Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
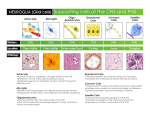
Markers of Central Nervous System Glia and Neurons In Vivo During Normal and Pathological Conditions J.M. REDWINE and C.F. EVANS 1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 119 2 Limits of Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 120 3 3.1 3.2 3.3 3.4 3.5 3.6 Constitutively Expressed Markers . Mature Oligodendrocytes . . . . . . Adult Oligodendrocyte Progenitors . Astrocytes . . . . . . . . . . . . . . . Microglia . . . . . . . . . . . . . . . Other Glial Cells . . . . . . . . . . . Neurons . . . . . . . . . . . . . . . . 4 Markers Induced During Pathological Conditions . . . . . . . . . . . . . . . . . . . . . . . . . 130 5 Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 133 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 121 121 123 125 127 128 129 References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 134 1 Introduction Cell markers are valuable tools for examining the function of cells in normal conditions as well as during disease and repair processes. In fact, our understanding of the cell types that make up the central nervous system (CNS) is very much shaped by the markers available to identify them. CNS cell types were originally identi®ed by morphology. The discovery of various proteins speci®c to certain cells led to the production of cell-type-speci®c antibodies that have been used to identify cells in situ. An ideal marker is speci®c to a given cell type in normal conditions and/or during conditions involving injury or disease. As simple as these criteria sound, they are not easy to ful®ll. Markers can be expressed on more that one cell type. Astrocytes and olfactory-tract-ensheathing glia both express glial ®brillary acidic protein (GFAP), even though they have clear phenotypic, anatomical, and functional dierences (RAMON-CUETO and VALVERDE 1995). Also, a marker that is speci®c for a given cell type in normal conditions Dept. of Neuropharmacology, The Scripps Research Institute, 10550 North Torrey Pines Road, La Jolla, CA 92037, USA Present addresses: C.F. Evans, Digital Genes Technologies Inc., 11149 N. Torrey Pines Rd. #110, La Jolla, CA 92037, USA J.M. Redwine, Neurome, 11149 N. Torrey Pines Rd, La Jolla, CA 92037, USA 120 J.M. Redwine and C.F. Evans can be induced or up-regulated on other cell types during conditions such as in¯ammation, disease, or injury. Therefore cell type markers alone do not always conclusively identify a cell type. Antibodies to cell-type-speci®c markers identify a cell at the molecular level and are considered more reliable than identi®cation of cell type by morphology. Therefore, if a marker for a cell type is not available, it is possible that the existence of a particular cell type may be overlooked. An example of this in the CNS was the characterization of an oligodendrocyte progenitor cell type in the adult rodent and human CNS that was only recently identi®ed (FRENCH-CONSTANT and RAFF 1986; LEVINE and CARD 1987). Adult oligodendrocyte progenitors are scattered throughout the parenchyma of the brain and spinal cord, but were completely overlooked prior to the availability of in vivo markers speci®c for these cells. A more accurate understanding of CNS cell types and their distinct functions may lead to a better understanding of disease pathology and development of novel therapeutic agents for CNS diseases. The current division of cell types within the CNS into four major classes (neurons, astrocytes, oligodendrocytes, and microglia) is likely an oversimpli®cation. This article is written with the anticipation that additional cell types distinct in developmental origin, function, and expression pattern of antigenic markers will be characterized in the future. Markers that exist for cell types today should be looked at as work in progress. Some markers used presently may eventually be shown not to be consistent with a speci®c cell phenotype, and it is very likely that novel markers will be available in the future. Novel markers may identify known cell types more clearly, or may lead to the discovery of entirely ``new'' cell types. Identi®cation of new antigenic markers will broaden our understanding of the true complexity of the CNS. 2 Limits of Discussion The ®rst section of this chapter will review constitutively expressed markers of glia and neurons in the CNS, and the second section will discuss markers during in¯ammatory or pathological conditions. This review will focus on commonly used markers for glial cell populations and pan-neuronal markers in adult rodents and humans. Neuronal markers that label sub-populations, such as neurotransmitters, neurotransmitter-synthesizing enzymes, neurotransmitter receptors, and neuropeptides, are beyond the scope of this chapter. CNS cell type markers used in vitro or during embryonic or neonatal development are often dierent than markers speci®c for cell types in vivo and in adults. Since this volume focuses on in¯ammation and pathology in the adult CNS, in vitro and developmental marker expression patterns will not be addressed. In addition, discussion will be limited to markers expressed at the protein level that are not secreted molecules. Markers of Central Nervous System Glia and Neurons 121 3 Constitutively Expressed Markers 3.1 Mature Oligodendrocytes Oligodendrocyte precursors originate from germinal regions located in subventricular zones in the brain (REYNOLDS and WILKIN 1988; LEVISON and GOLDMAN 1993) and germinal zones in the spinal cord (PRINGLE and RICHARDSON 1993). Neonatal oligodendrocyte precursors are small with few processes in vivo. After proliferating in germinal zones and migrating to the parenchyma, oligodendrocytes stop dividing, extend processes, and begin synthesizing myelin sheaths. In vivo, mature oligodendrocytes have a round cell body and long, thin processes (Fig. 1A). Oligodendrocytes are perhaps the most specialized macroglial cell type. The primary function of oligodendrocytes in normal conditions is to myelinate axons. Oligodendrocytes begin myelinating axons postnatally by extending modi®ed processes that wrap around nearby axons and eventually form a myelin sheath. A single process from an oligodendrocyte forms a single segment of myelin around an axon that consists of layers, or lamellae. The myelin sheath is a modi®ed membrane containing lipids, glycolipids, gangliosides, and myelin-speci®c proteins. Myelin sheaths and oligodendrocytes are often identi®ed by immunolabeling myelin- Fig. 1A±D. Examples of markers and morphologies for oligodendrocytes, neurons, and astrocytes in mouse spinal cord are shown. A The dorsal root entry zone of a mouse spinal cord stained with CNP (Sigma, St. Louis, Mo., clone 11-5-B). The arrows indicate small round oligodendrocyte cell bodies. Thin processes are also visible. B Mouse spinal cord neuron cell bodies stained with NSE (Polysciences, Warrington, Pa.). C,D Astrocytes in mouse spinal cord stained with GFAP (DAKO, Carpinteria, Calif.). C Radial astrocyte processes seen in white matter. The punctate pattern is due to labeling of crosssections of astrocyte processes. D Another astrocyte morphology, the stellate morphology, seen in gray matter. All stains are indirect immuno¯uorescence imaged with a laser scanning confocal microscope. A±C Single planes, D 3-dimensional reconstruction of 45 planes 0.2lm apart 122 J.M. Redwine and C.F. Evans speci®c proteins. Myelin-speci®c proteins have various functions, which include interacting with the axonal membrane and promoting orderly compaction of the many lamellae of the myelin sheath. The action potential in a myelinated axon is propagated via saltatory conduction at the spaces where ion channels are concentrated, which are referred to as the nodes of Ranvier. The myelin sheath: (1) helps structure saltatory conduction, and (2) increases the resistance and reduces the capacitance of the axonal membrane. Both of these functions greatly increase the speed and eciency of the action potential. Therefore, the presence of myelin sheaths is crucial to the normal functioning of the CNS, and the ability of an axon to maintain conduction is impaired if the myelin sheath is lost at any point along the axon, as is observed in the human disease multiple sclerosis. Myelin proteins synthesized by oligodendrocytes in the CNS are the most well characterized and most widely used markers for the oligodendrocyte cell type. Antibodies to myelin proteins include proteolipid protein (PLP; SOBEL et al. 1994; GREER et al. 1996), myelin basic protein (MBP; STERNBERGER et al. 1978; SHINE et al. 1992), 20 ,30 -cyclic nucleotide-30 -phosphodiesterase (CNP; SHEEDLO and SPRINKLE 1983; REISER et al. 1994), myelin oligodendrocyte glycoprotein (MOG; BIRLING et al. 1993; GARDINIER and MATTHIEU 1993), myelin-associated glycoprotein (MAG; STERNBERGER et al. 1979; MCGARRY et al. 1983; DOBERSEN et al. 1985; TRAPP et al. 1989) and myelin-associated oligodendrocytic basic protein (MOBP; HOLZ et al. 1996). The most common antigens used to identify oligodendrocytes are PLP, MBP, and CNP. PLP and MBP are the most abundant myelin proteins (50% and 30%, respectively), and antibodies to these proteins label oligodendrocyte cell bodies, processes, and the compact myelin sheath. CNP is less abundant than PLP or MBP, and it is not found in the compact myelin sheath. Antibodies against CNP are useful for identifying oligodendrocytes in vivo since they label oligodendrocyte cell bodies and processes, as well as the outer myelin membrane, but not inner compact myelin lamellae. Anti-MOG antibody labels oligodendrocytes and myelin sheaths (Birling et al. 1993), anti-MAG antibodies label the periaxonal myelin membrane closely associated with the axon (TRAPP et al. 1989), and anti-MOBP antibody primarily labels the myelin sheath (HOLZ and SCHWAB 1997). A monoclonal antibody named Rip (FRIEDMAN et al. 1989) has also been used in vivo to identify rat oligodendrocytes (FRIEDMAN et al. 1989; LEVINE et al. 1993; REYNOLDS and Hardy 1997). The Rip antibody stains the oligodendrocyte cell body, processes, and myelin and the staining pattern resembles that by antibodies to CNP. A more recently identi®ed antibody that recognizes oligodendrocytes is monoclonal antibody CC-1 (SMITH et al. 1993). CC-1 recognizes the N-terminal of a protein product of the APC (adenomatous polyposis coli) gene originally isolated from human colorectal tumor cell lines (SMITH et al. 1993). Mutations in the APC gene are associated with a syndrome called familial adenomatous polyposis that predisposes patients to colon cancer, and in rarer cases brain cancer (POWELL et al. 1992; LASSER et al. 1994). In rodent tissue, CC-1 clearly labels the oligodendrocyte cell body, but not processes or myelin sheaths (BHAT Markers of Central Nervous System Glia and Neurons 123 et al. 1996). The speci®city of CC-1 for oligodendrocytes has been demonstrated in rat optic nerve (B.A. Barres, personal communication), mouse brain (BHAT et al. 1996; FUSS et al. 2000), and mouse spinal cord (FUSS et al. 2000; MESSERSMITH et al. 2000), so CC-1 is a useful oligodendrocyte marker. While CC-1 labels oligodendrocytes, Bhat and colleagues found that CC-1 also labels cerebellar Purkinje neurons (BHAT et al. 1996). Because CC-1 is relatively new, colocalization studies with various cell type markers should continue in dierent models and tissues to further investigate oligodendrocyte speci®city. The ability of this antibody to label the oligodendrocyte cell body without labeling processes (in contrast to most other markers) is very advantageous for the study of oligodendrocytes. The heavy labeling of myelin sheaths by myelin protein antibodies often prevents visualization of the oligodendrocyte cell body in white matter. By labeling the cell body exclusively, the density of oligodendrocytes present or the extent of oligodendrocyte loss can be more clearly observed. Future experiments will hopefully continue to provide evidence for the speci®city of CC-1 as an oligodendrocyte marker in rodents, and possibly in humans. An additional oligodendrocyte marker less often used is transferrin (CONNER and FINE 1986). Transferrin is primarily expressed by oligodendrocytes, but can be detected in ependymal cells in some periventricular regions and in the choroid plexus of the rat CNS (BENKOVIC and CONNOR 1993). Anti-galactocerebroside (GC) antibodies (SOMMER and SCHACHNER 1981) have been extensively used in vitro to speci®cally label mature oligodendrocytes. Although there are some reports of anti-GC staining in situ (REYNOLDS and HARDY 1997), immunohistochemical staining in tissue is often technically more dicult compared to myelin protein antibodies. 3.2 Adult Oligodendrocyte Progenitors An example of our reliance on markers to identify cell types was made evident by the relatively recent discovery and characterization of adult oligodendrocyte progenitors that occurred only after speci®c markers were available. This cell type is worth noting since it may play a role in in¯ammatory and repair processes in the CNS. Initial immunohistochemistry experiments on rat brain using antibodies generated against a chondroitin sulfate proteoglycan (NG2) labeled cells that did not clearly ®t into a known class of cell types (STALLCUP 1981; WILSON et al. 1981). The cells were tentatively identi®ed as interneurons in the cortex (STALLCUP et al. 1983), or ``smooth protoplasmic astrocytes'' (astrocytes with low levels of ®laments) in the cerebellum (LEVINE and CARD 1987). The data regarding NG2 labeling in these initial reports have proved to be reliable and have been con®rmed by independent laboratories (NISHIYAMA et al. 1997, 1999), but the cell type classi®cation has changed. At the time of the initial reports, the existence of an adult oligodendrocyte progenitor was not characterized or expected, especially at the high density seen throughout the CNS by immunohistochemistry using the NG2 antibody. Adult oligodendrocyte progenitors were just beginning to be identi®ed in 124 J.M. Redwine and C.F. Evans culture experiments (FRENCH-CONSTANT and RAFF 1986; WOLSWIJK and NOBLE 1989), and therefore almost nothing was known about them. NG2-expressing cells in the adult CNS were identi®ed as adult oligodendrocyte progenitors in vivo only after: (1) immunohistochemical studies using NG2 antibody were performed on tissues at dierent stages of development (STALLCUP and BEASLEY 1987; LEVINE et al. 1993), (2) culture experiments were performed that followed dierentiation of NG2+ cells into mature oligodendrocytes in vitro (LEVI et al. 1987; LEVINE and STALLCUP 1987; LEVINE et al. 1993), and (3) a second in vivo oligodendrocyte progenitor marker, platelet-derived-growth-factor-a receptor (PDGFaR), was identi®ed (HART et al. 1989; PRINGLE et al. 1992; OUMESMAR et al. 1997). Eventually, adult oligodendrocyte progenitors (labeled with NG2 and/or PDGFaR) were found to be abundantly present in white and gray matter of adult rats and mice (LEVINE et al. 1993; NISHIYAMA et al. 1997, 1999; REYNOLDS and Hardy 1997), and in normal and demyelinated human brain (GOGATE et al. 1994; SCOLDING et al. 1998; CHANG et al. 2000). The function of adult oligodendrocyte progenitors in normal conditions is not well understood. They proliferate to a small extent (LEVINE et al. 1993) and thus may replace mature oligodendrocytes as needed. A striking observation was the abundance of these cells in the CNS. They are spread throughout the gray and white matter of the brain and spinal cord and are as numerous as other glial cells such as microglia (DAWSON et al. 2000), so they should perhaps be considered as a fourth major class of glial cells. Adult oligodendrocyte progenitors are found in increased numbers at sites of injury, in¯ammation, demyelination, and remyelination (LEVINE 1994; NISHIYAMA et al. 1997; KEIRSTEAD et al. 1998; REDWINE and ARMSTRONG 1998). In rat experimental autoimmune encephalomyelitis (EAE), NG2+ adult oligodendrocyte progenitors are often seen closely associated with activated microglia/macrophages and lymphocytes (NISHIYAMA et al. 1997). The ability of adult oligodendrocyte progenitors to react to dierent types of CNS damage and their abundance throughout the CNS may signify that they have multiple functions (LEVINE et al. 2000). In normal conditions, adult oligodendrocyte progenitors can be identi®ed by antibodies against NG2 (WILSON et al. 1980; STALLCUP 1981; LEVINE and CARD 1987; NISHIYAMA et al. 1999; DAWSON et al. 2000), PDGFaR (HERMANSON et al. 1992; NISHIYAMA et al. 1996), and 04 (SOMMER and SCHACHNER 1981; REYNOLDS and HARDY 1997). The anti-NG2 antibody labels adult oligodendrocyte progenitors in normal adult rodent brain and spinal cord (WILSON et al. 1980; STALLCUP 1981; LEVINE and CARD 1987; NISHIYAMA et al. 1999; DAWSON et al. 2000). NG2 is the most reliable marker for this cell type in rat and mouse since it has been found to speci®cally label adult oligodendrocyte progenitors in normal and reactive or in¯ammatory conditions (NISHIYAMA et al. 1997; KEIRSTEAD et al. 1998; REDWINE and ARMSTRONG 1998). Human anti-NG2 antibodies are available and have been used to identify oligodendrocyte progenitors in adult human tissue (CHANG et al. 2000), but the speci®city of NG2 to this cell type in human tissue needs further characterization, since NG2 immunoreactivity was reported on cultured human microglia (POULY et al. 1999). Anti-PDGFaR antibody (HERMANSON et al. 1992) Markers of Central Nervous System Glia and Neurons 125 labels adult oligodendrocyte progenitors in normal rodent and human tissue (NISHIYAMA et al. 1996, 1999), but during reactive conditions such as virus-induced demyelination PDGFaR has also been identi®ed in astrocytes (REDWINE and ARMSTRONG 1998). Therefore, if PDGFaR is used as a marker for oligodendrocyte progenitors in reactive conditions, additional criteria must be used, such as cell morphology. Reactive astrocytes that express PDGFaR have larger cell bodies, larger nuclei, and fatter processes than the respective features seen on the adult oligodendrocyte progenitor (REDWINE and ARMSTRONG 1998). Antibodies to O4 (SOMMER and SCHACHNER 1981) label adult (and neonatal) oligodendrocyte progenitors (REYNOLDS and HARDY 1997). However this antibody must be used with a second oligodendrocyte marker antibody to con®rm progenitor phenotype. Adult oligodendrocyte progenitors express O4 but not mature oligodendrocyte markers, whereas mature oligodendrocytes express O4 and mature oligodendrocyte markers (REYNOLDS and HARDY 1997). An additional marker used to identify adult oligodendrocyte progenitors in rats is GD3 ganglioside. However, there is recent evidence that GD3 is not cell-type-speci®c (GOLDMAN and REYNOLDS 1996). 3.3 Astrocytes Astrocytes have a similar temporal and spatial pattern of origin as oligodendrocytes. There is evidence that astrocytes and oligodendrocytes share a common progenitor from the subventricular zone that migrates into the parenchyma postnatally (LEVISON and GOLDMAN 1993); however, other reports have not supported this observation (LUSKIN et al. 1993; LUSKIN and MCDERMOTT 1994). Radial glia present during development are also a source of astrocytes. Radial glia guide embryonic neuronal migration, express vimentin but not GFAP, and later dierentiate into GFAP+ astrocytes (CHANAS-SACRE et al. 2000). Once astrocytes migrate from germinal zones and dierentiate, they have more than one morphology. A radial or ®brous morphology and a stellate, or star-shaped morphology (also called ``protoplasmic astrocytes'') are observed in normal CNS (Fig. 1C,D). Fibrous astrocyte morphology is typically seen in white matter with radial processes that often parallel axons. Stellate astrocytes are more often found in gray matter and have less ®lament structure in their processes compared to ®brous astrocytes (PETERS et al. 1991). Astrocytes are likely to be more diverse than is currently known (MILLER et al. 1994) and to have diverse functions during normal and pathological conditions. In normal conditions, astrocyte processes are typically found in the perivascular region. Perivascular astrocytic end-feet form a glial-limiting membrane that is separated from endothelial cell walls by a basal lamina. These astrocyte processes have been proposed to help maintain or modulate the blood-brain barrier, but the exact functional relation to the blood-brain barrier is not well understood (PETERS et al. 1991). Astrocyte processes in perisynaptic regions take up excitatory neurotransmitter glutamate from the synapse and recycle it to its precursor glutamine (WESTERGAARD et al. 1995; BEZZI et al. 1999), thus protecting neurons from excitotoxicity. Astrocyte processes are also observed closely 126 J.M. Redwine and C.F. Evans associated with the axolema at the nodes of Ranvier (PETERS et al. 1991) and may be involved in regulating ion concentrations (BLACK and WAXMAN 1988). In addition, an increased number of astrocyte processes in perisynaptic regions of rat dentate gyrus following long-term potentiation stimulation indicates a possible role of astrocytes in maintaining or modulating changes in synaptic function (WENZEL et al. 1991). Astrocytes are involved in local in¯ammatory responses in many disease models. Reactive astrocytes often seen within and near regions of in¯ammation and tissue damage have shorter, thicker processes and a larger cell body and nucleus than the resting astrocyte. Astrocytes proliferate in response to viral infection, demyelination, and injury (MIYAKE et al. 1992; MORRIS et al. 1994; REDWINE and ARMSTRONG 1998; NORTON 1999). Following demyelination induced by viral infection, astrocytes synthesize growth factors thought to be involved in myelin repair (REDWINE and ARMSTRONG 1998; MESSERSMITH et al. 2000). Astrocytes have been reported to phagocytose debris in certain conditions (AL-ALI and AL-HUSSAIN 1996; BECHMANN and NITSCH 1997), and they may also be involved in promoting blood coagulation (EDDLESTON et al. 1993). They produce in¯ammatory cytokines and chemokines involved in activating and maintaining a local immune response (discussed elsewhere in this volume). The most well characterized speci®c marker for the astrocyte cell type is GFAP (BIGNAMI et al. 1972). Many monoclonal and polyclonal antibodies to GFAP are available, and most work well. Antibodies to GFAP recognize astrocytes throughout the brain and spinal cord and label astrocytes with either radial or stellate morphology (Fig. 1C,D). S100, a calcium-binding protein, is also expressed primarily in astrocytes (LUDWIN et al. 1976; HAAN et al. 1982); however, S100 immunoreactivity can be detected in some neuronal populations in rat brain (RICKMANN and WOLFF 1995; YAMASHITA et al. 1999). S100 is a secreted protein that can increase neurite outgrowth and is considered a neurotropic cytokine. Since S100 is secreted it may not serve as the best astrocyte-speci®c marker. Finally, tissue factor is speci®cally expressed by astrocytes at the mRNA level (EDDLESTON et al. 1993), but tissue factor protein as a marker for astrocytes has not been well characterized. Thus, the only marker that is well characterized as a speci®c marker for astrocytes is GFAP. As discussed above, astrocytes can have dierent morphologies and multiple functions. The possibility that astrocytes are a heterogeneous cell population has been discussed in reviews (MILLER et al. 1994) and is perhaps best stated in Peters et al., ``Astrocytes... represent a family of cells that have some shared properties and other distinctive morphological and functional characteristics that depend on their location in the nervous system.'' (PETERS et al. 1991, p 284). It is likely that cells labeled with GFAP actually represent more than one functionally distinct cell type. For example, a GFAP+ cell producing in¯ammatory cytokines in response to a viral infection may not be capable of glutamate uptake. There is clearly a need for more astrocyte markers that identify functionally distinct populations of cells. A more accurate understanding of astrocyte cell types will be important for understanding pathology underlying disease. Markers of Central Nervous System Glia and Neurons 127 3.4 Microglia The developmental origin of microglia is still somewhat controversial (LING and WONG 1993). There is evidence that microglia originate from bone marrow (HICKEY et al. 1992; KRIVIT et al. 1995; EGLITIS and MEZEY 1997) and from progenitor cells originating from neuroectoderm (RICHARDSON et al. 1993; FEDOROFF et al. 1997). Microglia share many functions and antigenic markers with peripheral macrophages (discussed below) and are considered to be the resident macrophages of the CNS. Resting microglia have elaborate rami®ed processes (Fig. 2A), while activated microglia have shorter, fatter processes (Fig. 2B) and can even resemble macrophages. Resting microglia are potential immunoeector cells within the CNS (STREIT et al. 1988). Microglia may promote neuronal survival during development (NAGATA et al. 1993; ELKABES et al. 1996), and in the adult CNS, it has been assumed that their primary function is immune surveillance due to their expression of macrophage markers. However, it is possible that microglia have other functions. In pathological conditions, they are capable of locally modulating in¯ammatory responses by expressing cytokines, chemokines, and adhesion molecules (GEHRMANN et al. 1995; BAR-OR et al. 1999; LEE and BENVENISTE 1999; SORENSEN et al. 1999). They are capable of phagocytosing debris or damaged cells (STREIT and KREITZBERG 1988). In addition, microglia have been reported to express costimulatory molecules (DE SIMONE et al. 1995; DANGOND et al. 1997) and to present antigen to T cells (MATSUMOTO et al. 1992; KATZ-LEVY et al. 1999). Their ability to stimulate T cell proliferation is reduced compared to peripheral antigen-presenting cells (CARSON et al. 1998, 1999), but microglia are able to induce T cell in¯ammatory cytokine production (CARSON et al. 1999). Commonly used markers for microglia (and macrophages) are Mac-1 (complement receptor type 3, CR3), F4/80, and lectin binding. Mac-1 binds complement fragment C3bi and intercellular adhesion molecule-1 (ICAM-1), and is used as a receptor for some strains of gram-negative bacteria (HORWITZ 1992). Mac-1 Fig. 2A,B. Resting and activated microglia express F4/80. A monoclonal antibody to F4/80 (Serotec, Raleigh, N.C.) was used to identify a resting microglial cell (arrow, A) in which F4/80 is expressed at a low level, and an activated microglial cell (arrow, B) de®ned by high levels of immunoreactive F4/80. Both panels are taken from a transgenic mouse that had focal areas of chronic CNS immune in®ltration following infection with lymphocytic choriomeningitis virus (EVANS et al. 1996). The chromagen used in the immunostaining procedure was diaminobenzidine, which gives a brown product where primary antibody bound. Original magni®cation 600´ 128 J.M. Redwine and C.F. Evans consists of an a-chain, CD11b, and a b-chain, CD18. Anti-Mac-1 and anti-CD11b antibodies are available for mouse (Ho 1982) and human (BELLER et al. 1982) tissues. In the rat, a commonly used monoclonal antibody that binds Mac-1 is MRC OX-42 (ROBINSON et al. 1986). Another microglia/macrophage marker, F4/ 80, is a cell surface glycoprotein expressed by microglia and macrophages in mouse and rat tissues (AUSTYN and GORDON 1981; STARKEY et al. 1987; Fig. 2A). F4/80 has recently been implicated in cell-to-cell signaling pathways (WARSCHKAU and KIDERLEN 1999). Several lectins that bind to microglia (and macrophages) in mouse, rat, and human tissues have been used to identify these cells (MANOJI et al. 1986; STREIT and KREUTZBERG 1987; SUZUKI et al. 1988; HAUKE and KORR 1993; ACARIN et al. 1994; SCHUMACHER et al. 1994). Monoclonal antibody ED1, which binds a cytoplasmic antigen in rat microglia and macrophages, has been used as a microglial marker (DIJKSTRA et al. 1985). In humans, CD68 and Ham-56 are also used as microglial markers (ADAMS and POSTON 1990; HULETTE et al. 1992; FOX et al. 1996; ANDJELKOVIC et al. 1998). The cell type speci®city of markers can change during in¯ammation, which adds to the diculty of distinguishing activated microglia from other cell types. For example, CD11b, which is speci®c for microglia and macrophages in normal conditions, can be expressed by activated CD8+ T cells (MCFARLAND et al. 1992). As mentioned above, currently available markers used to identify microglia are also expressed by macrophages. During injury, viral infection, in¯ammation, or demyelination, microglia can become activated and macrophages can in®ltrate the CNS parenchyma. Therefore, in these conditions it becomes very dicult to distinguish between either cell type in situ by morphology or with immunohistochemical methods. In human tissue, some markers have been found on monocytes that are not expressed by microglia. Antibodies to L1 (clone MAC 387) and lysozyme bind to monocytes in culture but do not bind to microglia in situ or in vitro in the presence of interferon-c (ULVESTAD et al. 1994). Microglia and macrophages isolated from the rodent CNS can be distinguished using double labeling with antibodies to CD45 and CD11b and FACS analysis. CD45 high intensity/ CD11b identi®es macrophages, and CD45 low intensity/CD11b identi®es microglia (FORD et al. 1995; CARSON et al. 1999). It would be very useful to be able to easily distinguish between microglia and macrophages in situ, and hopefully markers for these speci®c cell types will be identi®ed in the near future. 3.5 Other Glial Cells Other minor glial cell populations are present in the adult CNS. These have been de®ned based on morphology, function, and anatomical location, and few distinct cell markers exist to identify them. Perivascular microglia are bone-marrow-derived and are thought to play a crucial role in the initiation of immune cell in®ltration and antigen presentation (HICKEY and KIMURA 1988). In rats, they express microglia markers and at least one additional marker, ED2 (DIJKSTRA et al. 1985; GRAEBER et al. 1989), which distinguishes perivascular microglia from parenchymal Markers of Central Nervous System Glia and Neurons 129 microglia. Perivascular microglia may have distinct functions that are important in modulating in®ltration and in¯ammation in the CNS. Future characterization of additional speci®c markers will help increase our understanding of these cells. Bergmann glia processes are found in the molecular layer of the adult cerebellum and are often closely associated with dendritic trees and cell bodies of Purkinje neurons. They express GFAP and vimentin (SHAW et al. 1981) and are involved in the reuptake of neurotransmitters (MULLER and KETTENMANN 1995). Ensheathing glia are specialized cells in the olfactory tract that have functions similar to myelinating cells and astrocytes. They ensheath olfactory neurons and form a glial limiting membrane. They express vimentin, GFAP, and adhesion molecules such as L1, laminin, and neural-cell adhesion molecule (RAMON-CUETO and VALVERDE 1995; BARNETT and FRANCESCHINI 1999). 3.6 Neurons Neuronal stem cells proliferate in germinal zones such as the ventricular zone in the brain, or ventricular and intermediate zones in the spinal cord, and migrate along processes of radial glia out to parenchymal regions where they dierentiate. Many types of neurons exist and can be de®ned by patterns of neurotransmitter and/or neuropeptide expression and various morphologies. Neuronal morphologies in the brain range from small interneurons with round cell bodies and few processes to pyramidal neurons (in the motor cortex, for example) that have larger cell bodies with processes, to cerebellar Purkinje neurons that have a large cell body and an extensive dendritic network. Neuronal morphologies in the spinal cord are less varied, but vary from small sensory neurons in laminae I and II, and small to intermediate-size interneurons, sensory or motor neurons in laminae I±VIII, to large ventral a-motor neurons in the ventral horn in lamina IX. A broad, basic de®nition of the function of a neuron is to receive and transmit electrochemical signals through synaptic connections. Although this de®nition includes most neurons (with some exceptions including pseudo-unipolar sensory neurons, neuroendocrine neurons, and chemoreceptor neurons in the brainstem and hypothalamus), the chemical signals that neurons use vary greatly. This review is strictly concerned with how to identify a neuron in any anatomical location and how to distinguish it from other cell types. Neuronal cell bodies are best identi®ed with neuron-speci®c enolase (NSE). In the normal CNS, NSE is speci®cally expressed in most neurons, and not other cell types (MARANGOS et al. 1978, 1979; SCHMECHEL et al. 1978; FORSS-PETTER et al. 1986). NSE immunoreactivity appears primarily in the cytoplasm, with little immunoreactivity in dendritic or axonal processes (Fig. 1B). A second neuron-speci®c marker is neuron-speci®c nuclear protein, neuN (MULLEN et al. 1992), which is expressed in most neuronal cells in rodent and human CNS (MULLEN et al. 1992; HARVEY et al. 1997; TODD et al. 1998). Other pan-neuronal markers are found primarily in processes and include microtubule associated protein-2 (MAP2; HUBER and MATUS 1984; TUCKER et al. 1988), neuron-speci®c b-tubulin (monoclonal antibody Tuj1; LEWIS et al. 1985), 130 J.M. Redwine and C.F. Evans neuro®lament (SHAW et al. 1981), and tau protein (MERCKEN et al. 1992; ZHENGFISCHHOFER et al. 1998). Antibodies to these markers do not always label neurons clearly in situ due to the ®lamentous pattern of the label. The presence of MAP-2 antigen in dendritic processes can obscure visualization of neuronal cell bodies. In addition, neuro®lament and tau are located primarily in axons, and thus antibodies to these proteins eciently label neuronal structures, but not the neuronal cell body. Finally, neuronal markers that label the peri-synaptic region include growthassociated protein-43 (GAP-43) (MERCKEN et al. 1992) and synaptophysin (WIEDENMANN and FRANKE 1985; SILLEVIS SMITT et al. 1993; STRIDSBERG et al. 1994). GAP-43 and synaptophysin in most cases do not clearly label neuronal cell bodies in situ. However, synaptophysin does label the cell body of some discreet populations of neurons (GOTO et al. 1993). 4 Markers Induced During Pathological Conditions The expression pattern of many antigenic markers changes during pathological conditions, often in response to cytokines or chemokines. The level of expression of a marker can change, and the cell types that express a marker can also change. Many molecules induced during pathological conditions are expressed by more than one kind of cell, making it dicult to use them as true inducible markers to detect speci®c cell types. Also, when in¯ammatory cells from the periphery in®ltrate the CNS, it often becomes more dicult to identify speci®c cells since some immune cells and CNS resident cells can express the same protein (e.g., microglia and macrophages). NG2 immunoreactivity is increased within and near regions of injury and in¯ammation such as demyelination (KEIRSTEAD et al. 1998; REDWINE and ARMSTRONG 1998) and stab wounds (LEVINE 1994). Although NG2 expression is up-regulated, it remains speci®c for adult oligodendrocyte progenitors and does not colocalize with the astrocyte marker GFAP, or microglial markers OX-42 (LEVINE 1994), F4/80 or CD45 (NISHIYAMA et al. 1997). This reactive response of progenitors may promote tissue repair. In contrast to NG2, PDGFaR does not remain cell-type-speci®c in pathological conditions. In normal conditions the oligodendrocyte progenitor is the only adult CNS cell type that constitutively expresses detectable levels of PDGFaR (discussed above). However during murine-hepatitisvirus-induced demyelination, astrocytes also express the PDGFaR (REDWINE and ARMSTRONG 1998). During pathological conditions such as demyelination, viral infection, or injury, there is often an increase in GFAP immunoreactivity (VIJAYAN et al. 1990; SUN et al. 1995; REDWINE and ARMSTRONG 1998). This is due to increases in the intensity of GFAP immunoreactivity within reactive astrocytes and an increase in the number of GFAP+ astrocytes present as a result of proliferation (VIJAYAN et al. 1990; MIYAKE et al. 1992). This astrocyte reactive response may be triggered Markers of Central Nervous System Glia and Neurons 131 by factors such as nucleotides released following injury (FRANKE et al. 1999). In¯ammatory cytokines such as interferon-c may also induce astrocytosis (YONG et al. 1991), but may inhibit astrocytosis in some cases (PAWLINSKI and JANECZKO 1997). Microglial/macrophage markers CD11b and F4/80 are up-regulated during many in¯ammatory responses. For example, in a model of virus-induced chronic CNS in¯ammation, F4/80 expression is enhanced on microglia located in white matter regions (Fig. 2). Other types of proteins induced during CNS in¯ammation include MHC molecules, cell adhesion molecules, and costimulatory molecules, as well as secreted molecules such as cytokines and chemokines. Most of these molecules have been detected on more than one type of cell, so they cannot be used as true cell type markers, yet they deserve mention since they are commonly induced in many dierent types of CNS pathology. MHC class I expression is induced in microglia/macrophages in many in¯ammatory conditions (Fig. 3A,B). In addition, we have detected oligodendrocytes expressing MHC class I in several dierent models of CNS in¯ammation. Although the MHC class I expression was not restricted to oligodendrocytes, this Fig. 3A±D. MHC expression is up-regulated in mouse spinal cord microglia during demyelination or in¯ammation. A,B The same ®eld double-labeled with A microglia marker CD11b (Pharmingen, San Diego, Calif., clone M1/70) and B MHC class I (Bachem, Torrance, Calif., clone ER-HR-52) during murine-hepatitis-virus-induced demyelination. Under normal conditions, MHC class I cannot be detected on microglia by immuno¯uorescence, and CD11b is very faint. However during demyelination, CD11b (A) is more intense, and nearly all microglia express MHC class I (B). The arrows in A and B indicate microglia expressing MHC class I. MHC class II can also be detected on microglia in this model (not shown). C,D The same ®eld double-labeled with C CD11b and MHC class II (D) during an in¯ammatory response in transgenic mice expressing lymphocytic choriomeningitis virus (LCMV) nucleoprotein and costimulatory molecule B7.1 in oligodendrocytes (Evans et al., in preparation). Following peripheral infection with LCMV, an extensive in¯ammatory response in the CNS is observed characterized by T cell in®ltration and MHC class I and class II expression. The arrow in C indicates a microglial/macrophage cell expressing MHC class II (arrow in D). Microglia/macrophages also express MHC class I in this model (not shown) 132 J.M. Redwine and C.F. Evans ®nding merits further discussion, since there are few reports of proteins induced in oligodendrocytes during in¯ammation. In transgenic mice expressing interferon-c in oligodendrocytes, demyelination occurred along with an in¯ammatory response characterized by MHC class I and class II expression (HORWITZ et al. 1997). Extensive MHC class I was observed in white matter and was shown to be colocalized with CNP (HORWITZ et al. 1999). White matter MHC class I expression was also observed during an in¯ammatory response in transgenic mice expressing a viral protein in oligodendrocytes (EVANS et al. 1996). Transgenic mice were designed which expressed the nucleoprotein (NP) of lymphocytic choriomeningitis virus (LCMV) under the control of a MBP promoter construct that targeted the NP transgene expression to oligodendrocytes. Epitopes of NP are known to be presented by MHC class I molecules to CD8+ T cells in the periphery (WHITTON et al. 1989). Following peripheral LCMV infection of these transgenic mice, a CNS in¯ammatory response was observed and persisted despite clearance of LCMV. During the in¯ammatory response, MHC class I immunoreactivity was observed on cells and along ®ne processes in white matter that resembled oligodendrocyte cell bodies and myelinated processes. In double transgenic mice expressing the LCMV-NP and the costimulatory molecule B7.1 in oligodendrocytes, a more extensive MHC class I white matter expression pattern was observed following LCMV infection (Evans et al., manuscript in preparation). Double immuno¯uorescence labeling with antibodies to CNP and MHC class I clearly identi®ed MHC class I on oligodendrocyte cell bodies and processes (Redwine et al., manuscript in preparation). Oligodendrocytes expressing MHC class I were also observed in a model of CNS in¯ammation induced by murine hepatitis virus infection (Redwine et al. 2001). The induction of MHC molecules on oligodendrocytes is likely due to in¯ammatory cytokines secreted by in®ltrating T cells (SUN et al. 1995). These studies provide evidence that, in addition to myelinating axons, oligodendrocytes may be capable of presenting antigen to CD8+ T cells via MHC class I, and thus may be targets of CD8+ cytotoxic T cells. MHC class II expression within the CNS is often up-regulated during in¯ammation or viral infection. MHC class II expression is usually limited to microglia/macrophages in the perivascular region as well as within the parenchyma (Fig. 3C,D; LAMPSON 1995; REDWINE et al. 2001), although it has also been detected on oligodendrocytes in two models (RODRIGUEZ et al. 1987; HORWITZ et al. 1999). Adhesion molecules such as ICAM are up-regulated in animal models of CNS in¯ammation and demyelination and have been reported to be expressed on astrocytes and microglia (LEE and BENVENISTE 1999). B7 costimulatory molecules are up-regulated during in¯ammation and have been detected on microglia in human multiple sclerosis acute lesions (WILLIAMS et al. 1994; DE SIMONE et al. 1995). In one experimental model of EAE, B7.2 was identi®ed on astrocytes and microglia/ macrophages during acute disease, and B7.1 was found on microglia, astrocytes, and neurons during remission (DANGOND et al. 1997). Although all of these proteins play important roles in CNS immune responses, double labeling studies need to be performed to con®rm the cell type expressing these molecules. Markers of Central Nervous System Glia and Neurons 133 5 Conclusions There are sucient markers currently available to clearly identify the major types of cells within the CNS. The best way to identify a cell type is to use a marker along with morphology, since either criterion alone can be ambiguous. Our current knowledge of cell types within the CNS is still developing, and markers available to identify them are works in progress. The number of cell type markers will certainly increase over time and will likely be followed by subsequent increases in the number of identi®able specialized cells in the CNS. Future increases in the knowledge of cell type markers in the CNS may: (1) help to more clearly de®ne distinct populations of astrocytes and neurons, (2) increase our understanding of adult oligodendrocyte progenitor function during normal conditions and in¯ammation, (3) help to better de®ne the phenotypes and functions of cells in the perivascular region, (4) provide the ability to distinguish between resident microglia and in®ltrating macrophages, and (5) help to identify functions of dierent cell types during disease. During pathological conditions, the speci®city of markers can change, and cell morphology can change. Therefore, clearly identifying cell types becomes much more dicult. Choosing the appropriate marker and showing morphology as clearly as possible are important strategies to use when identifying cell types during in¯ammation, viral infection or injury. Much research examining CNS disease heavily depends upon the ability to clearly identify a cell phenotype with an antigenic marker. However, the true complexity of in¯ammatory and disease processes in the CNS may not always conform to established labels and categories associated with cell type markers. Newly developed gene screening technologies may provide the ability to identify new distinct markers for CNS cell types. For example, a PCR-based gene expression screening assay called ``total gene expression analysis'' (TOGA) (SUTCLIFFE et al. 2000) is currently being used to identify genes expressed in activated microglia but not activated macrophages (M.J. Carson, manuscript in preparation). This work may lead to the characterization of novel markers that can be used to distinguish microglia from macrophages. Identi®cation of new markers to distinguish CNS cell types will most certainly help in understanding the remarkable responses of the CNS to injury and disease. Acknowledgements. We thank M.J. Carson, D.B. McGavern, and R.C. Armstrong for helpful comments on the manuscript, B.A. Barres for helpful comments regarding CC-1, and M.B.A. Oldstone for helpful suggestions and support. This work was supported in part by a grant to C.F.E. from the National Multiple Sclerosis Society, and by National Institutes of Health grants NS37135 (to C.F.E), AI43103 (to M.J.B.) and NS38719 (to M.B.A.O.). J.M.R. was supported by a National Institutes of Health Training Grant (5 T32 AG00080-21) and a postdoctoral fellowship from the National Multiple Sclerosis Society. This is manuscript #13739-NP from The Scripps Research Institute. 134 J.M. Redwine and C.F. Evans References Acarin L, Vela JM, Gonzales B, Castallano B (1994) Demonstration of poly-N-acetyl lactosamine residues in amoeboid and rami®ed microglial cells in rat bfain by tomato lectin binding. J Histochem Cytochem 42:1033±1041 Adams CW, Poston RN (1990) Macrophage histology in paran-embedded multiple sclerosis plaques is demonstrated by the monoclonal pan-macrophage marker HAM-56: correlation with chronicity of the lesion. Acta Neuropathol (Berl 80:208±11 al-Ali SY, al-Hussain SM (1996) An ultrastructural study of the phagocytic activity of astrocytes in adult rat brain. J Anat 188:257±62 Andjelkovic AV, Nikolic B, Pachter JS, Zecevic N (1998) Macrophages/microglial cells in human central nervous system during development: an immunohistochemical study. Brain Res 814:13±25 Austyn JM, Gordon S (1981) F4/80, a monoclonal antibody directed speci®cally against the mouse macrophage. Eur J Immunol 11:805±15 Bar-Or A, Oliveira EM, Anderson DE, Ha¯er DA (1999) Molecular pathogenesis of multiple sclerosis. J Neuroimmunol 100:252±259 Barnett SC, Franceschini IA (1999) Adhesion molecule expression and phenotype of glial cells in the olfactory tract. Adv Exp Med Biol 468:297±307 Bechmann I, Nitsch R (1997) Astrocytes and microglial cells incorporate degenerating ®bers following entorhinal lesion: a light, confocal, and electron microscopical study using a phagocytosis-dependent labeling technique. Glia 20:145±54 Beller DI, Springer TA, Schreiber RD (1982) Anti mac-1 selectively inhibits the mouse and human type three complement receptor. J Exp Med 156:1000±1009 Benkovic SA, Connor JR (1993) Ferritin, transferrin, and iron in selected regions of the adult and aged rat brain. J Comp Neurol 338:97±113 Bezzi P, Vesce S, Panzarasa P, Volterra A (1999) Astrocytes as active participants of glutamatergic function and regulators of its homeostasis. Adv Exp Med Biol 468:69±80 Bhat RV, Axt KJ, Fosnaugh JS, Smith KJ, Johnson KA, Hill DE, Kinzler KW, Baraban JM (1996) Expression of the APC tumor suppressor protein in oligodendroglia. Glia 17:169±74 Bignami A, Eng LF, Dahl D, Uyeda CT (1972) Localization of glial ®brillary acidic protein in astrocytes by immuno¯uorescence. Brain Res 43:429±435 Birling MC, Roussel G, Nussbaum F, Nussbaum JL (1993) Biochemical and immunohistochemical studies with speci®c polyclonal antibodies directed against bovine myelin/oligodendrocyte glycoprotein. Neurochem Res 18:937±45 Black JA, Waxman SG (1988) The perinodal astrocyte. Glia 1:169±83 Carson MJ, Reilly CR, Sutclie JG, Lo D (1998) Mature microglia resemble immature antigen-presenting cells. Glia 22:72±85 Carson MJ, Reilly CR, Sutclie JG, Lo D (1999) Disproportionate recruitment of CD8+ T cells into the central nervous system by professional antigen-presenting cells. Am J Pathol 154:481±94 Carson MJ, Sutclie JG, Campbell IL (1999) Microglia stimulate naive T-cell dierentiation without stimulating T-cell proliferation. J Neurosci Res 55:127±134 Chanas-Sacre G, Rogister B, Moonen G, Leprince P (2000) Radial glia phenotype: origin, regulation, and transdierentiation. J Neurosci Res 61:357±63 Chang A, Nishiyama A, Peterson J, Prineas J, Trapp BD (2000) NG2-positive oligodendrocyte progenitor cells in adult human brain and multiple sclerosis lesions. J Neurosci 20:6404±12 Conner JR, Fine RE (1986) The distribution of transferrin immunoreactivity in the rat central nervous system. Brain Res 368:319±328 Dangond F, Windhagen A, Groves CJ, Ha¯er DA (1997) Constitutive expression of costimulatory molecules by human microglia and its relevance to CNS autoimmunity. J Neuroimmunol 76:132±138 Dawson MR, Levine JM, Reynolds R (2000) NG2-expressing cells in the central nervous system: are they oligodendroglial progenitors? J Neurosci Res 61:471±479 De Simone R, Giampaolo A, Giometto B, Gallo P, Levi G, Peschle C, Aloisi F (1995) The costimulatory molecule B7 is expressed on human microglia in culture and in multiple sclerosis acute lesions. J Neuropath Exp Neurol 54:175±187 Dijkstra CD, Dopp EA, Joling P, Kraal G (1985) The heterogeneity of mononuclear phagocytes in lymphoid organs: distinct macrophage subpopulations in the rat recognized by monoclonal antibodies ED1, ED2 and ED3. Immunology 54:589±99 Markers of Central Nervous System Glia and Neurons 135 Dobersen MJ, Hammer JA, Noronha AB, MacIntosh TD, Trapp BD, Brady RO, Quarles RH (1985) Generation and characterization of mouse monoclonal antibodies to the myelin-associated glycoprotein (MAG). Neurochem Res 10:499±513 Eddleston M, de la Torre JC, Oldstone MBA, Loskuto DJ, Edgington TS, Mackman N (1993) Astrocytes are the primary source of tissue factor in the murine central nervous system; a role for astrocytes in cerebral hemostasis. J Clin Invest 92:349±358 Eglitis MA, Mezey E (1997) Hematopoietic cells dierentiate into both microglia and macroglia in the brains of adult mice. Proc Natl Acad Sci USA 94:4080±5 Elkabes S, Di Cicco-Bloom EM, Black IB (1996) Brain microglia/macrophages express neurotrophins that selectively regulate microglial proliferation and function. J Neurosci 16:2508±21 Evans CF, Horwitz MS, Hobbs MV, Oldstone MB (1996) Viral infection of transgenic mice expressing a viral protein in oligodendrocytes leads to chronic central nervous system autoimmune disease. J Exp Med 184:2371±84 Fedoro S, Zhai R, Novak JP (1997) Microglia and astroglia have a common progenitor cell. J Neurosci Res 50:477±486 rench-Constant C, Ra MC (1986) Proliferating bipotential glial progenitor cells in adult rat optic nerve. Nature 319:499±502 Ford AL, Goodsall AL, Hickey WF, Sedgwick JD (1995) Normal adult rami®ed microglia separated from other central nervous system macrophages by ¯ow cytometric sorting. Phenotypic dierences de®ned and direct ex vivo antigen presentation to myelin basic protein-reactive CD4+ T cells compared. J Immunol 154:4309±21 Forss-Petter S, Danielson P, Sutclie JG (1986) Neuron-speci®c enolase: complete structure of rat mRNA, multiple transcriptional start sites, and evidence suggesting post-transcriptional control. J Neurosci Res 16:141±56 Fox RJ, Levin MC, Jacobson S (1996) Tumor necrosis factor alpha expression in the spinal cord of human T-cell lymphotrophic virus type I associated myelopathy/tropical spastic paraparesis patients. J Neurovirol 2:323±9 Franke H, Krugel U, Illes P (1999) P2 receptor-mediated proliferative eects on astrocytes in vivo. Glia 28:190±200 Friedman B, Hock®eld S, Black JA, Woodru KA, Waxman SG (1989) In situ demonstration of mature oligodendrocytes and their processes: an immunocytochemical study with a new monoclonal antibody, Rip. Glia 2:380±390 Fuss B, Mallon B, Phan T, Ohlemeyer C, Kirchho F, Nishiyama A, Macklin WB (2000) Puri®cation and analysis of in vivo-dierentiated oligodendrocytes expressing the green ¯uorescent protein. Dev Biol 218:259±74 Gardinier MV, Matthieu JM (1993) Cloning and cDNA sequence analysis of myelin/oligodendrocyte glycoprotein: a novel member of the immunoglobulin gene superfamily. Schweiz Arch Neurol Psychiatr 144:201±7 Gehrmann J, Matsumoto Y, Kreutzberg GW (1995) Microglia: intrinsic immunoeector cell of the brain. Brain Res Rev 20:269±287 Gogate N, Verva L, Zhou J, Milward E, Rusten R, O'Conner M, Kufta C, Kim J, Hudson L, DuboisDalq M (1994) Plasticity in the adult human oligodendrocyte lineage. J Neurosci 14:4571±4587 Goldman JE, Reynolds R (1996) A reappraisal of ganglioside GD3 expression in the CNS. Glia 16:291±5 Goto S, Korematsu K, Nagahiro S, Ushio Y (1993) Distinct neuronal subset reveals perikaryal immunostaining for synaptophysin (protein p38) in the striatum of rats. Acta Neuropathol (Berl 86:302±5 Graeber MB, Streit W, Kreutzberg GW (1989) Identity of ED2-positive perivascular cells in rat brain. J Neurosci Res 22:103±106 Greer JM, Dyer CA, Pakaski M, Symonowicz C, Lees MB (1996) Orientation of myelin proteolipid protein in the oligodendrocyte cell membrane. Neurochem Res 21:431±40 Haan EA, Boss BD, Cowan WM (1982) Production and characterization of monoclonal antibodies against the ``brain-speci®c'' proteins 14-3-2 and S-100. Proc Natl Acad Sci USA 79:7585±7589 Hart IK, Richardson WD, Heldin CH, Westermark B, Ra MC (1989) PDGF receptors on cells of the oligodendrocyte-type-2 astrocyte (O 2 A) cell lineage. Development 105:595±603 Harvey AR, Symons NA, Pollett MA, Brooker GJ, Bartlett PF (1997) Fate of adult neural precursors grafted to adult cortex monitored with a Y-chromosome marker. Neuroreport 8:3939±43 Hauke C, Korr H (1993) RCA-I lectin histochemistry after trypsinisation enables the identi®cation of microglial cells in thin paran sections of the mouse brain. J Neurosci Methods 50:273±7 136 J.M. Redwine and C.F. Evans Hermanson M, Funa K, Hartman M, Claesson-Welsh L, Heldin CH, Westermark, Nister M (1992) Platelet-derived growth factor and its receptors in human glioma tissue: expression of messenger RNA and protein suggests the presence of autocrine and paracrine loops. Cancer Res 52:3213±3219 Hickey WF, Kimura H (1988) Perivascular microglial cells of the CNS are bone marrow derived and present antigen in vivo. Science 239:290±292 Hickey WF, Vass K, Lassman H (1992) Bone marrow-derived elements in the central nervous system: an immunohistochemical and ultrastructural survey of rat chimeras. J Neuropathol Exp Neurol 51: 246±256 Ho MK, Springer, TA (1982) Mac-1 antigen: quantitative expression in macrophage populations and tissues, and immuno¯uorescent localization in spleen. J Immunol 128:2281±2286 Holz A, Schaeren-Wiemers N, Schaefer C, Pott U, Colello RJ, Schwab ME (1996) Molecular and developmental characterization of novel cDNAs of the myelin-associated/oligodendrocytic basic protein. J Neurosci 16:467±77 Holz A, Schwab ME (1997) Developmental expression of the myelin gene MOBP in the rat nervous system. J Neurocytol 26:467±477 Horwitz MA (1992) Interactions between macrophages and legionella pneumophila. In: Russel SW, Gordon S (eds) Current topics in microbiology and immunology. Springer-Verlag, Berlin Horwitz MS, Evans CF, Klier FG, Oldstone MB (1999) Detailed in vivo analysis of interferon-gamma induced major histocompatibility complex expression in the central nervous system: astrocytes fail to express major histocompatibility complex class I and II molecules. Lab Invest 79:235±42 Horwitz MS, Evans CF, McGavern DB, Rodriguez M, Oldstone MBA (1997) Primary demyelination in transgenic mice expressing interferon-gamma. Nature Med 3:1037±1041 Huber G, Matus A (1984) Dierences in the cellular distributions of two microtubule-associated proteins, MAP1 and MAP2, in rat brain. J Neurosci 4:151±60 Hulette CM, Downey BT, Burger PC (1992) Macrophage markers in diagnostic neuropathology [published erratum appears in Am J Surg Pathol 1992 Oct; 16(10):1029]. Am J Surg Pathol 16:493±9 Katz-Levy Y, Neville KL, Girvin AM, Vanderlugt CL, Pope JG, Tan LJ, Miller SD (1999) Endogenous presentation of self myelin epitopes by CNS-resident APCs in Theiler's virus-infected mice. J Clin Invest 104:599±610 Keirstead HS, Levine JM, Blakemore WF (1998) Response of the oligodendrocyte progenitor cell population (de®ned by NG2 labelling) to demyelination of the adult spinal cord. Glia 22:161±70 Krivit W, Sung JH, Shapiro EG, Lockman LA (1995) Microglia: the eector cell for reconstitution of the central nervous system following bone marrow transplantation for lysosomal and peroxisomal storage diseases. Cell Transplant 4:385±92 Lampson LA (1995) Interpreting MHC class I expression and class I/classII reciprocity in the CNS: Reconciling divergent ®ndings. Microsc Res Tech 32:267±285 Lasser DM, DeVivo DC, Garvin J, Wilhemsen KC (1994) Turcot's syndrome: evidence for linkage to the adenomatous polyposis coli (APC) locus. Neurology 44:1083±1086 Lee SJ, Benveniste EN (1999) Adhesion molecule expression and regulation on cells of the central nervous system. J Neuroimmunol 98:77±88 Levi G, Aloisi F, Wilkin GP (1987) Dierentiation of cerebellar bipotential glial precursors into oligodendrocytes in primary culture: developmental pro®le of surface antigens and mitotic activity. J Neurosci Res 18:407±417 Levine JM (1994) Increased expression of the NG2 chondroitin-sulfate proteoglycan after brain injury. J Neurosci 14:4716±4730 Levine JM, Card JP (1987) Light and electron microscopic localization of a cell surface antigen (NG2) in the rat cerebellum: association with smooth protoplasmic astrocytes. J Neurosci 7:2711±2720 Levine JM, Reynolds R, Fawcett JW (2000) The oligodendrocyte precursor cell in health and disease. TINS (in press) Levine JM, Stallcup WB (1987) Plasticity of developing cerebellar cells in vitro studied with antibodies against the NG2 antigen. J Neurosci 7:2721±2731 Levine JM, Stincone F, Lee YS (1993) Development and dierentiation of glial precursor cells in the rat cerebellum. Glia 7:307±21 Levison SW, Goldman JE (1993) Both oligodendrocytes and astrocytes develop from progenitors in the subventricular zone of postnatal rat forebrain. Neuron 10:201±212 Lewis SA, Lee G-S, Cowan NJ (1985) Five mouse tubulin isotypes and their regulated expression during development. J Cell Biol 101:852±860 Ling EA, Wong WC (1993) The origin and nature of rami®ed and amoeboid microglia: a historical review and current concepts. Glia 7:9±18 Markers of Central Nervous System Glia and Neurons 137 Ludwin SK, Kosek JC, Eng LF (1976) The topographical distribution of S-100 and GFA proteins in the adult rat brain: an immunohistochemical study using horseradish peroxidase-labelled antibodies. J Comp Neurol 165:197±208 Luskin MB, McDermott K (1994) Divergent lineages for oligodendrocytes and astrocytes originating in the neonatal forebrain subventricular zone. Glia 3:211±226 Luskin MB, Parnavelas JG, Bar®eld JA (1993) Neurons, astrocytes, and oligodendrocytes of the rat cerebral cortex originate from separate progenitor cells: an ultrastructural analysis of clonally related cells. J Neurosci 13:1730±1750 Manoji H, Yeger H, Becher LE (1986) A speci®c histochemical marker (lectin ricinus communis agglutinin-1) for normal human microglia and application to routine histopathology. Acta Neuropathol 71:341±343 Marangos PJ, Schmechel D, Zis AP, Goodwin FK (1979) The existence and neurobiological signi®cance of neuronal and glial forms of the glycolytic enzyme enolase. Biol Psychiatry 14:563±79 Marangos PJ, Zis AP, Clark RL, Goodwin FK (1978) Neuronal, non-neuronal and hybrid forms of enolase in brain: structural, immunological and functional comparisons. Brain Res 150:117±33 Matsumoto Y, Ohmori K, Fujiwara M (1992) Immune regulation by brain cells in the central nervous system: microglia but not astrocytes present myelin basic protein to encephalitogenic T cells under in vivo-mimicking conditions. Immunol 76:209±216 McFarland HI, Nahill SR, Maciaszek JW, Welsh RM (1992) CD11b (mac-1); a marker for CD8+ cytotoxic T cell activation and memory in virus infection. J Immunol 149:1326±1333 McGarry RC, Helfand SL, Quarles RH, Roder JC (1983) Recognition of myelin-associated glycoprotein by the monoclonal antibody HNK-1. Nature 306:376±8 Mercken M, Lubke U, Vandermeeren M, Gheuens J, Oestreicher AB (1992) Immunocytochemical detection of the growth-associated protein B-50 by newly characterized monoclonal antibodies in human brain and muscle. J Neurobiol 23:309±21 Mercken M, Vandermeeren M, Lubke U, Six J, Boons J, Vanmechelen E, Van de Voorde A, Gheuens J (1992) Anity puri®cation of human tau proteins and the construction of a sensitive sandwich enzyme-linked immunosorbent assay for human tau detection. J Neurochem 58:548±53 MesserSmith DJ, Murtie JC, Le TQ, Frost EE, Armstrong RC (2000) Fibroblast growth factor 2 (FGF2) and FGF receptor expression in an experimental demyelinating disease with extensive remyelination. J Neurosci Res 62:241±256 Miller RH, Zhang H, Fok-Seang J (1994) Glial cell heterogeneity in the mammalian spinal cord. Perspect Dev Neurobiol 2:225±231 Miyake T, Okada M, Kitamura T (1992) Reactive proliferation of astrocytes studied by immunohistochemistry for proliferating cell nuclear antigen. Brain Res 590:300±2 Morris CS, Esiri MM, Sprinkle TJ, Gregson N (1994) Oligodendrocyte reactions and cell proliferation markers in human demyelinating diseases. Neuropathol Appl Neurobiol 20:272±81 Mullen RJ, Buck CR, Smith AM (1992) NeuN, a neuronal speci®c nuclear protein in vertebrates. Development 116:201±11 Muller T, Kettenmann H (1995) Physiology of bergmann glial cells. Int Rev Neurobiol 38:341±359 Nagata K, Takei N, Nakajima K, Saito H, Kohsaka S (1993) Microglial conditioned medium promotes survival and development of cultured mesencephalic neurons from embryonic rat brain. J Neurosci Res 34:357±63 Nishiyama A, Chang A, Trapp BD (1999) NG2+ glial cells: a novel glial cell population in the adult brain. J Neuropathol Exp Neurol 58:1113±24 Nishiyama A, Lin XH, Giese N, Heldin CH, Stallcup WB (1996) Co-localization of NG2 proteoglycan and PDGF alpha-receptor on O2 A progenitor cells in the developing rat brain. J Neurosci Res 43:299±314 Nishiyama A, Yu M, Drazba JA, Tuohy VK (1997) Normal and reactive NG2+ glial cells are distinct from resting and activated microglia. J Neurosci Res 48:299±312 Norton WT (1999) Cell reactions following acute brain injury: a review. Neurochem Res 24:213±8 Oumesmar BN, Vignais L, Baron-Van Evercooren A (1997) Developmental expression of platelet-derived growth factor alpha receptor in neurons and glial cells of the mouse CNS. J Neurosci 17:125±139 Pawlinski R, Janeczko K (1997) An inhibitory eect of interferon gamma on the injury-induced astrocyte proliferation in the postmitotic rat brain. Brain Res 773:231±4 Peters A, Palay SL, Webster Hd (1991) The ®ne structure of the nervous system. Oxford University Press, New York Pouly S, Becher B, Blain M, Antel JP (1999) Expression of a homologue of rat NG2 on human microglia. Glia 27:259±68 138 J.M. Redwine and C.F. Evans Powell SM, Zilz N, Beazer-Barclay Y, Bryan TM, Hamilton SR, Thibodeau SN, Vogelstein B, Kinzler KW (1992) APC mutations occur early during colorectal tumorgenesis. Nature 359:235±237 Pringle N, Mudhar HS, Collarini EJ, Richardson WD (1992) PDGF receptors in the rat CNS: during late neurogenesis, PDGF alpha receptor appears to be restricted to glial cells of the oligodendrocyte lineage. Development 115:535±551 Pringle NP, Richardson WD (1993) A singularity of PDGF alpha receptor expression in the dorsoventral axis of the neural tube may de®ne the origin of the oligodendrocyte lineage. Development 115: 535±551 Ramon-Cueto A, Valverde F (1995) Olfactory bulb ensheathing glia: a unique cell type with axonal growth-promoting properties. Glia 14:163±173 Redwine JM, Armstrong RC (1998) In vivo proliferation of oligodendrocyte progenitors expressing PDGF-alpha receptor during early remyelination. J Neurobiol 37:413±428 Redwine JM, Buchmeier MJ, Evans CF (2001) In vivo expression of MHC molecules on oligodendrocytes and neurons during viral infection. Am J Pathol 159:1219±1224 Reiser G, Kunzelmann U, Steinhilber G, BinmoÈller FJ (1994) Generation of a monoclonal antibody against the myelin protein CNP suitable for biochemical and for immunohistochemical investigations of CNP. Neurochem Res 19:1479±1485 Reynolds R, Hardy R (1997) Oligodendroglial progenitors labeled with the O4 antibody persist in the adult rat cerebral cortex in vivo. J Neurosci Res 47:455±470 Reynolds R, Wilkin GP (1988) Development of macroglial cells in the rat cerebellum II. An in situ immunohistochemical study of oligodendroglial lineage from precursor to mature myelinating cell. Development 102:409±425 Richardson A, Hao C, Fedoro S (1993) Microglia progenitor cells: a subpopulation on cultures of mouse neopallial astroglia. Glia 7:25±33 Rickmann M, Wol JR (1995) S100 protein expression in subpopulations of neurons of rat brain. Neuroscience 67:977±91 Robinson AP, White TM, Mason DW (1986) Macrophage heterogeneity in the rat as delineated by two monoclonal antibodies MRC OX-41 and MRC OX-42, the latter recognizing complement receptor type 3. Immunology 57:239±47 Rodriguez M, Pierce ML, Howie EA (1987) Immune response gene products (Ia antigens) on glial and endothelial cells in virus-induced demyelination. J Immunol 138:3438±3442 Schmechel D, Marangos PJ, Zis AP, Brightman M, Goodwin FK (1978) Brain endolases as speci®c markers of neuronal and glial cells. Science 199:313±5 Schumacher U, Adam E, Kretzschmar H, Pfuller U (1994) Binding patterns of mistletoe lectins I, II and III to microglia and Alzheimer plaque glycoproteins in human brains. Acta Histochem 96:399±403 Scolding N, Franklin R, Stevens S, Heldin CH, Compston A, Newcombe J (1998) Oligodendrocyte progenitors are present in the normal adult human CNS and in the lesions of multiple sclerosis. Brain 121:2221±8 Shaw G, Osborn M, Weber K (1981) An immuno¯uorescence microscopical study of the neuro®lament triplet proteins, vimentin and glial ®brillary acidic protein within the adult rat brain. Eur J Cell Biol 26:68±82 Sheedlo HJ, Sprinkle TJ (1983) The localization of 20 ,30 -cyclic nucleotide 30 -phosphodiesterase in bovine cerebrum by immuno¯uorescence. Brain Res 288:330±333 Shine HD, Readhead C, Popko B, Hood L, Sidman RL (1992) Morphometric analysis of normal, mutant, and transgenic CNS: correlation of myelin basic protein expression to myelinogenesis. J Neurochem 58:342±9 Sillevis Smitt PA, van der Loos C, Vianney de Jong JM, Troost D (1993) Tissue ®xation methods alter the immunohistochemical demonstrability of neuro®lament proteins, synaptophysin, and glial ®brillary acidic protein in human cerebellum. Acta Histochem 95:13±21 Smith KJ, Johnson KA, Bryan TM, Hill DE, Markowitz S, Willson JK, Paraskeva C, Petersen GM, Hamilton SR, Vogelstein B, et al. (1993) The APC gene product in normal and tumor cells. Proc Natl Acad Sci U S A 90:2846±50 Sobel RA, Greer JM, Isaac J, Fondren G, Lees MB (1994) Immunolocalization of proteolipid protein peptide 103±116 in myelin. J Neurosci Res 37:36±43 Sommer I, Schachner M (1981) Monoclonal antibodies (O1 and O4) to oligodendrocyte cell surfaces: an immunocytological study in the central nervous system. Dev Biol 83:328±228 Sorensen T, Tani M, Jensen J, Pierce V, Lucchinetti C, Folcik VA, Qin S, Rottman J, Sellebjerg F, Strieter RM, Frederiksen JL, Ransoho RM (1999) Expression of speci®c chemokines and chemokine receptors in the central nervous system of multiple sclerosis patients. J Clin Invest 103:807±815 Markers of Central Nervous System Glia and Neurons 139 Stallcup WB (1981) The NG2 antigen, a putative lineage marker: immuno¯uorescent localization in primary cultures of rat brain. Dev Biol 83:154±165 Stallcup WB, Beasley L (1987) Bipotential glial precursor cells of the optic nerve express the NG2 proteoglycan. J Neurosci 7:2737±2744 Stallcup WB, Beasley L, Levine J (1983) Cell-surface molecules that characterize dierent stages in the development of cerebellar interneurons. Cold Spr Harbor Symp Quant Biol 48:761±774 Starkey PM, Turley L, Gordon S (1987) The mouse macrophage-speci®c glycoprotein de®ned by monoclonal antibody F4/80: characterization, biosynthesis and demonstration of a rat analogue. Immunology 60:117±22 Sternberger NH, Itoyama Y, Kies MW, deF Webster H (1978) Myelin basic protein demonstrated immunocytochemically in oligodendroglia prior to myelin sheath formation. Proc Natl Acad Sci USA 75:2521±2524 Sternberger NH, Quarles RH, Itoyama Y, deF Webster H (1979) Myelin associated glycoprotein demonstrated immunocytochemically in myelin and myelin-forming cells of developing rats. Proc Natl Acad Sci USA 76:1510±1514 Streit WJ, Graeber MB, Kreutzberg GW (1988) Functional plasticity of microglia: a review. Glia 1:301±7 Streit WJ, Kreitzberg GW (1988) Response of endogenous glial cells to motor neuron degeneration induced by toxic ricin. J Comp Neurol 268:248±263 Streit WJ, Kreutzberg GW (1987) Lectin binding by resting and reactive microglia. J Neurocytol 16: 249±60 Stridsberg M, Lundqvist G, Engstrom U, Wilander E, Su H, Gobl A, Oberg K (1994) Development of polyclonal antibodies and evaluation of a sensitive radioimmunoassay for detection and measurement of synaptophysin. Acta Neuropathol (Berl) 87:635±41 Sun N, Grzybicki D, Castro RF, Murphy S, Perlman S (1995) Activation of astrocytes in the spinal cord of mice chronically infected with a neurotropic coronavirus. Virology 213:482±93 Sutclie JG, Foye PE, Erlander MG, Hilbush BS, Bodzin LJ, Durham JT, Hasel KW (2000) TOGA: an automated parsing technology for analyzing expression of nearly all genes. Proc Natl Acad Sci USA 97:1976±81 Suzuki H, Franz H, Yamamato Y, Iwasaki H, Konno H (1988) Identi®cation of normal microglia population in human and rodent nervous tissue using lectin-histochemistry. J Neuropathol Appl Neurobiol 14:221±227 Todd AJ, Spike RC, Polgar E (1998) A quantitative study of neurons which express neurokinin-1 or somatostatin sst2a receptor in rat spinal dorsal horn. Neuroscience 85:459±73 Trapp BD, Andrews SB, Cootauco C, Quarles R (1989) The myelin-associated glycoprotein is enriched in multivesicular bodies and periaxonal membranes of actively myelinating oligodendrocytes. J Cell Biol 109:2417±26 Tucker RP, Binder LI, Matus AI (1988) Neuronal microtubule-associated proteins in the embryonic avian spinal cord. J Comp Neurol 271:44±55 Ulvestad E, Williams K, Mork S, Antel J, Nyland H (1994) Phenotypic dierences between human monocytes/macrophages and microglial cells studied in situ and in vitro. J Neuropathol Exp Neurol 53:492±502 Vijayan VK, Lee YL, Eng LF (1990) Increase in glial ®brillary acidic protein following neural trauma. Mol Chem Neuropathol 13:107±18 Warschkau H, Kiderlen AF (1999) A monoclonal antibody directed against the nurine macrophage surface molecule F4/80 modulates natural immune response to listeria monocytogenes. J Immunol 163:3409±3416 Wenzel J, Lammert G, Meyer U, Krug M (1991) The in¯uence of long-term potentiation on the spatial relationship between astrocyte processes and potentiated synapses in the dentate gyrus neuropil of rat brain. Brain Res 560:122±31 Westergaard N, Sonnewald U, Schousboe A (1995) Metabolic tracking between neurons and astrocytes: the glutamate/glutamine cycle revisited. Dev Neurosci 17:203±11 Whitton JL, Tishon A, Lewicki H, Gebhard J, Cook T, Salvato M, Joly E, Oldstone MB (1989) Molecular analyses of a ®ve-amino-acid cytotoxic T-lymphocyte (CTL) epitope: an immunodominant region which induces nonreciprocal CTL cross-reactivity. J Virol 63:4303±10 Wiedenmann B, Franke WW (1985) Identi®cation and localization of synaptophysin, an integral membrane glycoprotein of Mr 38,000 characteristic of presynaptic vesicles. Cell 41:1017±28 Williams K, Ulvestad E, Antel JP (1994) B7/BB-1 antigen expression on adult human microglia studied in vitro and in situ. Eur J Immunol 24:3031±7 140 J.M. Redwine and C.F. Evans: Markers of Central Nervous System Glia and Neurons Wilson S-S, Baetge EE, Stallcup WB (1980) Antisera speci®c for cell lines with mixed neuronal and glial properties. Dev Biol 83:146±153 Wilson S-S, Baetge EE, Stallcup WB (1981) Antisera speci®c for cell lines with mixed neuronal and glial properties. Dev Biol 83:146±153 Wolswijk G, Noble M (1989) Identi®cation of an adult-speci®c glial progenitor cell. Development 105:387±400 Yamashita N, Ilg EC, Schafer BW, Heizmann CW, Kosaka T (1999) Distribution of a speci®c calciumbinding protein of the S100 protein family, S100A6 (calcyclin), in subpopulations of neurons and glial cells of the adult rat nervous system. J Comp Neurol 404:235±57 Yong VW, Moumdjian R, Yong FP, Ruijs TC, Freedman MS, Cashman N, Antel JP (1991) Gammainterferon promotes proliferation of adult human astrocytes in vitro and reactive gliosis in the adult mouse brain in vivo. Proc Natl Acad Sci USA 88:7016±20 Zheng-Fischhofer Q, Biernat J, Mandelkow EM, Illenberger S, Godemann R, Mandelkow E (1998) Sequential phosphorylation of Tau by glycogen synthase kinase-3beta and protein kinase A at Thr212 and Ser214 generates the Alzheimer-speci®c epitope of antibody AT100 and requires a paired-helical®lament-like conformation. Eur J Biochem 252:542±52