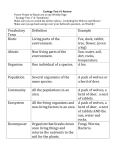
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Grade 9 Academic Science – Ecology Wolves and Yellowstone Hamann, Jack. 1997. Wolves Rreturn to Yellowstone Sparks Controversy. CNN. http://www.cnn.com/EARTH/9711/12/yellowstone.wolves/ The meals are catered, but the menu lacks variety -- cold carcasses of deer and elk road-kill served up to a wolfpack on probation, locked in a pen. The Nez Perce Pack is at the centre of a firestorm over which wolves should live and which should die at Yellowstone National Park. Named for a tribe of American Indians who fled through the park hoping for sanctuary in Canada, the pack of wolves is also looking for a safe haven in this remote wilderness. It is been almost three years since the federal government began the controversial measure of reintroducing wolves into Yellowstone. While conservationists supported the move, ranchers were wary. Breeding program successful Some 80 wolves now live in or near the park, many of them pups. Wolves are held in "acclimation pens" where, it is hoped, they will get used to their surroundings and form packs. Packs, it is believed, are less likely to stray from the park than stragglers. "In general, the acclimation has worked," says Doug Smith of Yellowstone Park. "Of the 31 wolves brought from Canada, there was really only one wolf, Number 27, where we feel acclimation did not work." Number 27 killed almost 50 sheep a few weeks ago, and had to be destroyed. The rancher who lost sheep was compensated with money from Defenders of Wildlife, a pro-wolf group. How great a threat? The 50 lost sheep attracted attention, but were hardly a rarity. In Montana alone, tens of thousands of freerange livestock die of various causes every year. "Predators are just a minute portion of that," says Ed Bangs of the National Fish and Wildlife Service. "Wolves are an immeasurable portion of that. On average, wolves kill about four or five a year, so industry wise, wolf predation means nothing to the industry or the economy of this area." "Some ranchers say, 'Look, it's people or wolves, not both,'" Smith says. "I think we're at point now with wildlife and biodiversity and conservation biology in the United States that we can do better. It is not black or white. We can have both." While the wolves pose some threat to humans' domesticated animals, there is little risk to people. While humans have killed an estimated two million wolves in this century, there is not a single documented case of a human being killed by a healthy wild wolf. A pack of trouble As biologists and rangers approach the Nez Perce pack's chain-link pen deep in the park's interior, they hear a surprising sound -- a wolf barking from outside. Somehow the alpha male, the leader of the pack, has escaped, only two weeks after he was recaptured. In his last escape, he dug under the fence and freed the rest of the pack. "I think one's out," says biologist Carrie Shaefer. "That bark is an aggressive, defensive behavior." If the pack escapes before spring when they are scheduled for release, they will likely threaten livestock and will have to be destroyed. The Nez Perce pack was released in April 1996, and immediately lost "pack unity," the National Fish and Wildlife Service said. Individuals strayed into areas with little wild prey and plenty of livestock, a handful of sheep and cattle were killed and two pack members were shot dead. Since most of the Nez Perce pack are still in the pen, recapturing the stray is not Bangs' top priority. "I think the main thing is to keep the fence running, because he's not going anywhere," he says. "Not with his woman or buddies in the pen. He is a family-oriented animal." In the cramped offices of the Wolf Recovery Program, Smith and Bangs struggle over the fate of fugitive wolves. The return of the wolves to Yellowstone requires a balancing act among ranchers and conservationists, a dispute with these men in the middle. "I am getting a call from a rancher, yelling at me because the wolves are out and the killed some sheep," Bangs says. "The next phone call is from an animal rights person saying I am a bastard who should burn in Hell for murdering babies. Then the next call is another rancher." The wolves already in Yellowstone have bred so successfully that plans to transplant other wolves into the area are on hold. That is a success story to some, but how success will be defined -- and what it will cost -is the ongoing debate. Yellowstone and the Gray Wolf American Farm Bureau Federation. 1998. AFBF Responds to Distortions by Defenders of Wildlife. http://www.fb.org/news/graywolf.html Defenders of Wildlife, in its attempt to persuade the American Farm Bureau Federation to drop its lawsuit against the Interior Department’s Canadian Gray Wolf Introduction Program in the Yellowstone and central Idaho region, has distorted the nation's largest farm organization's position in the case. The American Farm Bureau Federation's arguments, which are fully supported by Judge William Downes, have been made clear. The following points are intended to clarify some of the distortions generated by Defenders of Wildlife and other groups. The American Farm Bureau Federation filed its lawsuit, in conjunction with the Montana, Idaho and Wyoming Farm Bureaus, because the Interior Department broke the law in launching the wolf introduction program in 1994. Interior Secretary Bruce Babbitt violated provisions of the Endangered Species Act when he brought the gray wolves into the Yellowstone and central Idaho region. The gray wolf is not threatened by extinction. Bringing them into the Yellowstone and central Idaho region under the guise of the Endangered Species Act was illegal. The American Farm Bureau Federation never has advocated the killing of the Canadian gray wolves as a means of their removal from the Yellowstone and central Idaho region. The Interior Department assured Judge Downes that it could recapture and relocate the wolves from the Yellowstone and central Idaho region. It is Farm Bureau’s hope that the federal government backs up its assertion that it could easily capture and remove all Canadian wolves from the Yellowstone and central Idaho region. Defenders of Wildlife -- not Farm Bureau -- is the only organization that continues to raise the issue of killing wolves as a means of removal. In a Jan. 16, 1998 follow-up ruling to his Dec. 12, 1997 decision, Judge William Downes wrote, “The evidence before this court revealed that the Defendants (the Interior Department) could remove nonnative wolves from the experimental population areas and transplant them elsewhere within the territory of the United States. The record suggests that if non-native wolves can be humanely captured and transferred to the experimental population areas, they can also be humanely captured and removed. The order to remove the wolves is not intended to serve as a license to euthanize wolves and any interpretation that it does so is misplaced.” This issue is about the rule of law. It’s a matter of what’s right -- and legal. Interior Secretary Bruce Babbitt ignored the implications the Wolf Introduction Program would have on native wolves, ranchers and their livestock when he launched the program. Because of this overly ambitious program, farmers and ranchers were forced into a position of defending their property and rights. The American Farm Bureau Federation tried to stop the introduction of the Canadian gray wolves to the Yellowstone and central Idaho region before the Gray Wolf Reintroduction Program was launched. Farm Bureau’s suit was filed before any Canadian wolves were even captured. Yet, in the face of our lawsuit, the Interior Department, knowing that the court could find the program illegal and order the Canadian gray wolves’ removal, brought the Canadian wolves into the United States before the court could decide the legal issues. Wolf Restoration to Yellowstone USA National Park Service - Yellowstone National Park. 2006. Wolf Restoration to Yellowstone. http://www.nps.gov/yell/nature/animals/wolf/wolfrest.html Northern Rocky Mountain wolves, a subspecies of the gray wolf (Canis lupus), were native to Yellowstone when the park was established in 1872. Predator control was practiced here in the late 1800s and early 1900s. Between 1914 and 1926, at least 136 wolves were killed in the park; by the 1940s, wolf packs were rarely reported. By the 1970s, scientists found no evidence of a wolf population in Yellowstone; wolves persisted in the lower 48 states only in northern Minnesota and on Isle Royale in Michigan. An occasional wolf likely wandered into the Yellowstone area; however, no verifiable evidence of a breeding pair of wolves existed through the mid 1990s. In the early 1980s, wolves began to reestablish themselves near Glacier National Park in northern Montana; an estimated 75 wolves inhabited Montana in 1996. At the same time, wolf reports were increasing in central and north-central Idaho, and wolves were occasionally reported in the state of Washington. The wolf is listed as "endangered" throughout its historic range in the lower 48 states except in Minnesota, where it is "threatened." National Park Service (NPS) policy calls for restoring native species when: a) sufficient habitat exists to support a self-perpetuating population, b) management can prevent serious threats to outside interests, c) the restored subspecies most nearly resembles the extirpated subspecies, and d) extirpation resulted from human activities. The U.S. Fish & Wildlife Service 1987 Northern Rocky Mountain Wolf Recovery Plan proposed reintroduction of an "experimental population" of wolves into Yellowstone. In a report to Congress, scientists from the University of Wyoming predicted reductions of elk (15%-25%), bison (5%-15%), moose, and mule deer could result from wolf restoration in Yellowstone. A separate panel of 15 experts predicted decreases in moose (10%-15%) and mule deer (20%-30%). Minor effects were predicted for grizzly bears and mountain lions. Coyotes probably would decline and red foxes probably would increase. In October 1991, Congress provided funds to the U.S Fish & Wildlife Service (USFWS) to prepare, in consultation with the NPS and the U.S. Forest Service, an Environmental Impact Statement (EIS) on restoring wolves to Yellowstone and central Idaho. After several years and a near-record number of public comments, the Secretary of Interior signed the Record of Decision on the Final Environmental Impact Statement (FEIS) for reintroduction of gray wolves to both areas. Staff from Yellowstone, the USFWS, and participating states prepared to implement wolf restoration. The USFWS prepared special regulations outlining how wolves would be managed as a nonessential experimental population under section 10(j) of the Endangered Species Act. These regulations took effect in November 1994. As outlined in the Record of Decision, the states and tribes would implement and lead wolf management outside the boundaries of national parks and wildlife refuges, within federal guidelines. The states of Idaho, Wyoming, and Montana have begun preparation of wolf management plans. Park staff assisted with planning for a soft release of wolves in Yellowstone. This technique has been used to restore red wolves in the southeastern United States and swift fox in the Great Plains and involves holding animals temporarily in areas of suitable habitat. Penning of the animals is intended to discourage immediate long-distance dispersal. In contrast, a hard release allows animals to disperse immediately wherever they choose, and has been used in Idaho where there is limited access to the central Idaho wilderness. In the autumn of 1995 at three sites in the Lamar Valley, park staff completed site plans, and archaeological and sensitive plant surveys. Approximately 1 acre was enclosed at each site with 9-gauge chain link fence in 10' x 10' panels. These enclosures could be dismantled and reconstructed at other sites if necessary. The fences had a 2' overhang and a 4' skirt at the bottom to discourage climbing over or digging under the enclosure. Each pen had a small holding area attached, to allow a wolf to be separated from the group for medical treatment. Inside each pen were several plywood security boxes to provide shelter. For the 1996 release, one pen was relocated to Blacktail Plateau and another was constructed in the Firehole Valley in central Yellowstone. Subsequently pens have been relocated from Lamar to other areas in the park interior to facilitate releases into other geographic areas or the park or special circumstances that require the temporary penning of wolves. USFWS and Canadian wildlife biologists captured wolves in Canada and released them in both recovery areas in 1995 and 1996. As planned, wolves of dispersal age (1-2 years old) were released in Idaho, while Yellowstone released pups of the year (7+ months old), together with one or more of the alpha pair (breeding adults). Young pups weigh about 75 lbs. and are less likely to have established a home range. The goal was to have 5-7 wolves from one social group together in each release pen. Each wolf was radio-collared when captured in Canada. For about 8-10 weeks while temporarily penned, the wolves experienced minimal human contact. Approximately once each week, they were fed road-kills. They were guarded by rangers and other volunteers who minimized the amount of visual contact between wolves and humans. The pen sites and surrounding areas were closed and marked to prevent unauthorized entry. Biologists used radio-telemetry to check on the welfare of wolves. Although concern was expressed about the wolves becoming habituated to humans or to the captive conditions, the temporary holding period was not long in the life of a wolf. In Alaska and Canada, wolves are seldom known to develop the habituated behaviors seen more commonly in grizzly bears. Wolves, while social among their own kind, typically avoid human contact. They are highly efficient predators with welldeveloped predatory instincts. Their social structure and pack behavior minimizes their need to scavenge food or garbage available from human sources. Compared to bears, whose diet is predominantly vegetarian, wolves have less specific habitat requirements. The wolves' primary need is for prey, which is most likely to be elk, deer, and other ungulates in these recovery areas. In 1995, fourteen wolves were released into Yellowstone National Park. In 1996, seventeen more wolves were brought from Canada and released. After release, several thousand visitors were lucky to view wolves chasing and killing elk or interacting with bears during spring. A park ranger and a group of visitors watched a most exciting encounter between two packs which likely resulted in one young wolf's death. This was not the first fatal encounter between wolves, although human-caused mortalities still outnumber inter-pack strife as a cause of wolf deaths. Yellowstone's first fourteen wolves bore two litters totaling nine pups. In 1996, four packs produced fourteen pups. After the wolves’ release in 1996, plans to transplant additional wolves were terminated due to reduced funding and due to the wolves' unexpected early reproductive success. In early 1997, ten young wolves, orphaned when their parents were involved in livestock depredation on the Rocky Mountain Front in northwestern Montana, were released into the park. In the spring of the wolf restoration project's third year, nine packs of wolves produced 13 litters of 64 pups. Three of the packs produced multiple litters which, while documented in the literature, is still unusual. Alpha male wolves generally do not breed with their own offspring, possibly to prevent inbreeding. However, as wolves were matched up during temporary periods of penning and as pack members shifted or were killed and replaced by other dispersing wolves, the occasional result has been packs in which one or both of the alpha pair were not the parents of subordinate pack members. Consequently, the alpha males probably had less incentive to breed with only one female, especially since food was abundant and the packs were still in the early stages of establishing their territories. Lone wolves continued to roam widely, but most of the wolves remained primarily within the boundaries of Yellowstone National Park An estimated 100,000 park visitors have observed wolves since their return in 1995. The program's visibility has resulted in opportunities to educate audiences about predator-prey relationships, endangered species restoration, and the importance of maintaining intact ecosystems. The program has also generated numerous partnerships with private groups and individuals who generously donated their time and money— critical in an era of reduced budgets and staff downsizing. For both Idaho and Yellowstone, wolf population recovery is defined as having about 100 wolves, or approximately 10 breeding pairs, established in each area for 3 successive years. The goal to restore wolves and begin delisting them by approximately 2002 appears within reach. The return of the only species known to be missing from the world's first national park for the past half-century has been a milestone in ecological restoration. It has not only restored the wildlife complement of greater Yellowstone; it has been a symbolic victory for conservationists who patiently and persistently reversed the once-dominant attitude against predators to one of acceptance. We believe that Aldo Leopold would be proud that so many humans have come to respect even these "killer creatures" with whom we share the Earth. Yellowstone Park and Wolf Reintroduction Ives. Sarah. 2004. Wolves Reshape Yellowstone National Park. National Geographic. http://news.nationalgeographic.com/kids/2004/03/wolvesyellowstone.html In the late 1990s scientists began to notice changes in Yellowstone National Park, in Wyoming. Trees that had stopped growing decades ago began to grow again. Animals started behaving differently. What caused these mysterious changes? Scientists believe they have found the answer: Wolves used to live in Yellowstone, but many people thought wolves were pests that killed livestock and harmed crops. In 1926 the last wolf in Yellowstone was shot and killed. For the next 70 years Yellowstone did not have any wolves. Scientists decided to bring them back to the area in 1995. The U.S. Fish and Wildlife Service brought in 15 gray wolves from Canada. "Wolves were reintroduced because historically wolves lived in Yellowstone," explained William Ripple, a forest ecologist at Oregon State University. Ripple studies wolves' effects on Yellowstone. "Now with the reintroduction of wolves, Yellowstone has all of the top predators it has had for thousands of years." Today between 250 and 300 wolves live in Yellowstone, and they have already left their mark. "The effect has been dramatic," said Douglas Smith, leader of the Yellowstone Wolf Project. For example, wolves have been hunting and eating elk, a member of the deer family. The elk leftovers provide food for animals such as ravens, eagles, and bears. Wolves also scare elk from streams. With fewer elk near the water, plants that grow there, such as willows, can grow taller. "Benefits of this new plant growth include more habitat for birds and more plant food for beaver," Ripple explained. "The number of beaver in northern Yellowstone has increased dramatically since wolves were reintroduced." Wolves have such a big effect on Yellowstone because scientists believe wolves are a keystone species. Keystone species are species on which a large number of other plants and animals depend. "The removal of a keystone species can lead to the extinction of other species," Ripple said. According to Smith, the reintroduction of wolves has affected more than 25 species in Yellowstone. "We think that Yellowstone will be a different place in 20 years because of wolves," Smith said. Not everyone is happy with the wolves' return, however. Some farmers say that wolves kill their animals. "Ranchers want to limit the number of wolves in the area, because if there are too many wolves, they always get in trouble," said Jennifer Ellis of the Idaho Cattle Association. "Many people who live in wolf-inhabited areas have had not only their cows, calves, and sheep killed by wolves, but also their cats, dogs, and horses." Ripple noted that there is a program in place to pay farmers for any money they lose due to wolves killing cattle. Some farmers may not be happy, but many scientists believe that Yellowstone is starting to look more like it did before people began to interfere with the animals there. Reintroduction of Wolves to Yellowstone 2003. Wolves Are Rebalancing Yellowstone Ecosystem. Science Daily. http://www.sciencedaily.com/releases/2003/10/031029064909.htm The reintroduction of wolves into Yellowstone National Park may be the key to maintaining groves of cottonwood trees that were well on their way to localized extinction, and is working to rebalance a stream ecosystem in the park for the first time in seven decades, Oregon State University scientists say in two new studies. The data show a clear and remarkable linkage between the presence of wolves and the health of an entire streamside ecosystem, including two species of cottonwoods and the myriad of roles they play in erosion control, stream health, and nurturing diverse plant and animal life. The findings of these studies were recently published in Ecological Applications, a journal of the Ecological Society of America, and the journal Forest Ecology and Management. "In one portion of the elk's winter range along the Lamar River of Yellowstone National Park, we found that there were thousands of small cottonwood seedlings," said Robert Beschta, professor emeritus in the College of Forestry at OSU and an expert on streams and riparian systems. "There should also have been hundreds of young trees, but there were none. Long-term elk browsing had been preventing any seedlings from getting taller." That pattern was common throughout the study area - lots of seedlings in combination with large cottonwood trees generally more than 70 years old, but little or nothing in between. Young cottonwoods, willows, and other streamside woody species are a preferred food for browsing elk during the harsh winters in northern Yellowstone, when much of the other forage is buried under snow. But when packs of wolves historically roamed the area, food was not the only consideration for elk, which had to be very careful and apparently avoided browsing in high-risk areas with low visibility or escape barriers. Wolves were systematically killed in the Yellowstone region and many other areas of the West beginning in the late 1800s. A concentrated effort between 1914 and 1926 finished the job - the last known wolf pack disappeared in 1926. "I considered a variety of potential reasons that might explain the historical decline of cottonwoods that began in the 1920s and have continued up to the last couple of years," said Beschta. "I looked at climate change, lack of floods, fire suppression, natural stand dynamics, and numbers of elk. But none of those factors really explained the problem. "Ultimately, it became clear that wolves were the answer." While elk populations fluctuated over the decades when wolves were absent, browsing behavior appears to represent an important factor related to streamside impacts. With no fear of wolves, the elk could graze anywhere they liked and for decades have been able to kill, by browsing, nearly all the young cottonwoods. Other streamside species such as willows and berry-producing shrubs also suffered. That in turn began to play havoc with an entire streamside ecosystem and associated wildlife, including birds, insects, fish and others. Trees and shrubs were lost that could have helped control stream erosion. Food webs broke down. "Before the wolves came back, it was pretty clear that in some areas we were heading towards an outright extinction of cottonwoods," Beschta said. Now, with the recent reintroduction of wolves back into Yellowstone in 1995, streamside shrubs and cottonwoods within the Lamar Valley are beginning to become more prevalent and taller, and were the focus of a second study in the same area. That study outlines how the fear of attack by wolves apparently prevents browsing elk from eating young cottonwood and willows in some streamside zones. With the renewed presence of wolves, young cottonwoods and willows have been growing taller each year over the last four years on "high-risk" sites, where elk apparently feel vulnerable due to terrain or other conditions that might prevent escape. In contrast, on "low-risk" sites, they are still being browsed by elk and show little increase in height. "In one case where a gully formed an escape barrier for elk, the tree height went up proportionally as the gully deepened and formed an increasing barrier to escape," said William Ripple, a professor with the College of Forestry at OSU. "Where the fear factor of wolves is high, the young trees and willows are doing much better and growing taller." Traditionally, "keystone" predators such as wolves were known to influence the population of other animals that they preyed on directly, such as elk or antelope. What researchers are now coming to better understand is the "trophic effect," or cascade of changes that can take place in an ecosystem when an important part is removed, Ripple said. The comparatively pristine conditions of a national park allowed this type of research to make "cause and effect" studies more feasible, the scientists point out. "The removal of wolves for 70 years - and then their return - actually set the stage for a scientific experiment with fairly compelling results," Beschta said. In a larger context, the studies also raise valid questions about other complex and poorly understood interactions between plants, animals, and wildlife in disturbed ecosystems across much of the American West, and perhaps elsewhere in the world, the scientists say. In some areas of the West, the disappearance of up to 90 percent of the aspen trees has been documented - another species of plant that is also highly vulnerable to animal browsing when it is young. "The last period when aspen trees in Yellowstone escaped the effects of elk browsing to generate trees into the forest overstory was the 1920s," Ripple said, "which is also when wolves were removed from the park." In at least one place - America's first national park - there is now cause for hope. While it is too early to confirm the widespread recovery of cottonwoods and willows, the reintroduction of wolves appears to have put a stop to major declines in the survival of these plants, the researchers found. "One point that should not be missed is this is actually great news for the potential recovery of cottonwood trees and mature willows in Yellowstone National Park," Ripple said. "We now have a pretty good idea why they were in decline and the return of wolves should help pave the way for their recovery. Even though it may take a very long time, for a change it looks like we're headed in the right direction." Yellowstone Wolves - Data Table 1. Yellowstone National Park Wolf Pack Sizes October – December 2003 Pack Name Pup Count Summer Pup Count Fall Adults Yearlings Total Pack Group 1 5 0 2 2 Agate Creek 8 4 5 9 Bechler 4 4 4 8 Buffalo Fork 2 0 3 3 Chief Joseph 5 5 2 7 Cougar Creek 4 4 6 10 Druid Creek 13 9 8 17 Geode Creek 8 2 5 7 Gibbon* 5 5 - - Leopold 8 8 11 19 Mollie 2 2 5 7 Nez Perce 2 2 13 15 Rose Creek 4 4 2 6 Slough Creek 6 6 9 15 Swan Lake 6 6 14 20 Yellowstone Delta 4 4 13 17 Loners - - 2 2 TOTAL 79 60 109 169 *Breeding female died June 2003 Source – Defenders of Wildlife. 2004. Wolves at Yellowstone. http://www.defenders.org/wildlife/wolf/ynp.html The current status of the 1995 wolves by individual wolf. January 2003 In March 1995, 14 wolves were released in Yellowstone National Park. When the last of the original 14 died, the wolf population was 148 wolves within the Park and 270 wolves in the Greater Yellowstone ecosystem population (includes the Park). Table 2. Current status of 1995 wolves in Yellowstone Location Number Age Weight (pounds) Crystal Creek Rose Creek Soda Butte 1 Current Status pup 77 Alpha male Leopold Pack. 1996-2002, killed by rival pack 2 pup 80 Killed 1996 3 adult male 98 Was alpha male Crystal Creek Pack killed by the Druids 1996 4 adult female 100 Alpha female of the Crystal Creek Pack until early 2000. Became a lone wolf. Her radio-collar failed. As of 2002 presumed to have died, but her body never found 5 pup 75 Was alpha male of the Crystal Creek Pack killed by an elk 1998 6 pup 72 Alpha male Rose Creek Pack. Natural mortality July 2000. 1 pup 77 Alpha female of the naturally formed Leopold Pack, until late spring of 2002 when she was killed by another wolf pack. 2 adult female 98 Alpha female of the Rose Creek Pack from 199599. Later she became a beta member of the new Beartooth Pack east of Yellowstone. Most famous wolf in the Yellowstone recovery area. She presumably died sometime after fall 2001 3 adult 122 Illegally shot spring of 1995 1 adult 92 Illegally shot spring of 1996 2 adult 113 Illegally shot spring of 1995 3 adult 113 Was alpha male Soda Butte Pack died of old age March 1997 4 adult 89 Alpha female Soda Butte Pack 1995 to April 2000. Probably killed by a moose. 5 pup 75 Alpha male of the naturally-formed Washakie Pack Control kill 1998 Source – Defenders of Wildlife. 2004. Wolves at Yellowstone. http://www.defenders.org/wildlife/wolf/ynp.html DelGiudice, Glenn D. 1998. The Ecological Relationship of Gray Wolves and White-Tailed Deer in Minnesota. Forest Wildlife Populations & Research Group - Minnesota Department of Natural Resources. http://www.wolf.org/wolves/learn/scientific/delguidice.asp INTRODUCTION To best understand and appreciate the relationship of gray wolves (Canis lupus) and white-tailed deer (Odocoileus virginianus) in Minnesota today, information concerning their evolutionary backgrounds is helpful. Modern white-tailed deer have been evolving for about 1 million years (since the Pleistocene epoch). However, artiodactyls (even-toed ungulates or hooved animals) from which they evolved have existed for at least 58 million years (Kulp 1961). Similarly, wolves have existed in their present form for 1-2 million years, but evolved from flesh-eating ancestors that occurred some 45-55 million year ago (Matthew 1930). Because wolves have ranged over most of the Northern Hemisphere, a greater area than inhabited by any of their prey, primary species in the wolf's diet, has and still, depends on geographic location (Mech 1970). Today in the Great Lakes region, and in Minnesota specifically, white-tailed deer are the primary prey of wolves, with moose (Alces alc es) typically being of secondary importance where they both exist (Stenlund 1955, Mech and Frenzel 1971, Van Ballenberghe et al. 1975, Fritts and Mech 1981, Fuller 1989). Clearly, white-tailed deer and gray wolves have helped hone morphological and behavioral adaptations in each other for at least 1 million years of coevolution (Nelson and Mech 1981). Herein, the objectives of the Minnesota Department of Natural Resources (MNDNR) are to (1) provide a general historical perspective for gray wolves and white-tailed deer in Minnesota and (2) highlight research findings specifically concerning wolf-deer interactions and aspects of each species' ecology pertinent to those interactions. WOLF AND DEER POPULATIONS Historical Background White-tailed deer.--White-tailed deer range throughout Minnesota and are considered the state's most valued big game animal. Recent estimates of the state's pre-fawning (spring) population have exceeded 600,000 deer, with fall estimates reaching as high as 1 million deer (MNDNR 1990). White-tailed deer were not always so abundant. Prior to European settlement (before 1860), deer were most common in the hardwood forests of central and southeastern Minnesota and were relatively rare in the pine (Pinus spp.) forests east and north of the Mississippi River (MNDNR 1990). Two principal factors positively impacting the growth of the deer population have been (1) alterations of their habitat by timber harvesting, fire, and agriculture; and (2) increasingly intense management directly by manipulation of the population's sex and age composition (through hunting) and indirectly via habitat management programs. The first big game law affecting deer in Minnesota was established in 1858, although enforcement was likely difficult (MNDNR 1974). Initial management attempts at estimating the annual deer harvest and written records date back to 1918. The methodology for estimating annual harvests and hunter success rate gradually improved, so that by 1956 a statistically valid sampling method was adopted (MNDNR 1974). Over the years, deer population trends have varied around the state. By 1920, deer had become fairly common in northern Minnesota, but they were nearly eliminated from the prairies due to farming and subsistence hunting (MNDNR 1990). Since the 1950s, documented fluctuations in the state's deer populations have been attributable to changes in winter severity, habitat, hunting, and predation primarily by wolves. A pronounced decline in deer numbers and the harvest in northern Minnesota occurred in response to a series of severe winters, over-harvests (either sex seasons), and overly mature habitat during the late 1960s and early 1970s (MNDNR 1974, 1990; Mech and Karns 1977). Deer populations across eastern North America similarly experienced dramatic decreases, irrespective of whether the deer were impacted by wolf predation or hunting (Voigt 1990). Minnesota's 1971 deer hunting season was closed, a practice which had been exercised intermittently by the state in the past. Minnesota was divided into Deer Management Units and Permit Areas (subunits) following the closed season of 1971. The Permit Areas allow a more refined level of local deer management, specifically with respect to setting goals and managing the annual harvest. Further, the MNDNR changed from earlier eithersex hunting to a limited harvest of females. From 1976 to 1995, the state's deer population increased rather steadily with record harvests being documented (MNDNR 1974, 1997). Relatively high losses of deer were observed in the state's northern forests during the historically severe winter of 1995-96, and evidence indicated low reproductive success during the subsequent spring (DelGiudice 1996, 1998; Nelson, personal communication). Mortality rate was markedly lower during winter 1996-97, despite weather conditions nearly as severe (G. D. DelGiudice, unpublished data; M. E. Nelson, personal communication). Followed by a historically mild winter (1997-98) and an expect ed relatively high reproductive success during the 1998 fawning season, initiation of recovery of the northern deer population is imminent. Timber Wolves.--Today, wolves do not range statewide, and their numbers have followed a trend different from that of deer. Further, the factors directly responsible for the changes in their numbers are somewhat different from those affecting deer. Historically, wolves occupied all habitats of Minnesota, ranging from prairies to forests (Young and Goldman 1944, Mech and Frenzel 1971). As early as 1849, the state began paying bounties for wolves, and by 1900, wolves were rare in southern and western Minnesota (Herrick 1892, Surber 1932, Fuller et al. 1992). As both forests and wolves were harvested aggressively, wolf numbers and range continued to diminish, despite increases in some of the local deer populations. By 1941, the highest wolf densities (estimated 39 wolves/1,000 km 2 [386 mi2], Olson 1938) occurred in the northern third of Minnesota (Surber 1932, Fuller et al. 1992). Based on the field research of Stenlund (1955) in northeastern Minnesota and MNDNR records of bounty rates in northwestern Minnesota, Fuller et al. (1992) estimated that during 1950-1952, the wolf population was roughly 430-636 for the 31,080 km2 (12,000 mi2) of "major" wolf range and 450-700 wolves when areas of occasional wolf observations were included. The average number of wolves submitted annually for bounty was 253, not including individuals collected by MNDNR personnel (MNDNR files, Fuller et al. 1992). The average annual number of wolves submitted for bounty decreased to 186-189 during 1953-1965, and a gross estimate of the population was 350-700 wolves, restricted primarily to the northeastern portion of the state (MNDNR files, Cahalane 1964). Bounties for wolves and all predator species were discontinued in 1965, and in 1966, wolves were classified by the federal government as an "endangered species," but legal harvests continued through 1973. Harvests included wolves trapped by a MNDNR Directed Predator Control Program which concentrated efforts in areas of livestock depredation (Fritts 1982, Fuller et al. 1992). By 1970, the estimate of wolves was 750, changing little since the 1953-1965 estimate (Leirfallom 1970, Nelson 1971, Fuller et al. 1992), and they were assigned full protection in the Superior National Forest (Van Ballenberghe 1974). In 1974, wolves were fully protected statewide by the Endangered Species Act of 1973. Subsequent estimates of wolf numbers were 500-1,000; 1,000-1,200; and 1,235 in 1973, 1976, and 1979, respectively (Mech and Rausch 1975, Bailey 1978, Berg and Kuehn 1982). The latter estimate was associated with a major or "primary" wolf range of 36,500 km2 (14,093 mi2) and a "peripheral" range of 55,600 km2 (21,467 mi2) (Berg and Kuehn 1982). Wolves in Minnesota were reclassified as "threatened" in 1978 (U.S. Fish and Wildlife Service 1978 from Fuller et al. 1992), and occupied wolf range had increased to 57,050 km 2 (22,027 mi2), nearly double the estimate for 1950-1952 (Fuller et al. 1992). Most wolves occurred in areas where there was less than 0.70 km roads/km2 and less than 4 humans/km2 (Mech et al. 1988b). An extensive survey during winter 1988-89, estimated an occupied wolf range of at least 53,100 km 2 (20,502 mi2), excluding approximately 8,000 km2 (3,089 mi2) within this contiguous range (Fuller et al. 1992). These authors identified an additional 11,500 km2 (4,440 mi2) of potential range. Us ing two different approaches, Fuller et al. (1992) estimated the population of winter 1988-89 at 1,521 and 1,710 wolves. The survey of 1988-89 was repeated during winter 1997-98 with far more observers and technology, and although the data are not yet analyzed, preliminary evidence indicates that wolf numbers have increased and range expanded (Paul 1997). Wolf Territories, Deer Movements and Distribution, and Predation RisK Olson (1938), Murie (1944), and Young and Goldman (1944) pioneered ecological study of wolves and their prey. The earliest studies of the relationship between wolves and white-tailed deer in Minnesota and Wisconsin were conducted by Olson (1938), Thompson (1952), and Stenlund (1955). Including and since that time, most of the research information concerning wolf-deer interactions has been generated from studies conducted in what is considered primary wolf range of northern Minnesota. The most long-term and diverse ecological data have been generated from research conducted in northeastern Minnesota (Mech and Frenzel 1971; Hoskinson and Mech 1976; Mech 1977a,b,c, 1994; Mech and Karns 1977; Rogers et al. 1980; Nelson and Mech 1981, 1986a,b, 1991; DelGiudice et al. 1991; Kunkel and Mech 1994), followed by northcentral (Berg and Kuehn 1980, 1982; Fuller and Snow 1988; Fuller 1989, 1990, 1991; DelGiudice 1995, 1996, 1998; DelGiudice and Riggs 1996) and northwestern Minnesota (Fritts and Mech 1981). Overall, these regional studies have illuminated both strong similarities, as well as quantitative variation of different aspects of wolf and white-tailed deer ecology and the interactions of these 2 species. The territorial nature of wolves has been thoroughly documented. In Minnesota, their territories range from 52 to 555 km2 (20-214 mi2) and remain relatively stable throughout the year with no distinctly different winter and summer ranges (Mech 1972, 1973, 1974, 1977a; Van Ballenberghe et al. 1975; Fritts and Mech 1981; Fuller 1989). The wide variation in territory sizes is in part linked to prey density. Wolf pack territories are smaller as deer become more numerous (Fritts and Mech 1981, Fuller 1989). Further, territory size of a newly formed pack may be large, but decreases have been observed once such packs become established in an area (Fritts and Mech 1981). Wolf packs tend to be discriminatory in the use of their territory, using some portions more than others; this appears to be related to differences in physiography and deer densities, as well as proximity of neighboring wolf pack territories (Ho skinson and Mech 1976; Mech 1977a,b,c; 1994; Fritts and Mech 1981). With respect to the latter, evidence has indicated that boundaries of territories of neighboring packs commonly overlap by an estimated 6.4 km (4.0 mi) (3.2 km inside and outside estimated territorial borders, Mech 1994). This area, referred to as a "buffer zone," may be contested by neighboring packs, and consequently receives less use by the wolves of these packs (Peters and Mech 1975; Hoskinson and Mech 1976; Mech 1977a,c, 1994; Fritts and Mech 1981; Nelson and Mech 1981). Indeed, the inter-pack strife associated with buffer zones may be so serious that a wolf's "risk of a fatal encounter" with neighboring packs may be notably increased (Mech 1994). In contrast to wolves, most white-tailed deer in northern Minnesota, up to 85%, are seasonal migrators with a high fidelity for distinctly different winter and spring-summer-fall home ranges (Hoskinson and Mech 1976; Nelson and Mech 1981,1987; Fuller 1990; DelGiudice 1993). Observed spring and fall migration distances of deer have ranged from 3 to 37 km (2-23 mi), with most being 8-16 km (5-10 mi) (Hoskinson and Mech 1976, Nelson and Mech 1981, DelGiudice 1993). Long-term research collected in northeastern Minnesota has shown that deer are particularly vulnerable to predation by wolves during fall migration to winter yards (Nelson and Mech 1991). Duration of this migration is typically brief (average of 4.5 days), but daily mortality of deer has been disproportionately high. Specifically, study deer spent approximately 1% of the year migrating, but deer killed by wolves during this time comprised 21% of all kills. The reasons for this increased vulnerability are n ot fully understood, but changing weather conditions, less familiar terrain, and other factors have been proposed as contributing factors that warrant further study. Aggregations of deer and extensive trail networks once in the winter yard assist them in their defense against wolf predation (Nelson and Mech 1981, 1991). Higher winter mortality for male and female yearlings and adult females that did not yard compared to those that did has been reported (Fritts and Mech 1981, Nelson and Mech 1981). Further, several studies have observed a preponderence of deer-kills by wolves along the edges of wintering yards and more adult male than female deer killed by wolves during winter, which may be related to differences in the spatial distribution of the adults of the sexes in the yards (Stenlund 1955, Pimlott et al. 1969, Mech and Frenzel 1971, Mech and Karns 1977, Fritts and Mech 1981, Nelson and Mech 1986a). All of this suggests that yarding is, at least in part, an antipredato ry behavior not simply a strategy strictly linked to nutritional and thermal benefits. Size of seasonal home ranges of deer can vary markedly (Kohn and Mooty 1971, Hoskinson and Mech 1976, Nelson and Mech 1981). Winter home ranges are the smallest with reported average sizes ranging between 18 and 36 ha (44-89 ac) for all sex and age classes (Nelson and Mech 1981; G. D. DelGiudice, unpublished data). Following spring migration, widely dispersed summer home ranges have averaged 83, 109, and 319 ha (205, 269, and 788 ac) for adult females, yearling bucks, and adult bucks, respectively (Nelson and Mech 1981). Does with fawns reduce their home range size by as much as 65% after giving birth (e.g., from 108 to 38 ha [267 to 94 ac], Nelson and Mech 1981). The wide dispersion of does prior to fawning and the reduced home range sizes during the fawning season appear to be antipredatory strategies---newborn fawns attempt to avoid predation by hiding not fleeing. In northeastern Minnesota, fall home ranges of bucks increased significantly to 225 and 749 ha (556 and 1,850 ac) for yearlings and adults; ranges of adult females tended to increase, but differences were not significant (Nelson and Mech 1981). Interestingly, preliminary evidence has suggested that the territorial buffer zones of wolf packs may have real significance for the long-term persistence of deer populations in Minnesota's forests. Studies of radio- collared deer in primary wolf range have shown that most of their seasonal home ranges were located at the boundaries of juxtaposed territories of wolf packs, where, as discussed above, inter-pack strife is greatest and wolf activity tends to be less (Peters and Mech 1975; Hoskinson and Mech 1976; Mech 1977a,b, 1994; Fritts and Mech 1981; Nelson and Mech 1981). Deer home ranges in these buffer zones may serve as "reservoirs of deer" when the combined influence of several factors (e.g., severe winters, wolf predation, hunting, maturing habitat--Mech and Karns 1977) causes the decline of a local deer population. Wolf Predation of Deer and Food Habits of Wolves Across northern Minnesota, hunter harvest and wolf predation are the primary causes of mortality of whitetailed deer (Hoskinson and Mech 1976; Nelson and Mech 1981, 1986a,b; Fuller 1990; DelGiudice 1994, 1998; DelGiudice and Riggs 1996). However, it is important to note that the relative importance of these 2 causes of mortality vary among years and local deer populations, dependent upon differences in hunter pressure, winter severity, wolf and deer densities, and sex and age composition of the deer population. It has been well documented in studies across northern Minnesota that wolves primarily kill young of the year and older deer in the population, and these individuals are often compromised by deterioration of their overall physical condition or a specific abnormality (Mech and Frenzel 1971, Fritts and Mech 1981, DelGiudice 1996, DelGiudice and Riggs 1996). In northeastern Minnesota, the average age of deer (both sexes) killed by hunters was 2.6 years compared to 4.7 years for deer killed by wolves during DecemberMarch (1966-1969) (Mech and Frenzel 1971). Similarly, in northwestern Minnesota, the average ages of deer (both sexes) killed by hunters and wolves were 3.3 and 7.6 years, respectively (Fritts and Mech 1981). In northcentral Minnesota, the median ages (half the cause-specific mortality occurred by this age) of female deer killed by hunters and wolves were 2.5 and 6.5 years, respectively (DelGiudice 1996). Wolf predation on deer is greatest during mid-late winter, coinciding with the period of poorest condition and deepest snow (Mech and Frenzel 1971, Mech 1977b, Moen and Severinghaus 1981, DelGiudice et al. 1992, DelGiudice 1998), then again during the fawning period of early summer when neonates are vulnerable prey to wolves (Pimlott et al. 1969; Mech et al. 1971; Fritts and Mech 1981; Nelson and Mech 1981, 1986a,b; Fuller 1989; Kunkel and Mech 1994) and other predators (see Fuller 1990; Dickson 1992; L. L. Rogers, unpublished data). For the period 1973-1984, Nelson and Mech (1986a) reported an annual mortality rate of radio-collared deer (both sexes) attributable to wolves of 0.16-0.19 (i.e., 16-19%), except for yearling females which experienced a rate of 0.05. In a study of wolves and deer in northcentral Minnesota (Bearville study area north of Grand Rapids), Fuller (1990) documented an annual wolf-caused mortality rate of 0.04 for radio-collared dee r >/=1.0 year old (both sexes) during the period 1981-1986. From 1991 to 1997, DelGiudice (1996) observed annual, wolf-caused mortality rates of 0.09-0.26 for radio-collared, female deer (primarily >/=1 yr old). In eastern Ontario, Kolenosky (1972) documented a wolf-caused mortality rate of 0.09-0.11 when snow depths were fairly deep. The variation of mortality rates of deer from area to area and year to year are primarily attributable to variations in deer:wolf ratios, winter severity, presence of alternate prey (e.g., moose), and the sex and age composition of the deer populations. Of note, wolf predation rates on deer appear to be higher in areas with lower deer:wolf ratios (Kolenosky 1972, Nelson and Mech 1981, Fuller 1990). Moreover, there is good evidence that pairs of wolves in a given area have higher winter kill rates of deer per wolf than packs of wolves (>/=3 members), suggesting a low optimum pack size for maximizing energy to individual members (Fritts and Mech 1981, Schmidt and Mech 1997). Severe winter weather negatively impacts survival of northern white-tailed deer either through nutritional restriction when predators are scarce or by predation where wolves or other predators are common (Severinghaus 1947; Erickson et al. 1961; Mech et al. 1971; Verme and Ozoga 1971; Nelson and Mech 1986b; DelGiudice 1996, 1998). Several studies have demonstrated that snow depth (and density) specifically has a strong, direct influence on wolf predation of deer in Minnesota and elsewhere (Pimlott et al. 1969; Mech et al. 1971; Nelson and Mech 1986b; DelGiudice 1996, 1998). In the extreme case, during severe winters of exceptionally deep snow (>/=70 cm), excessive or surplus killing of deer by wolves, characterized as multiple kills made over short distances and brief periods of time with less than complete or no consumption, has been documented (Mech et al. 1971, DelGiudice 1998). Deteriorating condition of deer as winter progresses and impedence of deer move ments by deep snow that is less supportive of deer than wolves have been implicated as primary factors contributing to this phenomenon (Mech et al. 1971, DelGiudice 1998). Surplus killing of prey under certain environmental conditions has been documented for a wide variety of predators (Kruuk 1972). Scat analyses have revealed much about the year-round food habits of wolves, including that deer are their single most important food, except during winter in the Boundary Waters Canoe Area where they rely primarily on moose (Frenzel 1974, Van Ballenberghe et al. 1975, Fritts and Mech 1981, Fuller 1989). Both in northwestern (i.e., Beltrami State Forest) and northeastern Minnesota, deer occurred in the scats of wolves more in winter than in the summer (Thompson 1952, Pimlott et al. 1969, Van Ballenberghe et al. 1975, Theberge et al. 1978, Fritts and Mech 1981); this is congruent with the evidence from monitoring causespecific mortality of radio-collared deer. Fawns are an important food item in the summer diet of wolves in the Lake Superior region (Thompson et al. 1952, Pimlott et al. 1969, Frenzel 1974, Van Ballenberghe et al. 1975, Fritts and Mech 1981, Nelson and Mech 1986a, Kunkel and Mech 1994). Indeed, evidence indicates that fawns probably account for most of the individual deer consumed by wolves during June-July, as well as a higher proportion of the overall biomass consumed (Fritts and Mech 1981). Vulnerability of fawns to predation by wolves and black bears (Ursus americanus) is at least partly related to their nutritional status, which may be directly influenced by the severity of the previous winter and its effect on prenatal nutrition (Verme 1962, Mech and Karns 1977, Mech et al. 1987, Kunkel and Mech 1994). Annual percentages of young fawns taken by wolves (and other predators) are still relatively unknown, and additional study is warranted. In some areas such as northwestern Minnesota, where deer are largely the most important year-round food for wolves, there are brief periods (April-May) when wolves rely more on moose. Certainly, moose are the second most important food item in this area during summer and winter, with wolves consuming more individuals and biomass during summer than winter (Fritts and Mech 1981). The relative importance of moose in the diet of wolves that depend primarily on deer year-round may vary, dependent upon deer densities and the degree with which factors such as winter severity and disease or parasites (e.g., winter tick infestation, cerebrospinal nematodiasis) predispose moose to predation or scavenging by wolves. Snowshoe hares (Lepus americanus) are consumed by wolves year-round, and although the number of individuals consumed may be second only to deer, the biomass contributed to the diet is rather insignificant (< 1%) (Fritts and Mech 1981, Fuller 1989). Beaver (Castor canadensis) have been found in the diet of wolves in spring and summer (Byman, unpublished thesis, Frenzel 1974, Fritts and Mech 1981, Fuller 1989); data indicate that its importance may be greater in northeastern than northwestern Minnesota. An increased consumption of beaver accompanied a decline in white-tailed deer populations in southern Ontario (Voigt et al. 1976, Theberge et al. 1978). Other mammals documented in the diet of wolves include bog lemming (Synaptomys sp.), fox squirrel (Sciurus niger), jumping mouse (Zapus hudsonius), meadow vole (Microtus pennsylvanicus), white-footed mouse (Peromyscus sp.), and woodchuck (Marmota monax) (Young and Goldman 1944, Fritts and Mech 1981). Additional miscellaneous foods have included black bear, striped skunk (Mephitis mephitis), wolf, unidentified Canis, various bird species, duck and duck egg shells, and insects. Wolves have also consumed fruits such as blueberries (Vaccinium sp.) and strawberries (Fragaria sp.) during summer and early fall (Van Ballenberghe et al. 1975, Fritts and Mech 1981). Livestock Depredation by Wolves Where they coexist, wolves will occasionally prey on livestock, including cattle (primarily calves), sheep, turkeys, swine, and goats, but generally, they occur in the wolf's diet in low amounts, both with respect to the number of individuals and biomass consumed (Fritts and Mech 1981, Fuller 1989, Fritts et al. 1992). There is substantial temporal and spatial variation concerning livestock depredation by wolves, with wolves in some areas relying on domestic prey more heavily than others in certain years (Fritts et al. 1992). Most depredations occur in northcentral and northwestern counties where farm and livestock densities are highest within wolf range (Fritts et al. 1992). Research has indicated an inverse relationship between the severity of the previous winter, and thus the vulnerability of newborn fawns, and the amount of depredation on livestock (and pets) (Mech et al. 1988a, Fritts et al. 1992). Li vestock depredation by wolves tends to be higher in some areas following a less severe winter when the nutritional status of deer fawns is better and they are less vulnerable. During the period 1975-1986, the wolf depredation control program verified an average 30 complaints per year involving an average 21 farms. However, for the shorter, more recent interval 1979-1986, these figures were 33 verified complaints and 24 farms (0.33% of farms in wolf range). During 1979-1986, average annual, verified losses of 23 cattle, 49 sheep, and 173 turkeys were reported; most complaints occurred during May-September. Numbers of animals wounded or killed peaked in May, July-August, and AugustSeptember for cattle, sheep, and turkeys, respectively (Fritts et al. 1992). The area affected by verified depredations expanded 48.7% from 38,228 km 2 (14,760 mi2) in 1975-1980 to 56,827 km2 (21,941 mi2) in 1981-1986; the expansion occurred in all directions, but north (Fritts et al. 1992). When adult and yearling wolves were removed by the contro l program, additional losses did not occur in 55% of the cases, 22% of the cases when young of the year were removed. Removal of breeding versus nonbreeding members of packs did not have any greater impact on subsequent losses of livestock. From 1987 to 1997, verified complaints of wolf depredation on livestock and the number of farms involved has tripled (Paul 1997). Specifically, in 1997, there were 109 verified complaints at 93 farms. Also during this period, the number of wolves captured and removed from the population has increased from 30-35 per year to just over 200 per year; the average has been 158 wolves captured per year during the past 5 years (Paul 1997). A state program which compensates farmers for livestock lost to wolves, paid an average $37,299 per year over the past 5 years. Greater detail of wolf depredation of livestock and the depredation control program in Minnesota can be obtained elsewhere (Fritts et al.1992, Paul 1997). Wolves and Deer---Influences at the Population Level As is evident from the above, wolf and white-tailed deer populations are intricately linked, each having a potentially strong influence on the other's population performance (i.e., survival rates, reproductive success). The specific type and strength of the influences or interactions between wolf and deer populations are a function of factors such as individual densities, sex and age structures, human-related activities (e.g., hunting, poaching, dogs, timber harvesting, road development, supplemental feeding), winter severity, presence of alternate prey, and habitat quality (Mech 1986, Mech et al. 1988b). Clearly, nutrition is the basis of this link in both directions. Wolves have a direct impact on white-tailed deer populations in being the primary cause of natural mortality, again, with the most vulnerable individuals being newborn fawns and fawns (>/=0.6 yr old) and older adults during winter (Mech and Frenzel 1971; Nelson and Mech 1986a,b; DelGiudice and Riggs 1996; DelGiudice 1996). Thus far, there is little evidence to show that wolves are a limiting factor on deer, or alone, have caused population declines (Voigt 1990). However, in combination with severe winter conditions, overly mature habitat, and/or human exploitation, wolf predation has contributed to locally declining deer populations such as occurred in the central Superior National Forest in the late 1960s-early 1970s and elsewhere during winter 1995-96 (Mech and Karns 1977; DelGiudice 1996, 1998). In the central Superior National Forest and areas further north and northeast, wolves may be affecting the recovery of the deer population (Nelson and Mech 1981). Wolf predation may represent compensatory or additive mortality. For example, if wolves kill deer during winter that ultimately would have died of starvation, the predation is compensatory mortality. However, if in very severe winters, wolves are able to kill some relatively healthy deer that would have survived t he winter nutritional stress, then that predation is additive. Generally, based upon ages of deer killed by hunters and wolves (discussed earlier), it is believed that most deer deaths from wolves in winter are a combination of compensatory and additive mortality, whereas, the hunter-kill tends to be more additive (i.e., prime age classes). A recent study reports that during 1975-1997, numbers of wolves in the east-central Superior National Forest accounted for only 23-38% of the inter-year variation in buck harvest for areas in and near the 2,060 km 2 (795 mi2) wolf census area (Mech and Nelson 1998). Year-to-year analysis of annual changes in numbers of wolves and harvested bucks revealed inconsistent associations. Increases in annual deer harvests in much of Minnesota wolf range, even as it expanded, indicates that generally, the wolves' effect on deer harvests has been minimal. Presently, too little is known about t he mortality of deer neonates to speculate whether its effects are compensatory or additive; however, preliminary evidence has indicated that some newborns killed by wolves and bears may be compromised nutritionally (Kunkel and Mech 1994). Because deer are the wolf's primary food across most of their range in Minnesota, it is easy to understand the eventual, nutritional consequences for wolves of a local deer decline. The estimated minimum maintenance requirement of food for free-ranging wolves is 1.7 kg per day of deer-equivalent biomass (assuming an average 54.5 kg of food provided from an adult deer [portions of this can be from beaver, snowshoe hares, moose, etc...]) (Mech 1970). Estimated daily consumption rates per wolf have ranged from 0.2 to 7.3 kg (Mech 1966, Mech and Frenzel 1971, Kolenosky 1972, Kuyt 1972, Fritts and Mech 1981, Fuller 1989). It has been estimated that a wolf requires the equivalent of 15-20 adult-sized deer per year (Mech 1971, Kolenosky 1972, Fuller 1989). However, low deer numbers; fewer old, vulnerable deer (i.e., a young age structure); or a mild winter in a given area will influence the number of deer or biomass each wolf actually consumes. Consequence s of submaintenance consumption and nutritional stress for wolves during the breeding (February-March) and denning seasons (late April-May) can be increased adult mortality (undernutrition, more trespassing and inter-pack strife), small litters, and decreased pup survival. In the past in the central Superior National Forest, the collective effect of such dynamics involved a local wolf population that decreased 50% in 4 years (Seal et al. 1975; Van Ballenberghe and Mech 1975; Mech 1977a,b, 1986). This type of predator-prey relationship---a predator population contributing to the decline of its prey population, followed by a lag period and subsequent decline in the predator population---is classical, but it is not simple. As mentioned above, many other factors will interact, influence the specific dynamics of the relationship (what changes occur and the magnitude of those changes) and determine the temporal scale over which they occur. LITERATURE CITED Bailey, R., editor. 1975. Recovery plan for the eastern timber wolf. U.S. Fish and Wildlife Service, Washington, D.C. 79pp. Berg, W. E., and D. W. Kuehn. 1980. A study of the timber wolf population on the Chippewa National Forest, Minnesota. Minnesota Department of Natural Resources. Wildlife Research Quarterly 40:1-16. Berg, W. E., and D. W. Kuehn. 1982. Ecology of wolves in northcentral Minnesota. Pages 4-11 in F. H. Harrington and P. C. Paquet, eds. Wolves: a worldwide perspective of their behavior, ecology, and conservation. Noyes Publ., Park Ridge, N. J. Cahalane, V. H. 1964. A preliminary study of distribution and numbers of cougar, grizzly, and wolf in North America. New York Zoological Society, New York, N. Y. 12pp. DelGiudice, G. D. 1993. Assessing the relationship of conifer thermal cover to winter distribution, movements, and survival of female white-tailed deer in northcentral Minnesota. Pages 25-44 in B. Joselyn, ed. Summaries of wildlife research findings. Minnesota Department of Natural Resources, St. Paul. 177pp. DelGiudice, G. D. 1994. Assessing the relationship of conifer thermal cover to winter distribution, movements, and survival of female white-tailed deer in northcentral Minnesota. Pages 45-63 in B. Joselyn, ed. Summaries of wildlife research findings. Minnesota Department of Natural Resources, St. Paul. 296pp. DelGiudice, G. D. 1995. Assessing the relationship of conifer thermal cover to winter distribution, movements, and survival of female white-tailed deer in northcentral Minnesota. Pages 113-133 in B. Joselyn, ed. Summaries of wildlife research findings. Minnesota Department of Natural Resources, St. Paul. 297pp. DelGiudice, G. D. 1996. Assessing the relationship of conifer thermal cover to winter distribution, movements, and survival of female white-tailed deer in northcentral Minnesota. Pages 59-81 in B. Joselyn, ed. Summaries of wildlife research findings. Minnesota Department of Natural Resources, St. Paul. 256pp. DelGiudice, G. D. 1998. Surplus killing of white-tailed deer by wolves in northcentral Minnesota. Journal of Mammalogy 79:227-235. DelGiudice, G. D., L. D. Mech, K. E. Kunkel, E. M. Gese, and U. S. Seal. 1992. Seasonal patterns of weight, hematology, and serum characteristics of free-ranging female white- tailed deer in Minnesota. Canadian Journal of Zoology 70:974-983. DelGiudice, G. D., L. D. Mech, and U. S. Seal. 1991. Gray wolf density and its association with weights and hematology of pups from 1970 to 1988. Journal of Wildlife Diseases 27:630- 636. DelGiudice, G. D., and M. R. Riggs. 1996. Long-term research on the white-tailed deer/conifer thermal cover relationship: aligning expectations with reality. Transactions of the North American Wildlife and Natural Resources Conference 61:416-428. Dickson, T. 1992. Tooth for a tooth. Minnesota Volunteer. November-December Issue:6-19. Erickson, A. B., V. E. Gunvalson, M. H. Stenlund, D. W. Burcalow, and L. H. Blankenship. 1961. The whitetailed deer of Minnesota. Minnesota Department of Conservation Technical Bulletin 5:1-64. Frenzel, L. D. 1974. Occurrence of moose in food of wolves as revealed by scat analysis: a review of North American studies. Naturaliste Canadiene 101:467-479. Fritts, S. H. 1982. Wolf depredation on livestock in Minnesota. U.S. Fish and Wildlife Service Resource Publication 145. 11pp. Fritts, S. H., and L. D. Mech. 1981. Dynamics, movements, and feeding ecology of a newly protected wolf population in northwestern Minnesota. Wildlife Monographs No. 80. 79pp. Fritts, S. H., W. J. Paul, L. D. Mech, and D. P. Scott. 1992. Trends and management of wolf- livestock conflicts in Minnesota. U. S. Fish and Wildlife Service, Resource Publication 181. 27pp. Fuller, T. K. 1989. Population dynamics of wolves in northcentral Minnesota. Wildlife Monographs No. 105. 41pp. Fuller, T. K. 1990. Dynamics of a declining white-tailed deer population in northcentral Minnesota. Wildlife Monographs No. 110. 37pp. Fuller, T. K. 1991. Effect of snow depth on wolf activity and prey selection in northcentral Minnesota. Canadian Journal of Zoology 69:283-287. Fuller, T. K., W. E. Berg, G. L. Radde, M. S. Lenarz, and G. B. Joselyn. 1992. A history and current estimate of wolf distribution and numbers in Minnesota. Wildlife Society Bulletin 20: 42-55. Fuller, T. K., and W. J. Snow. 1988. Estimating wolf densities from radio telemetry data. Wildlife Society Bulletin 16:367-370. Herrick, C. L. 1892. The mammals of Minnesota. Geol. Nat. Hist. Serv. Minnesota Bulletin 7. 299pp. Hoskinson, R. L., and L. D. Mech. 1976. White-tailed deer migration and its role in wolf predation. The Journal of Wildlife Management 40:429-441. Kohn, B. E., and J. J. Mooty. 1971. Summer habitat of white-tailed deer in northcentral Minnesota. The Journal of Wildlife Management 35:476-487. Kolenosky, G. B. 1972. Wolf predation on wintering deer in east-central Ontario. The Journal of Wildlife Management 36:357-369. Kruuk, H. 1972. Surplus killing by carnivores. Journal of Zoology (London). 166:233-244. Kulp, J. L. 1961. The geologic time scale. Science 133:1105-1114. Kunkel, K. E., and L. D. Mech. 1994. Wolf and bear predation on white-tailed deer fawns in northeastern Minnesota. Canadian Journal of Zoology 72:1557-1565. Kuyt, E. 1972. Food habits of wolves on barren-ground caribou range. Canadian Wildlife Service Report Series 21:1-36. Leirfallom, J. 1970. Wolf management in Minnesota. Pages 9-15 in S. E. Jorgenson, C. E. Faulkner, and L. D. Mech, eds. Proceedings of symposium on wolf management in selected areas of NorthAmerica. U.S. Fish and Wildlife Service, Twin Cities, Minnesota. Matthew, W. D. 1930. The phylogeny of dogs. Journal of Mammalogy 11:117-138. Mech, L. D. 1966. The wolves of Isle Royale. National Parks Fauna Series 7. U.S. Governement Printing Office, Washington, D.C. Mech, L. D. 1970. The wolf. Ecology and behavior of an endangered species. University of Minnesota Press, Minneapolis. 384pp. Mech, L. D. 1971. Wolves, coyotes, and dogs. Pages 19-22 in M. M. Nelson, ed. The white- tailed deer in Minnesota. Proceedings of symposium of the Minnesota Chapter of the Wildlife Society, Section of Game, Minnesota Department of Natural Resources, St. Paul. Mech, L. D. 1972. Spacing and possible mechanisms of population regulation in wolves. American Zoologist 12:642. Mech, L. D. 1973. Wolf numbers in the Superior National Forest of Minnesota. U.S.D.A. Forest Service Research Paper NC-97. North Central Forest Experiment Station, St. Paul, Minnesota. 10pp. Mech, L. D. 1974. Current techniques in the study of elusive wilderness carnivores. Proceedings of the International Congress of Game Biologists 11:315-322. Mech, L. D. 1977a. Population trend and winter deer consumption in a Minnesota wolf pack. Pages 55-83 in R. L. Phillips and C. Jonkel, eds. Proceedings 1975 predator symposium, Montana Forest Conservation Experiment Station Bulletin, University of Montana, Missoula. Mech, L. D. 1977b. Productivity, mortality, and population trends of wolves in northeastern Minnesota. Journal of Mammalogy 58:559-574. Mech, L. D. 1977c. Wolf-pack buffer zones as prey reservoirs. Science 198:320-321. Mech, L. D. 1986. Wolf population in the central Superior National Forest, 1967-1985. U.S.D.A. Forest Service Research Paper NC-270. North Central Forest Experiment Station, St. Paul, Minnesota. 6pp. Mech, L. D. 1994. Buffer zones of territories of gray wolves as regions of intraspecific strife. Journal of Mammalogy 75:199-202. Mech, L. D., and L. D. Frenzel, Jr. 1971. An analysis of the age, sex, and condition of deer killed by wolves in northeastern Minnesota. Pages 35-51 in L. D. Mech and L. D. Frenzel, Jr., eds. Ecological studies of the timber wolf in northeastern Minnesota. U.S.D.A. Forest Service Research Paper NC-52. North Central Forest Experiment Station, St. Paul, Minnesota. Mech, L. D., L. D. Frenzel, Jr., and P. D. Karns. 1971. The effect of snow conditions on the vulnerability of white-tailed deer to predation. Pages 51-59 in L. D. Mech and L. D. Frenzel, Jr., eds. Ecological studies of the timber wolf in northeastern Minnesota. U.S.D.A. Forest Service Research Paper NC-52. North Central Forest Experiment Station, St. Paul, Minnesota. Mech, L. D., S. H. Fritts, and W. J. Paul. 1988a. Relationship between winter severity and wolf depredation of domestic animals in Minnesota. Wildlife Society Bulletin 16:269-272. Mech, L. D., S. H. Fritts, G. L. Radde, and W. J. Paul. 1988b. Wolf distribution in Minnesota relative to road density. Wildlife Society Bulletin 16:85-87. Mech, L. D., and P. D. Karns. 1977. Role of the wolf in a deer decline in the Superior National Forest. U.S.D.A. Forest Service Research Paper NC-148. North Central Forest Experiment Station, St. Paul, Minnesota. 23pp . Mech, L. D., R. E. McRoberts, R. O. Peterson, and R. E. Page. 1987. Relationships of deer and moose populations to previous winter's snow. Journal of Animal Ecology 56:615-627. Mech, L. D., and M. E. Nelson. 1998. Do wolves affect deer harvesting in Minnesota? The Journal of Wildlife Management. Submitted. Mech, L. D., and R. A. Rausch. 1975. The status of the wolf in the United States, 1973. Pages 83-88 in D. H. Pimlott, ed. Wolves. International Union for the Conservation of Nature and Natural Resources. Publ. New Ser., Supplemental Paper 43. Morges, Switzerland. Minnesota Department of Natural Resources. 1974. Minnesota game population statistics, harvests, and hunting regulations. St. Paul. Minnesota Department of Natural Resources. 1990. White-tailed deer. Pages 18-1 to 18-15 in Planning for Minnesota fish, wildlife, and native plant resources. St. Paul. (Public Review Draft). Minnesota Department of Natural Resources. 1997. Minnesota deer harvest report. St. Paul. 39pp. Moen, A. N., and C. W. Severinghaus. 1981. The annual weight cycle and survival of white- tailed deer in New York. N.Y. Fish and Game Journal 28:162-177. Murie, A. 1944. The wolves of Mount McKinley. U.S. National Park Service Fauna Series 5. 238pp. Nelson, M. E., and L. D. Mech. 1981. Deer social organization and wolf predation in northeastern Minnesota. Wildlife Monographs No. 77.53pp. Nelson, M. E., and L. D. Mech. 1986a. Mortality of white-tailed deer in northeastern Minnesota. The Journal of Wildlife Management 50:691-698. Nelson, M. E., and L. D. Mech. 1986b. Relationship between snow depth and gray wolf predation on whitetailed deer. The Journal of Wildlife Management 50:471-474. Nelson, M. E., and L. D. Mech. 1987. Demes within a northeastern Minnesota deer population. Pages 27-40 in B. D. Chepko-Sade and Z. Halpin, eds. Mammalian dispersal patterns. University of Chicago Press, Chicago. Nelson, M. E., and L. D. Mech. 1991. Wolf predation risk associated with white-tailed deer movements. Canadian Journal of Zoology 69:2696-2699. Nelson, M. M. 1971. Predator management with emphasis on the timber wolf. Pages 68-77 in M. M. Nelson, ed. Proceedings of symposium on the white-tailed deer in Minnesota. Minnesota Department of Natural Resources, St. Paul. Olson, S. F. 1938. A study in predatory relationships with particular reference to the wolf. Science Monthly 46:323-336. Paul,W. J. 1997. U.S. Department of Agriculture wolf depredation control work in Minnesota, 1997--program summary. Grand Rapids, Minnesota. Peters, R. P., and L. D. Mech. 1975. Scent-marking in wolves. American Scientist 63:628-637. Pimlott, D. H., J. A. Shannon, and G. B. Kolenosky. 1969. The ecology of the timber wolf in Algonquin Provincial Park. Ontario Dep. Lands and Forests Research Paper (Wildlife) 87. 92pp. Rogers, L. L., L. D. Mech, D. K. Dawson, J. M. Peek, and M. Korb. 1980. Deer distribution in relation to wolf pack territory edges. The Journal of Wildlife Management 44:253-258. Schmidt, P. A., and L. D. Mech. 1997. Pack size and food acquisition. American Naturalist 150:513-517. Seal, U. S., L. D. Mech, and V. Van Ballenberghe. 1975. Blood analyses of wolf pups and their ecological and metabolic interpretations. Journal of Mammalogy 56:64-75. Severinghaus, C. W. 1947. Relationship of weather to winter mortality and population levels among deer in the Adirondack region of New York. Transactions of the North American Wildlife Conference 12:212-223. Stenlund, M. H. 1955. A field study of the timber wolf (Canis lupus) on the Superior National Forest, Minnesota Department of Conservation Technical Bulletin 4. St. Paul. Surber, T. 1932. The mammals of Minnesota. Minnesota Department of Conservation, St. Paul. 81pp. Therberge, J. B., S. M. Oosenbrug, and D. H. Pimlott. 1978. Site and seasonal variation in foods of wolves, Algonquin Park, Ontario. Canadian Field-Naturalist 92:91-94. Thompson, D. Q. 1952. Travel, range, and food habits of timber wolves in Wisconsin. Journal of Mammalogy 33:420-442. U.S. Fish and Wildlife Service. 1978. Title 50. Wildlife and fisheries. Federal Register 43 (47):9607-9615. Van Ballenberghe, V. 1974. Wolf management in Minnesota: an endangered species case history. Transactions North American Wildlife and Natural Resources Conference 39:313- 322. Van Ballenberghe, V., A. W. Erickson, and D. Byman. 1975. Ecology of the timber wolf in northeastern Minnesota. Wildlife Monographs No. 43. 43pp. Van Ballenberghe, V., and L. D. Mech. 1975. Weights, growth, and survival of timber pups in Minnesota. Journal of Mammalogy 56:44-63. Verme, L. J. 1962. Mortality of white-tailed deer fawns in relation to nutrition. Pages 15- 38 in Proceedings of the First white-tailed deer disease symposium. University of Georgia, Athens. Verme, L. J., and J. J. Ozoga. 1971. The influence of winter weather on white-tailed deer in upper Michigan. Pages 16-28 in A. O. Haugen, ed. Proceedings of Snow and Ice Symposium, Ames, Iowa. Voigt, D. R. 1990. White-tailed deer in Ontario. Background of a policy. (Draft). Ontario Ministry of Natural Resources. 106pp. Voigt, D. R., G. B. Kolenosky, and D. H. Pimlott. 1976. Changes in summer foods of wolves in central Ontario. The Journal of Wildlife Management 40:663-668. Young, S. P., and E. A. Goldman. 1944. The wolves of North America. American Wildlife Institute, Washington, D. C. 636pp. Grade 9 Academic Science – Ecology Wolves – Yellowstone and Isle Royale Smith, D.W., R.O. Peterson and D.B. Houston. 2003. Yellowstone after Wolves. Journal of Bioscience. American Institute of Biological Sciences. http://www.wolf.org/wolves/learn/intermed/inter_mgmt/ystone_wolves.asp Abstract: With gray wolves restored to Yellowstone National Park, this ecosystem once again supports the full native array of large ungulates and the attendant large carnivores. We consider the possible ecological implications of wolf restoration in the context of another national park, Isle Royale, where wolves restored themselves a half-century ago. At Isle Royale, with relatively few resident mammals, wolves completely eliminated coyotes and went on to control moose population dynamics, with implications for forest growth and composition. At Yellowstone, we predict that, to a degree, wolf restoration will have similar effects, reducing elk and coyote density. As at Isle Royale, Yellowstone plant communities will be affected, as well as mesocarnivores, but presently there is great uncertainty. At Yellowstone, ecosystem response to the arrival of the wolf will take decades to unfold, and we argue that comprehensive ecological research and monitoring should be an essential long-term component of the management of Yellowstone National Park. Reintroduction of gray wolves to Yellowstone National Park surely ranks, symbolically and ecologically, as one of the most important overt acts of wildlife conservation in the 20th century. Once again, Yellowstone harbors all native species of large carnivores - grizzly and black bears, mountain lions, and wolves. Before wolf reintroduction, there was a concerted effort to predict the ecological effects of wolves in Yellowstone (Cook 1993). Has reality, so far, met expectations? And has what we learned in Isle Royale National Park, where wolves introduced themselves over 50 years ago, relevance for Yellowstone in the future? Gray wolves were restored to Yellowstone National Park during 1995-96, with the release of 31 wolves captured in western Canada (Bangs and Fritts 1996, Phillips and Smith 1996). In the 7 years following their initial release wolves have recolonized the 8991-km2 park and several adjacent portions of the 72,800 km2 "greater Yellowstone ecosystem (GYE)." We use initial studies and field observations to determine to what extent wolves may have already begun to restructure the Yellowstone ecosystem. Although we consider wolves park-wide, we focus on the 1,530 km2 "northern Yellowstone winter range," an area dominated by steppe and shrub steppe vegetation that supports 7 species of native ungulates -- elk, bison, mule deer, white-tailed deer, moose, pronghorn antelope, and bighorn sheep, one non-native ungulate --mountain goat, and 5 species of native large carnivores -- gray wolf, coyote, grizzly bear, black bear, cougar. Only about 65% of the northern range is inside the park; the remaining 35% occurs on public and private lands north of the park along the Yellowstone River (Lemke et al 1998). Since many of these wildlife species are hunted outside the park, we include technological humans as additional, formidable, predators in the system. While the National Park Service manages Yellowstone with an overall goal of minimal human intervention allowing natural ecological processes to prevail inside park boundaries—its wildlife populations may be profoundly altered by human actions, including hunting, outside the park. Simplicity and Complexity - Isle Royale and Yellowstone We find it useful to contrast the Yellowstone system with that of Isle Royale National Park, a less complex ecosystem renowned for long-term studies of the interaction of gray wolves with moose conducted there (Peterson 1995, Peterson et al. 1998). Amid the complexity of Yellowstone, where might we expect to find the ecological footprints of wolves, and where might science make its greatest gains? We anticipate that long-term studies similar to those of Isle Royale will be required to understand the effects of wolves in Yellowstone. We could have picked other parks - Riding Mountain in Manitoba or Denali in Alaska which are both multi-carnivore/prey systems like Yellowstone- but long term data (turn of the century to present) on willdife population sizes from these other areas were lacking, and we do not have intimmate experience with these parks. Often what is important is subtle and detailed which can be the difference between an informed conclusion and one that is not. Where appropriate we make comparisons to other wolf-prey systems. Isle Royale and Yellowstone provide opposite extremes in faunal and food web complexity. Isle Royale is a closed system with fewer species (one-third the species found on the adjacent mainland), and Yellowstone is an open system with greater diversity of both predators and prey (Fig. 1). Thus, Isle Royale should be more amenable to scientific scrutiny, with clearer cause-and-effect relationships among a few key species, a good starting point and example for interpreting Yellowstone. - There are surprising parallels in the history of Isle Royale and Yellowstone during the past century, particularly in concerns raised over too many ungulates and their effects on their habitat. During a wolf-free period, both ecosystems saw ungulates increase to levels that alarmed some knowledgeable observers, and coyotes were numerous in both areas. It is not only ecology that is complex at Yellowstone. Its bureaucratic history as the nation's "first" national park (Haines 1977) is long and rich. Management of Yellowstone's wildlife, particularly on the northern range, has a history of concern and controversy dating from the establishment of the park in 1872 (Pritchard 1999). Early on, extirpation of many native species was feared because of intense hide and market hunting. Understandably, this period was followed by one of progressively increased husbandry of native ungulates, which eventually involved winter-feeding and predator control. Gray wolves were effectively eliminated by the 1930's (Weaver 1978). During the extended drought of the 1930s some ungulate species, particularly elk, were considered to be "overabundant," and "range deterioration" became an issue. This led, in turn, to intense and highly controversial reductions of elk, bison, and pronghorn populations by field shooting and trapping, aimed at testing the effects of reduced ungulate densities on vegetation conditions. By the late 1960s elk numbers had been reduced by perhaps 75%, to around 4,000 animals (Houston 1982). In 1969, a moratorium on reductions was instituted, as attempts were made to rely more upon natural regulation of ungulate numbers within the park and to restore hunting opportunities outside (reductions inside the park had essentially eliminated elk hunting outside). Efforts to rely on more natural processes have, in one sense, culminated in restoration of the wolf. This brief outline of management history is treated in detail by Meagher (1973), Houston (1982), and Pritchard (1999). Like Yellowstone, Isle Royale had a wolf-free era that also resulted in an over-abundant moose population (Allen 1979). Instead of artificial reductions to control moose, the Park Service unsuccessfully tried to reintroduce zoo-raised wolves in 1952 (Allen 1979). But unlike Yellowstone, wolves reintroduced themselves to Isle Royale in the late 1940s by crossing the ice of Lake Superior (Allen 1979). What did the arrival of the wolf mean for the Isle Royale ecosystem? While scientific uncertainty continues on the relative roles of bottom-up (nutrition/vegetation) and top-down (wolf predation) influences on moose population dynamics (Messier 1994, Peterson 1995), the historic chronology of moose numbers indicates that wolf predation tends to cap moose density (Fig. 2). The peak in moose numbers in the early 1970s ended when severe winters affected vulnerability (Peterson 1977), and the resulting increase in wolves kept the moose population low for many years. More wolves indirectly allowed forest recovery via less moose browsing (topdown; McLaren and Peterson 1994). However, when wolves crashed in the 1980s, from 50 to 14 in two years, and were limited because of a canine parvovirus, an accidentally and human introduced disease (Peterson 1995), moose increased until catastrophic starvation hit in 1996 (one of the most severe winters on record; Peterson et al. 1998). The rise and fall of Isle Royale's wolf population can be read in growth rings of balsam fir trees - trees flourish when wolf numbers increase and moose are reduced (McLaren and Peterson 1994, McLaren 1996). The relative abundance of coniferous and deciduous trees, strongly influenced by moose browsing, further affects litter composition and nutrient cycling in the soil, so the ripple effect beginning with the arrival of wolves extends far and wide (Pastor et al. 1993). But it is not that simple. On one-third of Isle Royale, fir trees are able to escape moose browsing (because of thick, high density stands) and grow into the canopy, but on the majority of the island, balsam fir trees are unable to grow out of the reach of moose (McLaren and Janke 1996). Hence, moose remain a powerful force shaping forest succession, even with intense wolf predation. Variations in soil types, disturbance history (fire and wind) and light intensity complicate a system that, in comparison to Yellowstone, is easily understood. Even after a century with moose, the forest of Isle Royale has not yet reached equilibrium. One needs a long-term perspective and study to understand completely the dynamics of long-lived plants and animals. In the public perception, however, the arrival of wolves solved the problem of an overpopulation of moose. Another Look at Predictions - Will wolves stabilize prey fluctuations in Yellowstone, especially those of elk (Boyce 1993), or will wolves destabilize elk fluctuations, exacerbating population fluctuations (National Research Council 2002)? How far will the ecological ripple extend? Moose no longer number 3,000 on Isle Royale like they did before wolves (Allen 1979), so will elk ever exceed 19,000 again like they did before wolves and after the artificial reductions in Yellowstone? - Before wolf introduction, several studies used modeling to predict future impacts of wolves on the Yellowstone ecosystem (Yellowstone National Park et al. 1990, Varley and Brewster 1992, Cook 1993, U.S. Fish and Wildlife Service 1994). These were comprehensive efforts, prepared for Congress and the general public, which focused on the interaction of wolves with native ungulates, livestock, and grizzly bears. Simulations predicted between 50-120 resident wolves in YNP, with packs on the northern range, MadisonFirehole, and possibly the Gallatin and Thorofare areas (Fig. 3; Cook 1993:254-256). All models suggested that elk would constitute the primary prey for Yellowstone wolves. Four models dealt with the impact of wolves on native ungulates (Vales and Peek 1990, Garton et al. 1992, Boyce 1993, Mack and Singer 1992, 1993); no simulation predicted large declines in ungulates following wolf restoration. The northern Yellowstone elk population was predicted to decline 5-30% over the long term, with levels of decline contingent on the extent of hunter harvest of female elk outside the park (Mack and Singer 1993, Boyce 1993). Boyce (1993) suggested that some reduction of hunter kill of cow elk outside the park might be necessary over time, but restrictions on bull harvests would be unnecessary. Significant effects on other prey species (bison, moose, and mule deer) were not anticipated. In contrast to most predictions based on modeling, Messier et al. (1995) suggested that elk might decline substantially following wolf recovery because of the number of predator species involved. In boreal ecosystems where moose deal with multiple predators, moose density typically declines with each additional carnivore species (including human hunters; Gasaway et al. 1992). According to this thinking, the exceptionally high density of moose at Isle Royale (averaging about 2 per km2) occurs because there is only one predator - the wolf. Where wolves and bears coexist, calf survival is consistently reduced, and moose density is always less than 1 per km2, usually less than 0.4 per km2 (Messier 1994). The only geographic region where moose density is comparable to that of Isle Royale is Fennoscandia, where humans are the predominant predator species (bears have a minor presence), or the Gaspe Peninsula in New Brunswick, where there are black bears but no wolves and no hunting is allowed. Messier et al. (1995) believed that Yellowstone elk would decline significantly, more than the 5-30% predicted by Boyce (1993) and Mack and Singer (1993), especially where human hunting of cow elk was permitted. Focusing on the northern Yellowstone elk herd, at a pre-wolf winter elk density of >10 per km2, they anticipated that elk numbers would decline during the inevitable severe winters and would not rebound because of relatively low calf survival. What will be critical for elk recovery after declines will be the level of human hunting of elk outside the park, the only mortality factor that can be completely managed. Both the historical record at Isle Royale and the predictions of Boyce (1993) for the northern Yellowstone elk underscore the dynamic future that will follow wolf recovery. Fluctuations in wildlife populations are normal; the renowned "balance of nature" at Isle Royale is decidedly dynamic. Wolf peaks lag behind those of prey, and wolf declines follow prey declines. In the past four decades, two major declines in moose at Isle Royale have occurred when severe winters coincided with high moose density (> 3 per km2; Peterson 1995). Predictions for the wolf-prey system at YNP were similarly variable over time (Boyce 1993). Media attention and scientific debate have focused heavily on population size for northern Yellowstone elk. Average population size is an interesting statistic, but no one should expect elk to spend any time there. At most times, they will either be increasing or decreasing, and at any given time wolves and elk will probably show opposite trends. Isle Royale moose have spent more time below the population mean, probably because of suppression by wolves. Possibly this reflects the resilience of wolves in the face of prey decline, and the anti-regulatory (inversely density-dependent) influence of wolf predation that has been noted by wildlife managers in Alaska (Gasaway et al. 1992). An important question for Yellowstone, however, is to what extent will wolves prey on bison, a more formidable prey species that is more difficult to kill (Smith et al. 2000). If wolves do utilize bison, which are widespread and abundant at 4,000 animals, that will certainly change predictions of wolf impacts on elk. For the threatened grizzly bear population of GYE, wolf restoration was predicted to have either no impact or a slightly positive impact (Servheen and Knight 1993). Wolf predation on bear cubs was expected to be offset by better feeding conditions as bears usurp wolf-kills (Servheen and Knight 1993). Carcasses would be more evenly distributed for bears throughout their seasons of activity, rather than coming as a pulse in late winter and early spring—the pre-wolf condition. Bears would not have to risk killing elk themselves, but could scavenge kills of wolves well distributed in space and time. Although there was a general awareness of interspecific competition among native canids when the effects of wolf reintroduction were being assessed a decade ago, there were few predictions about exactly what wolf recovery would mean for coyotes, which on the northern range existed at one of the highest densities known for the species (Crabtree and Sheldon 1999). Some predicted wolves would reduce coyotes and that coyote reduction would affect other species (YNP et al. 1990, Varley and Brewster 1992). On Isle Royale, where wolves and coyotes competed for all the same prey species, wolves eliminated coyotes in about 8 years (Mech 1966). Prior to wolf reintroduction in Yellowstone, there were no predictions about possible responses in northern range vegetation caused by changes in distribution or density of ungulates, particularly elk. The forage for most ungulates wintering on the northern range--elk, bison, mule deer, bighorn sheep, pronghorn--is produced primarily in the extensive grasslands and shrub steppes. Grasslands are dominated by native species, although several alien grasses have been introduced (both accidentally and deliberately) and dominate local sites (Yellowstone National Park 1997, Stohlgren et al. 1999). A series of studies suggests that this grazing system is stable and highly productive; ungulate herbivory accelerates nutrient cycling and actually enhances productivity of the range (Houston 1982). Long-term changes in the vegetation (increased distribution and density of coniferous forests, increased abundance of big sage, decline in aspen and willow communities) seem to be associated with herbivory and suppression of natural fires, which occurred during a shift to a warmer, dryer climate (Meagher and Houston 1998). It's worth noting, however, that aspen and willow are minor components of northern range vegetation (less than 1 or 2 %); the difficulty of basing management of the larger grazing system on minor components of the vegetation is explored by Meagher and Houston (1998:247). Another unresolved point, far too complex to realistically simulate, is the productivity of the northern range, which nourishes the elk in winter. This is a unique north temperate grassland, one that has been compared to Africa's Serengeti. A much higher proportion of plant biomass can be consumed by ungulate grazers than ungulate browsers, which depend on annual growth of twigs and buds of woody shrubs. It is possible that the bottom-up stimulation of productivity from this grassland system will sustain elk at high density with a full suite of predators, both wild and human. A review committee of eminent scientists recently focused on the condition of the northern range (National Research Council 2002), concluding that high ungulate density was not causing irreversible damage to this ecosystem. Now that wolves are present, this committee firmly endorsed the scientific imperative to monitor ecosystem status closely. The Unfolding Yellowstone Story Wolf numbers. As we write (summer 2002), at least 216 free-ranging wolves (pre-pup 2002) can be found in the GYE, with about 14 packs (132 wolves) holding territories in or mostly within YNP, and 14 packs (84 wolves) outside (Fig.4). About 77 wolves (in 8 packs) occur on the northern range (very close to the number predicted for this area (Boyce 1993, Mack and Singer 1993)). Initial rate of increase for the wolf population was very high (Fig. 5), but now population growth within YNP has slowed, and most recent increases have occurred outside the park. Here we summarize the current status of wolves and their primary prey, the northern Yellowstone elk, and note some preliminary observations of other selected species affected by wolf recovery. Wolf territories. The northern range, targeted during reintroduction, is well-occupied by wolves, and virtually all potential wolf habitat in the park is occupied to some extent, including several areas that may not prove suitable for long-term occupancy (Fig. 4). Wolf packs have established year-round territories, despite the seasonally migratory nature of their ungulate prey. This was an important uncertainty before wolf introduction (Boyce 1993). The territories have been quite labile, and further subdivision seems likely, especially for the very large Druid Pack (37 wolves in August 2000, split into 4 packs April 2002), which now dominates much of the northern range in the park and has forced some packs into peripheral areas. Across the park, wolf packs exist approximately in the places predicted by Boyce (1993; Fig. 3). Wolf-prey relationships. As expected, elk are the primary prey for wolves in the park year round, representing 92% of 1,582 wolf kills recorded from 1995-2001. As elsewhere, wolf predation in winter has been highly selective; calves represent about 43% of wolf-killed elk, cows 36 %, and bulls 21 % (compared to the approximate winter population proportion of 14 % calves, 60 % cows, and 23% bulls). The adult elk killed by wolves have been very old, with a mean age of 14 years for wolf-killed cow elk (Mech et al. 2001). In contrast, human hunters outside the park kill female elk in their reproductive prime, at an average age of 6 years. Bull elk killed by wolves are taken primarily in late winter and average 5 years old, which is the same average age for hunter-killed bull elk. Examinations of femur marrow from the wolf-killed elk on the northern range indicate that 34% (N = 494) had exhausted all fat reserves. Although elk represent the primary prey for wolves throughout the park, bison are taken during late winter in interior portions of YNP (Smith et al. 2000) and moose are important along the southern boundary. Yet neither of these species represents more than 2% of wolf diet in winter (although in some areas during late winter it is higher). While wolves have killed some bison (Smith et al. 2000), so far most Yellowstone packs are supported almost entirely by elk. Coyotes: Before wolf reintroduction, coyote population density on the northern range was about 0.45 per km2, organized as packs with well-established borders (Crabtree and Sheldon 1999). Wolves began to kill coyotes soon after they were released in YNP. During 1996-1998 wolf aggression toward coyotes resulted in a 50 percent decline in coyote density (up to 90% decline in core areas occupied by wolf packs) and reduced coyote pack size on the northern range (Crabtree and Sheldon 1999). In the Lamar Valley of the northern range the coyote population declined from 80 to 36 animals from 1995 to 1998, and average pack size dropped from 6 to 3.8 animals (Crabtree and Shelton 1999). With lower coyote density, litter size increased, but the increased production of pups has been insufficient to offset the effects of wolves. Although data are preliminary, pronghorn fawn survival seems positively correlated with wolf density and inversely correlated with coyote density, as most fawn mortality is caused from coyote predation (J. Byers personal communication). In about 84% of 145 wolf-coyote interactions observed at wolf kills, wolves prevailed over coyotes. Wolf kills clearly provide food for coyotes (virtually all winter kills are visited by coyotes), but coyotes that scavenge wolf-kills risk death from wolves. Scavengers. Besides coyotes, nine other scavenger species have been observed using wolf kills. All wolf kills are visited by ravens, magpies, and eagles. Many kills in the non-winter months are visited by both species of bears. In winter wolf kills are tremendous centers of activity for scavengers, and small packs of wolves lose large amounts of food to scavengers (Hayes et al. 2000). Kills are especially important to ravens -- the average number of ravens per wolf kill was 29 and the most recorded was 135, a record in the literature (Stahler et al. 2002). Ravens follow wolves and discover carcasses immediately, or even before, flying overhead as wolves pursue and kill prey (Stahler et al. 2002). Grizzly bears. The grizzly bear population in the GYE has increased dramatically since the 1970's, although the bears are still listed as "threatened" under provisions of the Endangered Species Act. The population was estimated at 354 bears, including 35 sows with cubs at heel during 2001 (Haroldson and Frey 2001). Fifty-eight wolf-bear interactions have been recorded in YNP. Most interactions occur at wolf kill sites, where control of the carcass is hotly contested; typically bears win the encounter even though outnumbered by wolves. In one case a bear held 24 wolves at bay. Although fully capable of killing ungulates, especially in spring, grizzly bears now appear to seek out wolf kills and are often successful at driving wolves from carcasses. Cougars. The cougar population on the northern range has been monitored intensively through most of the 1990s (Murphy 1998). The present population on the northern range, roughly 25 animals, appears to have slowly increased during the 1990s and in the presence of wolves (T. Ruth, personal communication). Documented interactions between wolves and cougars have been rare, seemingly because of separation in habitat used by the species (cougars inhabit rock outcrops and cliffs along rivers). Field observations suggest that cougars avoid wolves, are subordinate at carcasses, and are at risk of predation. In one incident four cougar kittens were killed by wolves (T. Ruth, personal communication). Mesocarnivores. The effect of wolves on these animals has yet to be documented; we indulge in some speculation, however. Yellowstone has robust populations of some mid-sized carnivores (weasels, marten, badger), but low populations of others (fishers, wolverines, red fox, lynx, bobcat, otter). Several species may benefit from the advent of wolves. The red fox, for example, which competes more closely with coyotes than wolves, may increase because of lower numbers of coyotes. Wolverines, which scavenge carcasses, may also increase. Elk. From 1981-82 through 1994-95, winter numbers for the northern Yellowstone herd averaged 15,520+ 2,324 (SD) elk (Lemke et al 1998). Annual hunter harvests out side the park during the same 14 year period were variable but averaged 1,823+ 1,022 elk, including 1,192+ 661 animals taken during the late hunt (65% of the total harvest; Fig. 6). The late hunt targets "antlerless" elk (females and calves of the year) that migrate from YNP; most elk harvested each year are females, followed by calves, with a quota limiting bull harvests to around 100 animals (Lemke et al 1998). No elk counts were made during the 1995-96 or 1996-97 winters, just after wolves were released on the northern range (Fig. 6). The winter of 1996-97 was severe; rain on deep snow during December and January was followed by subzero temperatures, sealing off the supply of winter forage. Record ungulate migrations from YNP were documented and large numbers perished. A count in the following January, 1998, tallied 11,736 elk in the northern herd, lower even than after the drought, fires, and severe winter of 19881989. Counts for the next 4 winters, 1999 through 2002, ranged from 11,742 to 14,539 (Fig. 6). While the elk population appeared to rebound slightly after the severe winter of 1996-1997, the pace of recovery was evidently very slow compared to the one that followed the dieoff in 1989. Already, in the media, attention has abruptly switched from concern about too many elk for the northern range to too few elk for human hunters outside the park. It's worth noting that elk are the main prey for cougars, and cougars have a greater per capita kill rate than do wolves (Murphy 1998). Cougars kill an elk about every 9 days throughout the year (Murphy 1998); in winter a wolf kills an elk about every 15 days on average (Mech et al. 2001; summer wolf kill rates are unknown). Elk calves are also seasonally important in the diet of coyotes and grizzly and black bears (Clark et al. 1999). Grizzly bears also kill adult elk and bison. Coyotes were estimated to take over 1,200 elk annually from the northern herd (about the same number as grizzly bears and cougars combined) before wolf restoration (Crabtree and Sheldon 1999). For all these predators, elk calves are a major dietary component. Thus, in a very real sense, the abundance and survival of cow elk, through their annual production of young, support major links in the Yellowstone food web and will determine the trajectory of the elk population in the future. Population data for the non-migratory Madison-Firehole elk in YNP, which numbers about 500 animals, suggests that their numbers have been relatively stable since wolf reintroduction (Eberhardt et al. 1998). This herd, surviving near warm geothermal "oases" in a region with very deep snow, could be at risk from wolf predation if packs hunt more intensively in that area. Data are sparse for the other 6 migratory herds that occupy YNP during summer (total summer elk population is approximately 30-35,000 elk) but do not suggest herd declines after wolf arrival. Bison: About 4,000 bison occur park wide, with about 600-700 wintering on the northern range. Bison management has been controversial because some animals harbor the bacterial disease brucellosis, and there is a remote chance the disease could be transmitted to livestock when bison migrate from the park (U.S. Department of the Interior-National Park Service 2000). Wolves kill bison (Smith et al. 2000), but predation on bison is not yet widespread. One pack in Pelican Valley utilizes bison during late winter when bison are vulnerable and migratory elk are unavailable (Smith et al. 2000). Wolves are much less successful killing bison than killing elk; most bison stand their ground when confronted and this behavior seems to pose great difficulty for the attacking wolves. Bison carrion has been important wolf food during summers in the Lamar and Hayden Valleys whenever bulls die from injuries received during the rut. Bison carcasses attract concentrations of carnivores; including wolves, which scavenge extensively. Two grizzly bear cubs have probably been killed by wolves at these bison carcasses. Moose: The moose population on the northern range, numbering only a few hundred, declined precipitously following the 1988 fires (Tyers and Irby 1995). This occurred because subalpine fir forests burned extensively, eliminating for many decades—if not centuries--- these high-elevation moose habitats used in winter. Only 26 instances of wolf predation on moose have been recorded since wolf restoration. Bighorn Sheep. We do not expect wolves to affect the small population of about 175 bighorn sheep. Only one kill has been recorded since wolf reintroduction, and wolves spend very little time in the steep terrain commonly frequented by sheep. Deer. We also do not expect wolves to significantly affect deer populations. The park does not contain good white-tailed deer habitat, and the low numbers have not changed following wolf reintroduction. Wolves are not known to have killed any in the park. Mule deer are abundant numbering about 2-3,000 but migrate out of the park in winter, escaping some of the most intense wolf predation. Additionally, many mule deer winter in close association with humans, largely avoided by wolves. Beaver. Beaver are widely but patchily distributed in Yellowstone; most areas have few to none, although they are abundant in the Yellowstone River delta (south of Yellowstone Lake) and along the boundary of the park north of West Yellowstone, Montana. Systematic ground surveys began in 1988 and have continued at 5-year intervals; aerial surveys began in 1996 and have continued in alternate years. The 2001 aerial survey revealed 77 colonies distributed across the park. During 1996, shortly following wolf reintroduction, there were no documented beaver colonies on the northern range. Since then, beavers have established 4 colonies in this portion of YNP, following recent beaver reintroductions on the adjacent Gallatin National Forest (D. Tyers, U.S. Forest Service personal communication). Only two wolf pack territories contain substantial populations of beaver, the Yellowstone Delta and Cougar Creek packs (Fig. 4). Wolves likely prey on beavers, as elsewhere, but we have documented only one beaver kill, and have only rarely found beaver remains in wolf scats (but these two packs are not well sampled). Beavers are also closely associated with willow communities, a situation with relevance to wolves (see below). Vegetation: Interesting changes in willow and aspen growth have occurred in the late 1990s. Increased height of some aspen stands has been attributed to elk redistribution following the arrival of wolves (Ripple et al. 2001), but the initial trend has ceased. Some stands of willow have also increased in stature, but it is still too early to know if this is due to wolves. Recovery of woody plants would be consistent with the speculation that some cottonwood stands dating from the 1880s could not recover until elk numbers were reduced or displaced, although this may be too simple an interpretation as there has been strong debate on this issue (Meagher and Houston 1998, National Research Council 2002). No large stands of cottonwoods have been established on the northern range in the past 120 years. Currently very rare, but in some places increasing since wolf reintroduction, willow and aspen are important for many bird species, small mammals, beavers and moose. Ongoing research will continue to sort through this very complex issue, one that could be the most important impact of wolf reintroduction to Yellowstone. What Next for Yellowstone? Science Amid Management and Controversy Predicting the future of the Yellowstone ecosystem is professionally hazardous--some would say foolhardy. Nevertheless, some initial patterns have emerged, and we deem this preliminary look ahead worthwhile. Predictions help frame research agendas and prepare the public, perhaps lessening some of the controversy that seems to engulf the management of Yellowstone. At both Yellowstone and Isle Royale, before the arrival of wolves, the condition of the vegetation was a source of concern and disagreement (Mech 1966, Pritchard 1999, National Research Council 2002). The controversy evaporated at Isle Royale when wolves were established and a natural "balance" was assumed to exist, although some people feared the wolves would kill all the moose and then start in on the people (Isle Royale National Park files, letter to the Superintendent, 1956). Similar letters in numerous newspapers in the Yellowstone area have appeared regularly. Based on the Isle Royale experience, we anticipate that the condition of the vegetation on the northern range will subside as a popular debate topic in the media and that, in time, the fear that wolves will kill all the elk will also be put to rest. Much will depend on the population trajectory for Yellowstone wolves. At what point will wolves have saturated YNP, particularly the northern range, and how will we know when that point is reached? There are indications that rapid population growth for wolves on the northern range has ceased, but wolves should continue to increase until chronic food limitation is evidenced through declining numbers and weights of pups and intraspecific killing. While this is hardly the case at present, body weight for young wolves born in the Druid pack was lower in 2002 than in previous years. Field evidence that wolves are approaching their carrying capacity at Yellowstone is also suggested by increased levels of intraspecific strife in 2001 and 2002 and by a plot of wolf locations that indicates little vacant territory (Fig. 4). Isle Royale history and other studies indicates that there will be continual flux in pack territorial relationships (Fritts and Mech 1981, Mech et al. 1998). Wolf-prey ratios may help indicate when wolves have saturated their Yellowstone niche. After wolf recovery the average predicted elk:wolf ratio was 166, based on specific examples illustrated by Boyce (1993) for 100-year simulations, very similar to the elk:wolf ratio of 154 actually observed in 2002. At Isle Royale, during 1959-2002, predator and prey have fluctuated around a mean moose/wolf ratio of 46, but this figure has fluctuated from 18 to 145 (Peterson et al. 1998). We speculate that the difference in prey-to-predator ratios between Isle Royale and Yellowstone may arise from the differences in body size (moose are twice as large as elk) and social behavior (elk live in groups, moose live singly). We stress that it will be some time before anything resembling a dynamic equilibrium at Yellowstone can be actually documented. Before 1995, Isle Royale supported one of the highest year-round densities of wolves, 42 wolves/1000 km2 in the world. By 2002, wolves on Yellowstone's northern range had already reached a density of 50 wolves/1000 km2. Boyce (1993) predicted a mean population size of 76 wolves in YNP, primarily in the northern range, and a range from 50-120 under most management scenarios; in 2002 there were 77 wolves using the northern range and 132 wolves within YNP. The wolf population might overshoot equilibrium levels in a manner analogous to the irruptive pattern commonly observed for ungulate populations colonizing from low densities. The density reached by wolves in their initial period of rapid growth may indeed be very high, but this says little about the nature of the equilibrium which will be attained, perhaps decades hence. We are confident that form and function of the Yellowstone ecosystem will change because of wolf recovery. Reductions in the coyote population have already occurred, and elk numbers rebounded less after the severe winter of 1996-1997 but not to a degree that threatens the survival of either species in Yellowstone. A resulting trophic cascade will reverberate through the ecosystem. From the complicated food web that exists inYellowstone (Fig. 1), it is not hard to imagine that indirect effects of wolf recovery will be substantial. While riparian willow habitats form a very small part of the northern range, any reestablishment of these woody shrub communities would increase biodiversity. Although our expectations for wolf effects in Yellowstone are based on inferences from other studies, and may seem self-evident, we realize that specific predictions may be wrong. Even in a system as simple as Isle Royale, predictability has been poor following four decades of scientific scrutiny; none of the expectations for the moose herd, voiced in turn by Mech (1966), and Peterson (1977) actually happened. Rather, external forces such as severe winters, summer heat and outbreaks of winter ticks (driven by warm, dry spring weather) have caused the moose population to decline (DelGiudice et al. 1997). Surprises, as in the arrival of exotic disease which caused a wolf crash at Isle Royale in the early 1980s, are virtually guaranteed in the long term, and they will assuredly influence, and possibly determine, the outcome of the great natural experiment in wolf-elk dynamics now launched at Yellowstone. Science will be challenged to clarify exactly which changes in Yellowstone have been prompted by the addition of wolves. No wildlife population response at Yellowstone can be attributed to the actions of just one species (although coyotes may be an exception) or just one external event - such simplification of cause and effect is rarely possible in the science of ecology. Large perturbations, as with unique weather-driven events, will loom large in the future of Yellowstone. The 1988 fires burned about 36% of the land area of the park, affecting forage supplies for native ungulates (positively and negatively), but there is plenty of room for future fires in a climate that seems more conducive to large conflagrations. Given time, the severe winter of 1996-1997 will be matched and exceeded. Climate change will magnify the scientific challenge (National Research Council 2002). The danger we perceive is that all changes to the system, now and in the future, will be attributed entirely to restoration of the wolf. Testing research hypotheses is particularly difficult in natural areas, where experimental manipulations are limited and controls are either absent or difficult to establish. In particular, the problem of multiple causation has plagued the testing of hypotheses in ecology and has frequently confounded inferences derived from field studies (National Research Council 1997). This stumbling block may be particularly troublesome in the complex Yellowstone ecosystem; the need to design research hypotheses that discriminate among potential competing causes is very real. As in the past, elk management decisions outside the park will influence future population levels of elk inside the park. Some curtailment of midwinter shooting of cow elk outside the park might be necessary because wolves and humans, with very different hunting strategies, nevertheless compete over common prey. Successful coexistence of wolves and human hunters is a management conundrum that will test wildlife managers and challenge long-held beliefs. This is not the first time that the Yellowstone ecosystem has led us into uncharted waters, and our responses to this latest natural experiment will be no less interesting than the welter of ecological effects. Literature Cited Allen, D.L. 1979. The wolves of Minong: Their vital role in a wild community. Houghton Mifflin Company, Boston. 499 pp. Bangs, E.E., and S.H. Fritts. 1996. Reintroducing the gray wolf to central Idaho and -Yellowstone National Park. Wildlife Society Bulletin 24: 402-413. Boyce, M. S. 1993. Predicting the consequences of wolf recovery to ungulates in Yellowstone National Park. Pages 234-269 in Cook, R.S., ed. 1993. Ecological issues on reintroducing wolves into Yellowstone National Park. Scientific Monograph NPS/NRYELL/NRSM-93/22, USDI Nat. Park Serv., Denver, Colo. 328pp. Clark, T.W., A.P. Curlee, S.C. Minta, P.M. Kareiva. 1999. Carnivores in ecosystems: the Yellowstone experience. Yale University Press, New Haven. 429 pp. Cook, R. S. 1993. Ecological Issues on reintroducing wolves into Yellowstone National Park. Scientific Monograph NPS/NRYELL/NRSM-93/22, U.S. Department of the Interior, National Park Service, Washington, D. C., USA. Crabtree, R.L. and J. W. Sheldon. 1999. Coyotes and canid coexistence. Pages 127-163 in Clark, T.W., A.P. Curlee, S.C. Minta, and P.Kareiva, eds. Carnivores in Ecosystems: the Yellowstone Experience. Yale Univ. Press, New Haven, Conn. 429pp. DelGiudice, G.D., R.O. Peterson, and W.M. Samuel. 1997. Trends of winter nutritional restriction, ticks, and numbers of moose on Isle Royale. Journal of Wildlife Management 61:895-903. Eberhardt, L. L, R. A. Garrott, P. J. White, and P. J. Gogan. 1998. Alternative approaches to aerial censusing of elk. Journal of Wildlife Management 62:1046-1055. Fritts, S.H. and L.D. Mech. 1981. Dynamics, movements, and feeding ecology of a newly protected wolf population in northwestern Minnesota. Wildlife Monographs, Number 80. Garton, E.O., R.L. Crabtree, B.B. Ackerman, and Gerald Wright. 1990. The potential impact of a reintroduced wolf population on the northern Yellowstone elk herd. Pages 3-59 to 3-91 in Yellowstone National Park et al. Wolves for Yellowstone? Vol II. Nat. Park Serv., Yellowstone Nat. Park, Wyo. 586pp. Gasaway, W.C., R.P. Boertje, D.V. Grangaard, D.G Kelleyhouse, R.O. Stephenson, and -D.G. Larsen. 1992. The role of predation in limiting moose at low densities in Alaska -and Yukon and implications for conservation. Wildlife Monograph No. 120, 59pp. Haines, A.L. 1977. The Yellowstone story. Colorado Associated University Press, Vol 2, 543pp. Haroldson, M.A., and K. Frey. 2001. Grizzly bear mortalities. Pages 24-29 in C.C. Schwartz and M.A. Haroldson, editors. Yellowstone grizzly bear investigations: annual report of the Interagency Grizzly Bear Study Team, 2000. U.S. Geological Survey, Bozeman, MT. Hayes, R.D., A.M. Baer, U. Wotschikowsky, and A.S. Harestad. 2000. Kill rate by wolves on moose in the Yukon. Canadian Journal of Zoology 78:49-59. Houston, D.B. 1982. The northern Yellowstone elk: Ecology and Management. Macmillan Publishing Company, Inc., New York, New York, USA. Lemke, T.O., J.A. Mack, and D. B. Houston. 1998. Winter range expansion by the -northern Yellowstone elk herd. Intermountain J. of Sciences 4: 1-9. Mack, J.A. and F. Singer. 1992. Population models for elk, mule deer, and moose on Yellowstone's northern winter range. Pages 4-3 to 4-31 in J.D. Varley and W.G. Brewster, eds. Wolves for Yellowstone? A report to the United States Congress, Volume 4, research and analysis. USDI, NPS, Yellowstone National Park, Wyo. Mack, J. A. and F. J. Singer. 1993. Using Pop-II models to predict effects of wolf predation and hunter harvests on elk, mule deer, and moose on the northern range. Pages 49-74 in Cook, R. S., ed. 1993. Ecological issues on reintroducing wolves into Yellowstone National Park. Scientific Monograph NPS/NRYELL/NRSM-93/22. USDI Nat. Park Serv., Denver, Colo. 328pp. McLaren, B.E. 1996. Plant-specific response to herbivory: simulated browsing of suppressed balsam fir on Isle Royale. Ecology 77:228-235 McLaren, B.E. and R.O. Peterson. 1994. Wolves, moose, and tree rings on Isle Royale. Science 266:15551558 McLaren, B.E. and R.A. Janke. 1996. Seedbed and canopy cover effects on balsam fir seedling establishment in Isle Royale National Park. Canadian Journal of Forest Research 26:782-793. Meagher, M.M. 1973. The bison of Yellowstone National Park. National Park Service -Scientific Monograph Series, Number 1. 161 pp. Meagher, M.M. and D.B. Houston. 1998. Yellowstone and the biology of time: photographs across a century. University of Oklahoma Press, Norman. Mech, L. D. 1966. The wolves of Isle Royale. U.S. National Parks Fauna Series 7. Mech, L.D., L.G. Adams, T.J. Meier, J.W. Burch, and B.W. Dale. 1998. The wolves of Denali. University of Minnesota Press, Minneapolis. 227pp. Mech, L.D., D. W. Smith, K. M. Murphy, and D. R. MacNulty. 2001. Winter severity and wolf predation on a formerly wolf-free elk herd. Journal of Wildlife Management 65:998-1003. Messier, F. 1994. Ungulate population models with predation: a case study with the North American moose. Ecology 75:478-488. Messier, F., W.C. Gasaway, and R.O. Peterson. 1995. Wolf-ungulate interactions in the northern range of Yellowstone: Hypotheses, research priorities, and methodologies. Report to Yellowstone National Park. Murphy, K.M. 1998. The ecology of the cougar (Puma concolor) in the northern Yellowstone ecosystem: interactions with prey, bears, and humans. Ph.D. dissertation. Univ. of Idaho, Moscow. 147pp. National Research Council. 2002. Ecological dynamics on Yellowstone's northern range. National Academy Press, Washington, D.C. Pastor, J., B. Dewey, R.J. Naiman, P.F. MacInnes, and Y. Cohen. 1993. Moose browsing and soil fertility in the boreal forests of Isle Royale National Park. Ecology 74:467-480. Peterson, R.O. 1977. Wolf ecology and prey relationships on Isle Royale. U.S. Natl. -Park Serv. Sci. Monogr. Ser. 11. 210pp. Peterson, R.O. 1995. The wolves of Isle Royale. Willow Press Creek, Minocqua, Wisconsin. 190pp. Peterson, R.O., N.J. Thomas, J.M. Thurber, J.A. Vucetich, and T.A. Waite. 1998. Population limitation and the wolves of Isle Royale. Journal of Mammalogy 79:828-841. Phillips, M.P. and D.W. Smith. 1996. The wolves of Yellowstone Park. Voyageur Press. -Stillwater, MN. 125 pp. [Check this citation] Pritchard, J.A. 1999. Preserving Yellowstone's natural conditions: science and the -perception of nature. University of Nebraska Press, Lincoln. 370 pp. Ripple, W.J., E.J. Larsen, R.A. Renkin, and D.W. Smith. 2001. Trophic cascades among -wolves, elk, and aspen on Yellowstone National Park's northern range. Biological -Conservation 102: 227-234. Servheen, C. W., and R. R. Knight. 1993. Possible effects of a restored wolf population on grizzly bears in the Yellowstone area. P. 28-37 in R. S. Cook, ed. Ecological issues on reintroducing wolves into Yellowstone National Park. NPS/NRYELL/NRSM-93-22. Natl. Park Serv., Denver, Colo. Smith, D.W., L.D. Mech, M. Meagher, W.E. Clark, R. Jaffe, M.K. Phillips, and J.A. -Mack. 2000. Wolf—bison interactions in Yellowstone National Park. Journal of -Mammalogy 81: 1128-1135. Stahler, D.R., B. Heinrich, and D.W. Smith. 2002. Common ravens, Corvus corax, preferentially associate with gray wolves, Canis lupus, as a foraging strategy. Animal Behavior 64:283-290. Stohlgren, T.J., L.D. Schell, and B. Vander Heuvel. 1999. How grazing and soil quality affect native and exotic planr diversity in Rocky Mountain grasslands. Ecological Applications 9:45-64. Tyers, D. B., and L. R. Irby. 1995. Shiras moose winter habitat use in the upper Yellowstone River Valley prior to and after the 1988 fires. Alces 31:35-43. U.S. Department of Interior, National Park Service. 2000. Bison Management plan for the state of Montana and Yellowstone National Park: Final Environmental Impact Statement. Vol. I, D-655a. 847pp. U.S. Fish and Wildlife Service. 1994. The reintroduction of gray wolves to Yellowstone -National Park and Central Idaho. Final Environmental Impact Statement. U.S. Fish -and Wildl. Serv., Helena, Mont. 608pp. Vales, D. J., and J. M. Peek. 1990. Estimates of the potential interactions between hunter harvest and wolf predation on the Sand Creek, Idaho, and Gallatin, Montana, elk populations. Pages 3-93 to 3-167 in Yellowstone Nat. Park et al. Wolves for Yellowstone? Vol. II. Nat. Park Serv., Yellowstone Nat. Park, Wyo. 586pp. Varley, J.D., and W.G. Brewster. (eds.) 1992. Wolves for Yellowstone? A report to the -United States Congress. Research and analysis. U.S. National Park Serv., -Yellowstone National Park, Mammoth, Wyo. 750pp. Weaver, J. 1978. The wolves of Yellowstone. Natural Resources Report No. 14, National -Park Service. 38 pp. Yellowstone National Park, U. S. Fish and Wildlife Service, Univ. of Wyoming, Univ. of Idaho, Interagency Grizzly Bear Study Team, and Univ. of Minnesota Coop. Park Studies Unit. 1990. Wolves for Yellowstone? A Report to the United States Congress. Vol I, Executive Summary. Vol II, Research and Analysis. US Govt. Printing Office, Billings, Mont. Yellowstone National Park. 1997. Yellowstone's northern range: complexity and change in a wildland ecosystem. National Park Service, Mammoth Hot Springs, WY.