal surface area of the body wall for the attachment of muscles in which case they are known as APODEMES. The large, sclerotized, plate-like areas delimited by sutures are termed as SCLERITES. Since certain of the SCLERITES and sutures are consistently present on many groups of insects they are given descriptive terms so we may refer to them when necessary. Many of these have been covered in lecture. Laboratory is your opportunity to see them on an actual insect. General Body Organization Like most other insects, the grasshopper (or other Orthopteroid) consists of three general body regions: the head, thorax and abdomen. (See Figure one.) The abdomen is further subdivided into numerous somites and segments which are readily observed in the Orthopteroids but are often obscured in other insects. These are reminiscent of the original somital organization found in the embryo, and are one of the carryover characteristics in the evolution of both the Annelida and Arthropoda. Unlike the annelids, however, the arthropods have amalgamated several of these primite somites into more workable functional units. The Head The Orthopteroid is quite generalized in structure with typical mandibulate, or chewing mouthparts. Although it exhibits some specialization, the Orthopteroid head may be easily compared with a hypothetical insectan head. Label Figure two as you complete this page. The box-like head capsule or cranium consists of several (we said six) embryonic segments which have coalesced to such an extent that the original segmentation is not easily distinguished. Most of the sutures that are present are secondarily developed and are not flexible, functioning primarily as internal bracing mechanisms for the cranium. Thus the cranium is essentially continuously sclerotized except for the posterior portion in which there is a large hole, the foramen magnum, to which the membranous neck (cervix) is attached. The foramen magnum allows for the passage of the gut and other vital internal organs from the head to the posterior regions of the animal. It is, of course, not visible unless the head is detached from the body. The more conspicuous parts of the head include the receptor organs--compound eyes, antennae, and ocelli. Locate the large oval compound eyes and examine them under the dissecting microscope. Note that the surface is divided into many hexagonally-shaped units which represent individual lenses called FACETS, each one functioning individually as an operational eye. In addition to the two compound eyes there are three simple eyes, or ocelli (sing. ocellus). Just median of the compound eyes are the lateral ocelli. The median ocellus is located in the center of the face between the antennae. What is the function of the ocelli? The antennae are the long feelers that are inserted on the upper region of the front of the head. They are perhaps one of the most important organs of the insect body for they are the location for many of the sensory receptors. The nature of these will vary from one insect species to another but commonly the senses of smell, taste, touch and heat perception are located in the antennae. They are variously shaped and undergo considerable modification from group to group, depending largely on the nature of the animal, its method of living, and the environmental conditions in which it normally lives. The various modifications may be grouped into antennal types and are each given descriptive terms useful for the identification of insects. We will look at these general types soon. The frons is a large sclerotized area occupying the entire front of the head from the median ocellus ventrally to a suture, the frontoclypeal suture, below which lies the clypeus. Located in the fontoclypeal suture near the lateral edges of the clypeus are the anterior tentorial pits. These pits represent the points of invagination of the integument to form the anterior arms of the TENTORIUM (the internal supporting skeleton of the head). The posterior tentorial pits are located on the lower part of the back of the head and can be seen better if the head is removed. Below the more or less rectangular-shaped clypeus is a moveable flap termed the labrum, or upper lip. To the sides of the frons and below the compound eyes are genae (sing. gena) or cheeks. Between the gena and the mouth parts is a very small narrow sclerite, the subgena. The dorsum or top part of the head, posterior to the frons, is termed the vertex. The Thorax Use Figure one and, to a lesser extent, Figure four as you look at the thorax. The head is attached to the thorax by the cervix. In the Orthopteroid this is obscured by the enlarged pronotum. The three segments which comprise the thorax are called the prothorax, mesothorax, and methathorax. Fundamentally, each segment is made up of three regions: The tergum or dorsal portion (in a thoracic segment it is referred to as a notum), the sternum, or ventral portion, and the pleura or side portions. From a theoretical aspect each of the pleura can be further subdivided by a median dorso-ventral suture which divides the sclerite into an anterior sclerite known as the episternum and a posterior sclerite known as the epimeron. You will note that the thorax bears the wings and the legs which are diagnostic and which delimit the thorax, no matter how fused the segments may become with more highly evolved insects. Note that there are three sets of legs, again reminiscent of the three primitive somites from which this region of the body has evolved. The prothorax is the thoracic segment directly behind the head of the insect. The tergal part of the prothorax is bonnet-shaped and extends over the dorsal and lateral regions of the segment practically obliterating the pleura, and also the tergum of the mesothorax. It is called the pronotum. The pronotum of the Orthopteriod is greatly enlarged. While this is common for many insects, it is not by any means the usual condition. The pronotum may be large, average, or very small in size. On each side of the body at the lower edge of the pronotum is found a smaller triangular sclerite located just in front of the point where the leg is attached. This is the episternum. It is the only visible part of the pleural region in the prothorax. The prothorax carries one pair of legs, the prothoracic legs. The mesothorax is the second thoracic segment. Its anterior margin can be readily seen by raising the posterior edge of the pronotum. The second pair of legs or mesothoracic legs originate from the lower part of the mesothorax. On the upper part of this segment is found the first pair of wings, the mesothoracic wings. (Figure four). In the cricket and other Orthoptera they are parchment-like and are called tegmina (Sing. tegmen). On the side of the body a distinct suture can be seen extending obliquely forward from the posterior border of the mesothoracic leg to the hind margin of the mesothoracic wing. This suture divides the mesothorax from the following thoracic segments, the metathorax. As previously pointed out, each pleuron or lateral region of the mesothorax is divided by a pleural suture into an anterior sclerite, the episternum, and a posterior sclerite, the epimeron. The pleural suture extends from coxal cavity of the leg up to the base of the wing. On each side of the body a small mesothoracic spiracle, an outlet of the insects gas exchange system, is located on the membrane connecting with prothorax and mesothorax on each side. This spiracle lies under a white membranous flap and can be seen by pushing the flap with a pin. The last thoracic segment, the metathorax, is very similar to the mesothorax. It bears the second pair of wings, the metathoracic wings. (Figure four). Compare the two pairs of wings as to size and texture. The wing-bearing segments of the thorax are collectively termed the pterothorax. The metathorax also bears the third pair of legs. These are used chiefly for jumping, while the first and second pair of legs are used primarily for walking. Again, each pleural area of the metathorax is divided by an oblique pleural suture into an episternum and an epimeron. The ventral sclerite of the metathorax is the metasternum. The spiracle seen just above the base of the mesothoracic leg is the metathoracic spiracle which is moved forward until it seems to lie between the meso - and metathorax. Ventrally, the insectan thorax follows a repetitive pattern of sclerites which are found between the leg bases and again are indicative of the original somital organization. The ventral sclerite is known as the sternum, and by applying the prefixes pro-, meso-, and meta-, you can again indicate a particular segment of the thorax. The Thoracic Appendages The legs of insects represent the primitive appendages; the wings are secondly acquired. The legs of insects vary greatly. Many of the modifications are extremely functional and easily recognized as such. The many degrees of specialization present excellent diagnostic characters and consequently these variations are used extensively in classification. Make a comparative study of the legs in the orthopteroid (that is, find the series of parts on all three legs). Each leg will be found to consist of five serially arranged segments. Label Figure four as you go. The first segment, which is attached to the thorax by a tough elastic membrane, is the short coxa. This segment is most distinct on the prothoracic legs of the orthopteroid. The second segment is shorter than the coxa and partially fused with the third. This is the trochanter. It is frequently rather triangular in shape. The third and largest segment is the femur. The fourth segment, the tibia, is more slender than the femur but almost equal to it in length. The remaining three segments re actually subdivisions of a single segment, the tarsus. In the orthopteroid there are three tarsal segments; the number varies in different insects but there are never more than five. These differences are useful as identification characteristics. On the terminal segment is a tarsal claw. Now find a leg slide in the Entomology slide box and draw and label a leg in your notebook. The wings of insects are secondarily acquired; that is, the primitive ancestors did not have wings and our more primitive insect forms today do not have wings. Wings of the type found on insects are found no other place in the animal kingdom, so their presence is a positive identification that a specimen is an insect. The wings do differ greatly, both in number and structure, and these differences, because they fit in definite patterns, are frequently used for identification purposes. The gross differences in number and structure will be observed as the different insect orders are studied. The Abdomen This body division consists of several segments (count them) joined by connective membranes. Each segment is clearly divided into an upper and lower part called the tergum and sternum respectively. The pleura are not always clear as they become fused with the terga; however, in this specimen the pleura are clear. The posterior segments show modification for copulation and oviposition, as explained below. The first abdominal segment appears to have only the tergum present. However, if the grasshopper is examined from the ventral side the sternum of the first abdominal segment will be found. The second to eighth abdominal segments are all quite similar. One pair of spiracles is located at the anterior margin of each segment. How many pairs of spiracles do you find altogether on the abdomen? The abdomen of the female is similar to that of the male through the first seven segments. Ventrally the abdomen terminates with the sternum of the eighth segment which is prolonged posteriorly and forms the subgenital plate. The terga of segments 9 and 10 are partially fused and the tergum of segment 11 is absent as in the male. The ovipositor consists of a pair of saber like blades. The cerci are long filamentous sensory structures. They are common in some insect orders but not in others. MOUTHPARTS OF INSECTS Insects have been extremely successful in adapting to all types of environments and modes of life. This has been accomplished by a multitude of modifications of all body parts from a common primitive plan. Of particular significance have been the structural modifications of the mouthparts, since they determine the type of food utilized by the insect. The mouthparts of insects are of three principal types: 1. Chewing or mandibulate mouthparts, adapted for cutting or grinding solid foods. 2. Combination mouthparts, which can be used for the intake of both solid and liquid foods. 3. Sucking or haustellate mouthparts, adapted for liquid, dissolved or suspended foods. There are intermediate types and therefore such a classification is very arbitrary and artificial. In this course it is not possible to study the mouthparts of immature insects but it should be remembered that they are built on the same general plan although even within the same species they may be entirely different in function and apparently in structure from those of adults. The structural elements which go into the makeup of mouthparts are composed of three pairs of segmental appendages (mandibles, maxillae, and labium) and lobes of the head capsule: the labrum, and the hypopharynx. Mandibulate Mouthparts The primitive mouthparts of insects are chewing or mandibulate mouthparts. They are characterized by more or less triangular mandibles which are utilized to cut, tear, or grind either plant or animal material that is solid in nature, and complex maxillae which are used to manipulate the food. Mouthparts of the orthopteroid (see Figure three) Read through the exercise and examine the mouthparts in place. Then dissect the mouthparts from your specimen, locate the parts mentioned, and be able to recognize them. It is best to turn the specimen ventral side up to observe the mouthparts. As you remove them, place them on a microscope slide as pictured on figure 3. Use the slide resin to prepare a permanent slide. This slide must be turned in when you complete this lab. 1. The labrum (note spelling) or upper lip is the flap-like piece previously referred to as being attached to the ventral edge of the clypeus. This sclerite is no longer a paired appendicular structure. The lower border is notched at the median line. The inner surface of the labrum forms the anterior wall of the preoral cavity. The remaining mouthparts lie behind (posterior to) the labrum. Examine these as to position and attachment before removing any for further study. 2. The mandibles (2). - Note their characteristic shape, points of articulation, and the absence of segmentation. Function? 3. The maxillae (2). - Note that these are complex organs. Attached to the maxilla is a fivesegmented palpus used in a sensory function. Often some of the parts of the maxilla are highly modified into special structures. 4. The hypopharynx (1). - Note its characteristic shape and resemblance to a bulbous tongue. Function? 5. The labium (1) is in reality formed by the fusion of two lateral structures resembling maxillae. 6. The salivarium is the salivary pocket between the hypopharynx and the labium. Combination Mouthparts The combination mouthparts combine well-developed mandibles for chewing and manipulation of solid food with highly modified maxillae and labium for the intake of liquids. Structurally they are considered to be mandibulate mouthparts. Mouthparts of a Honey Bee, Apis mellifera Linnaeus 1. Examine the head of a honeybee. Identify the antennae, ocelli and compound eyes. Holding the specimen in the fingers of your left hand examine the central part of the head below the antennae; this area is the clypeus. At its lower margin find the small flap-like labrum. Life the labrum and observe the two short, hard, spoon-shaped mandibles crossing below it. Separate the two mandibles. Look below the labrum and find a soft flap projecting near its base. This is the epipharnyx. The remaining conspicuous mouth structures form the proboscis of the honey bee and include the soft hypopharynx on the oral surface centrally facing the labrum and epipharynx, the maxillae laterally, and the labium centrally on the aboral (away from the mouth) surface and beyond the hypopharynx on the oral surface. 2. Proboscis - Examine the caudoventral (caudad-toward the tail end of the body along the median line) surface of the head and note the subcranial depression provided with a membranous cuticula (outer covering of chitin). Note how the proboscis can be retracted into and extended out from this depression by moving a number of rod-like sclerite at its base. 3. 4. 5. Maxilla. - The cardo articulates with the dark stripes, the basal part next to the labium. This is continued as the long galea, which is hollowed out on its oral surface. Near the base of the galea note a minute maxillary palpus. Labium. - The main, heavily sclerotized part of the labium is the prementum. In the middle it is continued as the “tongue”, formed of two fused glossae, provided with fine hairs and ending in a small rounded flabellum. Feeding Mechanism. - When the bee feeds on easily accessible liquids the maxillary galaea, the labial palpi, and the glossae are apposed to form a temporary tube. By means of rapid back and forth movement the liquid is drawn up into the tube where it is sucked up to the mouth by a special pump. In confined spaces such as the corolla of a flower, the tongue is thrust out beyond the other mouthparts. Saliva is ejected by means of a salivary syringe. Its outlet is the functional salivary orifice located on the distal extremity of the prementum between the base of the glossal tongue and the rudimentary hypopharynx. The mandibles are used mostly for manipulation of various substances in the hive. Haustellate Mouthparts Mouthparts of the House Fly, Musca domestica Linnaeus 1. Examine a housefly for a very advanced type of mouthparts. The proboscis is divided into three principal parts which are movable on one another. The basal part, in the form of an inverted cone, is known as the rostrum. The median part is cylindrical, provided with an anterior groove, and is known as the haustellum. The apical part, the oral disc, is composed of two broad pads. 2. Examine the rostrum. Note that it is largely membranous. Identify the maxillary palpi which project from it. Are they segmented? Note a small sclerite at the base of each palpus - this is the maxillary plate, all that remains of the body of the maxilla. The rostrum is a composite structure made up of part of the clypeal region of the head, and also includes the maxillae and probably the basal part of the labium. The mandibles are entirely absent. 3. Examine the haustellum. The body of the haustellum as well as the entire oral disc is derived from the labium. The anterior face of the haustellum is deeply channeled to form a labial groove. Forming the edge of this groove are two stylets. The upper one is the labrumepipharynx, whose epipharyngeal face is deeply channeled. The lower is the hypopharynx and contains the salivary canal. The two, when opposed form the food canal. 4. Examine the oral disc. Note that it is composed to two lateral lobes, the labellar lobes, united posteriorly. The labellar lobes are soft spongy structure that are supported by two sclerites. Examine the labellar lobes. Note that their upper surfaces are provided with bristles while the underside is transversed by series of transverse channels, the pseudotracheae. Note the pseudotracheae running from blind lateral ends toward larger anterior and posterior collecting channels along their mesal margins. These converge on each side near the middle. At this point the liquid collected through the pseudotracheae is transferred to the food canal between the labrum-epipharynx and hypopharynx. INTERNAL MORPHOLOGY OF THE ORTHOPTEROID Obtain and anesthetize an orthopteroid. With your scissors remove the legs and wings (if present). Pin ventral side down in the dissecting pan by pushing pins through the pronotum. Position under your microscope and examine the mid-dorsal line. You should be able to see the dorsal vessel or “heart” beneath the exoskeleton. It functions in a peristaltic wave from posterior to anterior. It is very fine and is usually destroyed upon dissection. How often does a given chamber “beat” per minute? Next, gently push the point of a pair of fine scissors beneath the posterior edge of the last abdominal tergum and cut the tergum forwards to the neck. Cut carefully so as to avoid damaging anything beneath the cuticle. With pins move the cuticle aside and hold in place by pushing the pin into the wax. What you see now will depend upon which sex you have. If you have a female that is gravid you may see little else except eggs. The ovaries are paired, with each ovary made up of many small egg tubes. Carefully tease out one of the ovaries without breaking the posterior connection and see if you can recognize the anterior germarium and the growth stages leading up to the mature eggs that are held in the calyx of the lateral oviducts. The lateral oviducts join the common or median oviduct at the midline. Connected to the median oviduct is the sperm storage organ known as the spermatheca, and the accessory gland. The latter is often larger in some species and is used for egg cement and/or egg cases. Upon opening a male specimen you will reveal the testes and the digestive tract. The testes are also paired organs which in crickets are very large, roughly triangular in shape and covered on the outside with many flat tracheal trunks. A testis is made up of a series of small tubes not dissimilar to an ovary. Each tube meets in the center of the testis at the vas deferens a tube which leads to the vas deferens. The vas deferens leads to the seminal vesicle and each seminal vesicle enters the ejaculatory duct. From the ejaculatory duct arises the accessory gland. This is a large circular organ with hundreds of small tubes on the outside. The function of the gland is production of the seminal fluid. The tracheae are the silvery white tubes that branch many times. The larger tubes are quite conspicuous. Take one of these larger tubes and pull on it with your forceps. Notice how it unravels. It appears to be made in a series of coils. These coils are thickened parts known as taenidia. The digestive system is a long, sometimes convoluted tube running through the mid-part of the insect. With your forceps, reach as far forward as possible in the thorax and gently pull the esophagus loose. Then by lifting and removing attached tracheae you should be able to stretch out the tract. Leave it attached at the posterior end. The esophagus leads to the crop, a bulbous storage organ. Behind the crop is the proventriculus, commonly termed the gizzard in the Orthoptera, since it contains a set of grinding teeth. Cut the proventriculus open. Also open outgrowths of the mid-gut known as gastric caecae (sing. caeca). They are usually found in multiples of two. Behind the mid-gut is the proctodeal valve which separates the mid-gut from the hind-gut. Near the junction of the mid - and hind-gut the Malpighian tubules attach to the gut. Be careful to trace out their actual junction because they may be attached to the gut in various places by tracheae. The tubules branch many times in the Orthoptera after they leave the hind-gut. The hind-gut in some insects may be divided into several regions, but terminates into the bulbous rectum which occurs just anterior to the anus. In the mid-ventral line lies the nerve cord. Start near the rectum of the digestive tract and work forward. The first ganglion that you should see is the largest abdominal ganglion, the most posterior. It is actually a fusion of ganglia of the last few segments. You should be able to trace it forward via the paired connectives. If you wish you can trace the nerve cord into the head of a new specimen and examine the structures there. You will need help if you decide to do this, so ask your instructor.