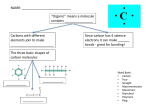
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Biochemistry Judit Kosáry (2016-1) The lecture gives a basic knowledge about the basic rules of living organisms (bioaffinity, biocatalysis, bioregulation), the role of different biomolecules (proteins, nucleic acids, carbohydrates, lipids) in living organisms, enzymes, the metabolism of biomolecules (catabolism, anabolism), generation and storage of metabolic energy, membrane transports. Biogenic elements Building biomolecules: carbon (C), hydrogen (H), oxygen (O), nitrogen (N) (and P és S). They are in the first and second periods (high charge concentration on their surface unit), their atoms are not susceptible to deformation and they form with own atoms and other biogenic elements strong -bonds. Electronegativity can be characterized the atoms connected by a -bond. The atom with higher electronegativity can collect a part of the electron density of the -bond, this causes an electron surplus (Ө) on it. This effect causes an electron deficiency () on the atom with lower electronegativity. EN 1 … 4 5 6 Columns EN Periodes 1 H EN=2.1 2 C N O EN=2.5 EN=3.0 EN=3.5 3 P S EN=2.2 EN=2.5 Position of biogenic elements in the periodical system and their electronegativity (EN) Biomolecules Biomolecules are organic molecules building up living organisms. Types of biomolecules: proteins, carbohydrates, nucleic acids and lipids (apolar biomolecules). Proteins, carbohydrates, nucleic acids are chiral biopolymers: Type of biomolecule Proteins Units -Amino acids Bonds between units Peptide bond (a special carboxamide bond) Carbohydrates Simple sugars O-Glycosidic bond (a special acetal bond) Nucleic acids Nucleotides 3’,5’-Phosphodiester bond ______________________ ______________________ _____________________ Lipids (apolar Simple lipids cannot be Complex lipids can be biomolecules) hydrolyzed by NaOH hydrolyzed by NaOH Characteristic data of the structure of proteins, carbohydrates and nucleic acids; characterization of lipids 2 Rules of biochemistry at the molecular level 1. Bioaffinity – there is at least one biological surface to interact with a biomolecule. 2. Biocatalysis – in living organisms practically all of reactions are catalyzed and the biocatalysts are called enzymes (mostly proteins). 3. Bioregulation – all of biochemical processes are regulated. Isomerism in biomolecules When two molecules have some differences in their structure but their molecular formula (the composition of elements) is the same, they are isomers. When the atoms bond in different order in isomers they are structural (constitutional) isomers. There are different types of structural isomers. In biomolecules tautomerism (the difference between two isomers is in the position of one hydrogen atom and a double bond) can be found frequently (e.g. aldoses and ketoses in carbohydrate chemistry). The stereoisomers have the same molecular formula and sequence of bonded atoms (constitution), but their atoms have differences in their three-dimensional orientation in space. There are different types of stereoisomers: optical isomers (enantiomers and diastereomers), geometrical isomers and conformers. Conformational isomers (conformers) differ by rotations around one or more single bonds (e.g. chair and sofa conformations of glucopyranoside). Optical isomerism In the case of saturated carbon atoms, due to hibridization (sp3), the angle of the bonds is 109,5°, i.e. its geometry is tetrahedral. A carbon atom with four different substituents (marked by a star) is called a chiral carbon atom (on the basis of the Greek word kheir– hand). In the case of a single chiral atom two isomers, called enantiomers are possible. Enantiomers (antipodes) are related as mirror images. The chemical and physical properties of the enantiomers are the same, because the microenvironment of the atoms is the same. The only difference is in their optical rotation, which is opposite. An enantiomer can be identified by the direction in which it rotates the plane of monochromatic and monopolarized light. If it rotates the light clockwise, that enantiomer is labeled (+), while its mirror-image is labeled (−). Group of highest oxidation number | Smallest functional group–C–Characteristic functional group | Other group The application of Fischer’s convention on the D- enantiomer Group of highest oxidation number | Characteristic functional group–C–Smallest functional group | Other group The application of Fischer’s convention on the L-enantiomer 3 CHO CHO HO C H HO C H CH2OH CHO CHO H C OH H C OH CH2OH CH2OH L-glicerinaldehid CH2OH D-glicerinaldehid CH2OH HO C H CHO Az aszimmetria-centrumok Fischer-féle ábrázolása COOH C R H NH2 COOH H2N C H R H2N COOH CH R Az L-aminosavak térbeli, Fischer-féle és a speciális, a Fischer-féle konvención alapuló ábrázolása The modified Fischer convention for the L--amino acids Distinction of enantiomers can be carried out by application of the Fischer convention that is based on the simplest aldose glyceraldehyde. When the characteristic group is on right side in Fischer projection, the enantiomer is called right-handed (D – after Latin word dexter) and the other enantiomer is the left-handed variation (L – after Latin word laevus). D and L are typically typeset in SMALL CAPS. Nowadays, except for sugars and amino acids, for the definition of chirality the R/S notation is used. This is defined by the Cahn-Ingold-Prelog priority rules based on atomic number. In the case of glyceraldehyde the D enantiomer is the clockwise (+) and L-enantiomer is the anti-clockwise (–) enantiomer. For the L--amino acids a modified Fischer convention is used because it is better for illustration of the peptide bond. Diastereomers contain at least two centers of chirality, and one of the centers has the same and the other has the opposite position. Diastereomers are not mirror images and therefore their chemical and physical properties of are different, the microenvironment of the atoms being different. This fact can serve as a basis for the separation of enantiomers from their mixtures (called racemic mixtures) by forming diastereomers. This process is called resolution. D L | | D D Diastereomers In compounds containing two chirality centers and the carbon atoms have the same substituents of opposite chirality therefore the direction of the optical rotation of two carbon atoms is opposite (resultant optical rotation is zero) – this variation is called meso form (e.g. meso-tartarate). H C OH HO C H HO C–H H 4 C–OH COOH D H H C L COOH COOH H OH C OH HO COOH mezo-borkôsav D D C OH C H COOH (+)-borkôsav Borkôsav (tartarát) diasztereomerek Meso-tartarate and (+)-tartarate are diastereomers Many biologically active molecules are chiral, including the naturally occurring proteins, carbohydrates and nucleic acids. As enzymes are mostly proteins and proteins are chiral, they preferentially catalyze the transformation of only one of the enantiomers of a chiral substrate. Naturally occurring proteins are made of L-amino acids, carbohydrates, di-, oligo- and polysaccharides are all made of D-sugars. Nucleic acids contain also D-sugars: ribose or deoxyribose. Geometrical isomerism In the case of free rotation in saturated hydrocarbons in eclipsed structure – hydrogen or other atoms are in disturbing each other; in the open structure – hydrogen or other atoms are in the in a rotated position, causing minimal disturbance, therefore this structure is preferred. H HH H3C CH3 etán H H H H rotáció H fedô állás (kedvezôtlen) H H H H nyitott állás (kedvezô) konformációk Az etán konformációi Conformations of ethane:eclipsed and open structures When there is a barrier to rotation (e.g. a ring system or a double bond) the large substituents can be on the same or on opposite sides. When large substituents are on the opposite side (trans isomer) the disturbance is less and therefore this isomer is more advantageous than when they are on the same side (cis isomer). Now in organic chemistry E-Z notation based on the Cahn-Ingold –Prelog rules is used. H H H Br C C C C Br Br Br H cisz-1,2-dibróm-etilén kevésbé stabil transz-1,2-dibróm-etilén stabilabb A simple example for geometric isomers: cis- and trans-1,2-dibromoethylene Carbohydrates 5 Carbohydrates (saccharides) are organic compounds of the general formula Cn(H2O)n. The ratio of hydrogen and oxygen is 2:1 as in the water. Formerly carbohydrates were viewed as hydrates of carbon and this is the origin of their name. In simple sugars (monosaccharides) the general formula is CnH2nOn (generally C3-C7). They are polyhydroxy carbonyl (oxo) compounds. The carbonyl group may be aldehyde (aldose) or ketone (ketose). In ketoses the carbonyl group is always at position 2 of the sugar. The sugars are named not only based on the type of their carbonyl group but by the number of the carbon atoms (on the basis of Greek name of numbers): triose (C3 – C3H6O3), tetrose (C4 – C4H8O4), pentose (C5 – C5H10O5), hexose (C6 – C6H12O6), heptose (C7 – C7H14O7), e.g. glyceraldehyde is an aldotriose, glucose is an aldohexose and fructose is a ketohexose. The secondary hydroxy groups of sugars are stereocenters and the chirality of the sugar is based on the configuration of the chiral carbon atom of highest number (e.g. C5 in hexoses). The different aldoses and ketoses with the same number of carbon atoms are diastereomers except one, which is their enantiomer. In the living organisms sugars are generally in D- and not L-enantiomer form It is known that only one kind of enantiomers can connect to different biological surfaces. Sugars are usually metabolized as their phosphate esters. HO O O HO HO CH CHO CHO CHO CHO H C OH H C OH H C OH H C OH CH2OH H C OH H C OH CH2OH H C OH H C OH CH2OH H C OH D-glicerinaldehid D-eritróz CH2OH D-ribóz L-aszkorbinsav aldózok HO C H CH2OH D-glükóz (C-vitamin) CH2OH CH2OH CH2OH CH2OH CH2OH C O C O C O C O C O HO C H HO C H HO C H CH2OH dihidroxi-aceton H C OH H C OH H C OH H C OH CH2OH CH2OH H C OH H C OH CH2OH H C OH D-fruktóz CH2OH D-ribulóz D-xilulóz ketózok A legfontosabb egyszerû cukrok és a C-vitamin képlete H C OH D-szeduheptulóz The most important straight-chain simple sugars and a sugar derivative L-ascorbic acid (ascorbate, glyceraldehyde, erithrose, ribose, glucose, dihydroxyacetone, ribulose, xylulose, fructose, sedoheptulose In sugars the different functional groups retain their original properties, therefore aldohexoses can be easily oxidized and carbonyl group can react with one of hydroxyl groups. Detection of sugars by oxidation: according to the condition of chemical oxidizability (i.e. the presence of a hydrogen connected to a carbon that is in a 6 polarized bond) only aldehydes can be easily oxidized to carboxylic acids. The example of acetaldehyde is presented: The Fehling reaction of acetaldehyde forming acetate and Cu2O as red precipitate During the reaction the oxidation of aldehyde is combined with the reduction of the oxidizing reagent (on this case 2Cu2 Cu22), therefore aldoses are called reducing sugars. Ketones cannot be oxidized because of the absence of CH in carbonyl group. But in forced conditions ketoses can be isomerized to aldoses by double oxo-enol-oxo tautomerism, therefore all of simple sugars are reducing sugars. H C O H C OH HO C H H C O H C O H H C OH H C OH HO C H O H HO C H H C OH H C OH H C OH H C OH H C H C CH2–OH CH2OH CH2OH CH2OH H HO O H OH O H OH H OH H - D-glükopiranozid Az egyszerû cukrok ciklizálódása Cyclization of simple sugars The cyclization of sugars is caused by a reversible intramolecular reaction (nucleophilic addition) between the carbonyl and one of hydroxyl group of the straight-chain (open form) sugar. This hydroxyl group is connected to the chiral carbon atom of highest number. The cycle that contains an oxygen and the new hydroxyl groups is connected to chiral carbon atom can be in different positions forming stereoisomers. Aldopentoses and ketohexoses form five-membered rings with plane surface (furanosides). Aldohexoses form three dimensional six-membered rings (pyranosides), those are generally projected as planar, and D-hydroxy groups are under the ring and L-hydroxy groups are above the ring. Cyclization generates a new 7 chiral center, in which the D-hydroxyl group is called the -anomer and the Lhydroxy group is called the -anomer. CH2OH O H H H OH H HO H OH OH H CH2OH O HO H H HO H OH H OH - D-glükopiranozid H HO HO CH2OH O H H OH OH H H - D-glükopiranozid A gyûrüs glükóz ábrázolási lehetôségei és glikozidos kötésének térállása Representations of - and -D-glucopyranoside The cyclized sugars contain a hemiacetal or hemiketal structure (in sugars the new hydroxyl group is called the glycosidic hydroxyl) that can be easily reacted with a hydroxyl group of another sugar. The product is a disaccharide. A ring-formed sugar can be reacted with a variety of hydroxyl groups; the general name of the product is glycoside that is a special form of an acetal. Because of the reversibility of this cyclization the disaccharides containing glycosidic hydroxyl group (maltose, cellobiose, lactose, gentiobiose) are reducing compounds. In saccharose (sucrose) the glycosidic bond is formed between two glycosidic hydroxyl groups therefore it is not a reducing sugar. The connection between two sugars in disaccharides is shown by their names (e.g. maltose is D-glucopyranosyl-[1,4-]-D-glucopyranose). 8 H HO CH2OH O H OH H HH O OH H CH2OH O H OH H H H H OH HO H CH2OH O H OH H H HO HOCH2 H H OH O O H HO CH2OH CH2OH O H OH H HO H H OH H O OH H H laktóz CH2OH O H OH H H CH2OH O H OH H OH H OH H szaharóz HO OH H H cellobióz H H H O maltóz CH2OH O H OH H OH OH H OH CH2OH O H OH H HOCH2 O CH2 H H OH HO genciobióz O H OH H H OH H H OH O H HO OH CH2OH OH H - D-fruktofuranóz Az ismertebb diszacharidok és a ciklizált fruktóz képlete The most important disaccharides and the ring-formed fructose Among polysaccharides starch and cellulose are the most important. The building unit of starch is maltose. Plants use starch as energy reserve. Its function is similar to that of glycogen in animals and people. There are two types of starch: amilose (a linear polymer containing only 1,4 glycosidic bonds) and amilopectin (a branched polymer containing both 1,4 and 1,6 glycosidic bonds). The building unit of cellulose is cellobiose. Cellulose is a structural components of primary cell walls of green plants and is the most wide-spread organic molecule in the world. The name of the polysaccharides containing only D-glucose molecules is glucan – starch is an glucan and cellulose is a -glucan. Proteins Proteins (polypeptides) are biopolymers made of -L-amino acids connected by peptide bonds (a special type of carboxamide bond). Units of the polypeptide chain, the L--amino acids They are the building blocks of proteins connected by peptide bonds. Standard (protein, proteinogenous) amino acids build up proteins, non-standard (non-protein, non-proteinogenous) amino acids can be important metabolic intermediates. The name of standard amino acids is used generally in their abbreviated form. The modified 9 Fischer conventions of the formulas of twenty standard amino acids and their abbreviations are presented in schemes. Ten of amino acids (Val, Leu, Ile, Phe, Lys, Thr, Trp, Met, Arg, His) are called essential amino acids, because the human body cannot synthesize them from other compounds at the level needed for normal growth, therefore they must be obtained from food. (Notice: while large quantities of the essential amino acids are needed, there are other essential compounds, e.g. vitamins, which we need only in small quantities). Often selenocysteine and taurine are also put on the list of standard amino acids, while Arg, His are classified as semiessential amino acids by several authors. H2N CH2 COOH H2N glicin (Gly) CH COOH H2N CH3 alanin (Ala) CH COOH CH3 H2N COOH CH CH2 CH3 CH CH3 CH3 leucin (Leu) COOH CH2 CH CH2 CH COOH CH CH valin (Val) H2N H2N H N COOH CH3 CH3 izoleucin (Ile) prolin (Pro) fenilalanin (Phe) A hidrofób kölcsönhatásra alkalmas fehérjealkotó aminosavak Amino acids of hydrophobic character H2N CH COOH H2N CH COOH H2N CH COOH H2N CH COOH CH2 CH2 (CH2)4 (CH2)3 COOH CH2 NH2 NH aszparaginsav (Asp) COOH lizin (Lys) glutaminsav (Glu) C=NH NH2 arginin (Arg) Az ionos kölcsönhatásra alkalmas fehérjealkotó aminosavak Amino acids with ionic character 10 H2N CH H2N COOH CH COOH H2N CH CH2OH CH3 szerin (Ser) CH COOH CH2 OH treonin (Thr) H2N CH H2N COOH CH COOH CH2 CH2 CONH2 CH2 OH tirozin (Tyr) CONH2 aszparagin (Asn) glutamin (Gln) H2N CH H2N COOH CH COOH CH2 CH2 NH N N H triptofán (Trp) hisztidin (His) A hidrogénkötésre alkalmas fehérjealkotó aminosavak Amino acids with hydrogen bonds H2N H CH 2N 2 Cl CH2 CH2COOH CH OH Cl Na O–H CH2SH cisztein (Cys) etilén-klórhidrin Cysteine with CH3 CH2 CH2 O CH2 COOH CH2 H + A dipólus-dipólus kölcsönhatásra AN H O fehérjealkotó CH2 CH2 O H alkalmas aminosav H S CH3 metán-tiol CH3CH2 O CH2CH3 dietiléter SNi szomszéd-csoport hatás metionin (Met) H2O etilénglikol disulphide bond and methionine with dipole-dipole Az etilénklórhidron reakciója S CH2 O etilén-oxid S–CH3 (szögfeszültség) H2O A diszulfidkötésre alkalmas NaOH CH2 CHaminosav fehérjealkotó 2 O CH "O" H2O "O" CH3CH2 CH3 S S interaction CH3 stabil diszulfid híd O O–CH2CH3 dietil-peroxid A peroxidok és a diszulfidok stabilitási különbsége Formation of disulphide bond and peroxides 2 CH3CH2 O 11 Structural levels of proteins Primary structure: The sequence of amino acids. On one end of every polypeptide chain, called the amino terminal or N-terminal, there is a free amino group. The other end, with its free carboxyl group, is called the carboxyl terminal or C-terminal. O H2N O CH N R H CH OH N-terminális C-terminális A fehérjék elsôdleges szerkezete Primary structure of proteins with the N- and C-terminals of the chain Peptide bonds are special carboxamide bonds with strong hydrogen bonds caused by a partial delocalization in the functional group. Because of this delocalization the peptide bond is planar and rigid. This partial delocalization is illustrated by the molecule acetamid. CH3 C O NH2 acetamid CH3 C O O N H H kis O CH3 C NH2 C C O N C C C H N C H dipoláris, gátolt rotációjú szakasz a savamidcsoportban (a peptidkötésben) NaOH nagyon nehezen A savamidcsoport jellemzése Partial delocalization and hindered rotation of acetamid illustrated by mesomeric structures Secondary structure:– Structures established by hydrogen bonds between peptide bonds: righ-handed -helix, -sheet – between antiparallel chains, collagen structures – there are three of left-handed extended helix structures rolled into a cable form of a right-handed helix in tropocollagen units containing Gly-Pro-Hyp triplets, 12 hydroxyproline is synthesized by a direct oxidation of proline in peptide chain by means of L-ascorbate). -helix structure a -sheet structure collagen structure O N O N 1/2 O2 (az aszkorbinsav közvetítésével) HO Hyp részlet a Pro részlet a fehérjeláncban fehérjeláncban A hidroxi-prolin képzôdése a peptidláncban Oxidation of proline to hydroxyproline in the peptide chain by L-ascorbate (vitamin C) Tertiary structure: – Connections between remote parts of the peptide chain by secondary bonds between the side chains of amino acids – globular structures (folded to three dimensional structures, they contain all of the secondary structures) and fibrous structures (folded to fibres, they contain only one of the secondary structures). Interactions: hydrophobic interactions – glycine (Gly), alanine (Ala), valine (Val), leucine (Leu), isoleucine (Ile), phenylalanine (Phe), proline (Pro) ionic interactions – aspartic acid (Asp) glutamic acid (Glu), lysine (Lys), arginine (Arg) hydrogen bonds – serine (Ser), threonine (Thr), tyrosine (Tyr), asparagine (Asn), glutamine (Glu), tryptophan (Trp), histidine (His) disulphide bond – cysteine (Cys) dipole-dipole interactions methionine (Met); Quaternery structure:– Connection between several polypeptide chains usually called protein subunits by secondary bonds between the side chains of amino acids. 13 Simple proteins contain only protein chains. Complex proteins contain other kinds of biomolecules or metal ions: glycoproteins (often in membranes), nucleoproteins (in ribosomes), lipoproteins (e.g. LDL – a cholesterol transferring lipoprotein), metalloproteins (e.g. some enzymes as lactate dehydrogenase contain zinc), chromoproteins (e.g. red hemoglobin), phosphoproteins (e.g. casein), etc. Biological function of proteins Enzyme proteins – catalysts of biochemical reactions, they are vital to metabolism Structural proteins – e.g. collage fibers as fibrin Contractile (mechanical) proteins – e.g. muscle proteins Transport proteins – e.g. hemoglobin transports oxygen Proteins for supply – e.g. myoglobin supplies oxygen Immune protection – etc. immunoglobulins Toxins (poisons) – e.g. snakes poison Biuret reaction is a colorimetric protein assay methods that use cupric ions as colouring agent. Cupric ions form a complex of faint blue-violet color with the imide tautomer of at least two (according to several authors four) peptide bonds. The intensity of the color produced is proportional to the number of peptide bonds participating in the reaction; therefore the biuret reaction is an often used analytical method for the quantitative determination the total protein concentration. The reaction was named after the organic compound biuret (NH2-CO-NH-CO-NH2) that is the simplest compound to give a colored (light blue) complex. O CH C R O H O NaOH N CH C N H R H R Cu O CH C R CH C O O H N CH C N Cu2+ R imid (a nátrium-hidroxiddal sót képezhet) N CH C N R ibolyaszínû komplex A biuret reakció Enzymes Enzymes are globular proteins generally with quaternary structure. As biocatalysts they give an alternative reaction for the product synthesis with lower activity energy than the original reaction of really high activity via forming a complex with substrate. Since enzymes are selective for their substrates and speed up only a few reactions from among many possibilities, the set of enzymes made in a cell determines which metabolic pathways occur in that cell. Enzymes are known to catalyze about 4,000 biochemical reactions. Activity of enzyme is affected by temperature, chemical environment (e.g., pH and salt concentration), and the concentration of substrate. 14 Reaction diagram without and with enzyme Enzyme reactions are reversible. The sum of the rate of the dissociation of the enzyme substrate complex (v-1) and the rate of the synthesis of product and regeneration of the enzyme (v2) from this complex can be equal to the rate of forming enzyme substrate complex (v1), this status is called ‘steady state’. E+S v1= k1[E].[S] v1 v-1 ES v2 v-1= k-1[ES] E+P v2= k2[ES] A saturation curve can be found when the concentration of the product [P] is plotted against reaction time. Also a saturation curve can be found for the relation between the substrate concentration [S] and rate (v0). This rate (v0) is the rate of enzyme reaction at the first period of the reaction. This can be characterized by the modified Michaelis-Menten plot that is called the equation of enzyme kinetics. As the substrate concentration increases, more and more of the free enzyme is converted into the substrate-bound ES form. At the maximum rate (Vmax) of the enzyme, all the enzyme active sites are bound to substrate, and the amount of ES complex is the same as the total amount of enzyme. The amount of substrate needed to achieve a given rate of reaction is also important. This is given by the Michaelis constant (KM), which is the substrate concentration required for an enzyme to reach one-half its maximum rate. 15 Diagrams and equal of enzyme kinetics Only the active site of an enzyme takes part in the catalytic reaction while other parts of the enzyme assure the active conformation of the active site, that contains two important parts. The substrate binding site can be characterized by KM for a given substrate, and this can show how tight the binding of the substrate is to the enzyme. The parameters and/or compounds decreasing the binding of the substrate can increase the value of KM. For a given substrate the catalytic site can be characterized by Vmax. The parameters and/or compounds decreasing the transformation of the substrate-enzyme complex to the product can decrease the value of Vmax. The double reciprocal plot The KM and Vmax values are the important kinetic constants of the kinetics of enzymes for a given substrate. The determination of these constants is given by a double reciprocal plot (Lineweaver-Burk plot) that yields a straight line with an intercept of 1/Vmax and a slope of KM/Vmax. Certain compounds can alter the activity of enzymes. Enzyme activity can be decreased by various inhibitors or can be increased by activators. The effect of such compounds can be reversible or irreversible. Reversible inhibitors are classified according to their linkage to the active site. Compounds of similar structure to the substrate can bind to the substrate binding site and are called competitive inhibitors. Compounds which disturb the function of the catalytic site are called non-competitive inhibitors. Compounds that can disturb the function of both the substrate binding and catalytic sites are called mixed inhibitors. 16 The types of reversible inhibitions: competitive inhibition, mixed inhibition, noncompetitive inhibition The classification of enzymes – enzymes can be identified by their number in Enzyme Nomenclature (Enzyme Catalogue EC). EC number is a combination of four numbers. The first number of the combination shows the type of the reaction catalyzed. 1. Oxidorecuctases – catalyze oxidation and reduction (dehydrogenases and oxigenases) 2. Transferases – catalyze subtitutions 3. Hydrolases – catalyze hydrolysis 4. Lyases – catalyze addition and elimination 5. Isomerases – catalyze tautomerism 6. Ligases – catalyze reactions using the energy of macroerg bonds Oxidoreductases and transferases need reagents (compounds with coenzyme function) for the catalyzed reactions. Compounds with coenzyme function (henceforth they are called as coenzymes) are connected to enzymes either by secondary bonds (they are really coenzymes – they can be regenerated also in other reactions) or by covalent bonds (prosthetic groups – they can be regenerated only in their original place). Compounds with coenzyme function have two forms (unreacted and reacted) – only lipoic acid has three forms. The starting materials for coenzymes are water soluble vitamins and in a few cases essential amino acids). In primary metabolism oxidoreductases are always dehydrogenases, because the reoxidation of reduced coenzymes is connected with the producing of energy in form of macroerg bonds. The mechanism of these oxidoreductase coenzymes can be ionic (hydrogen molecules are transported as hydride anions and protons) or radical (one hydrogen molecule is transported in form of two hydrogen atoms). In the oxidative degradative processes of catabolism NAD (its starting material is nicotinamide i.e. vitamin B3) – its reduced form is (NADH+H) (nicotinamide adenine dinucleotide) involve an ionic, while FAD (its starting material is riboflavine i.e. vitamin B2) – its reduced form is FADH2 (flavin adenine dinucteotide) and FMN – its reduced form is FMNH2 (flavin mononucleotide) a radical mechanism. FMN takes part only in terminal oxidation. In reductive biosyntheses of anabolism the coenzyme is (NADPH+H) in both mechanisms. The difference between NAD NADP is the presence of a phosphoryl group on the C-2 hydroxyl group of ribose in NADP. Flavin-containing coenzymes are always prosthetic groups. 17 H H H H O H O CONH2 HN HN N CONH2 NH + H O N H (DHU) = 260dihidro–uracil nm HOCH2O O O N H H H H max = 260 és 340 nm max OH OH pszeudo uridin (C) A nikotinamidot tartalmazó koenzimek redukálódási folyamata Néhány ritka nukleotid képlete The process of reduction of coenzymes containing a nicotinamide structure H H H CONH2 CONH2 CH2 O H P O H H OH OH NH2 N H N P H H H OH OH NH2 N H O P N N O CH2 2H (H + H ) O H H H H H R = H (NADH + H+ ) R = P (NADPH + H+ ) A NAD+ és NADP+ koenzimek Coenzymes NAD and NADPH H N 2H N N OH OR OH OR R = H (NAD+ ) nikotinamid-adenin-dinukleotid R = P (NADP+ ) N H N N O CH2 H H N O O H P O CH2 N O N H A flavint tartalmazó koenzimek redukálódási folyamata The process of reduction of coenzymes containing flavines 18 O H3C H3C N H NH N N O H3C N H3C N N CH2 H NH 2H O CH2 NH2 HCOH HCOH H2C O P O P O CH2 H HCOH H2C O H H N N HCOH N N O NH2 HCOH N N HCOH O P O H H N N O P O CH2 H H OH OH H OH OH FADH2 FAD (flavin-adenin-dinukleotid) O H3C N H3C N NH N CH2 HCOH O R = P FMN (flavin mononukleotid) R = H (B2 vitamin) riboflavin HCOH HCOH H2C O–R A flavint tartalmazó koenzimek és prekurzor vitaminjuk Flavin-containing coenzymes and their precursor vitamin Ubiquinone (coenzyme Q) (its starting material is tyrosine and its reduced form is ubiquinol) is that kind of oxidoreductase coenzyme, which can work by both ionic and radical mechanism. The name of the human ubiquinone is CoQ10. The starting material of ubiquinone is tyrosine. Redox reactions of ubiquinone In various kinds of cytochromes the coenzyme effecting electron transfer is hem (by ferrous-ferric transformation). 19 The structure of hem The coenzyme of direct oxygenases is ascorbic acid (vitamin C). O O HO ox O HO HO C H CH2OH L-aszkorbinsav (C-vitamin) red O O O HO C H CH2OH dehidro-aszkorbinsav (bomlékony) Az aszkorbinsav oxidált és redukált formája Redox reactions of L-ascorbic acid NH2 N Transferases can catalyze several N kinds of substitutions. The transferred groups can be different carbon skeletons: C1 – CO2 (biotin that is vitamin H), only methyl group (SAM – S-adenosylmethionine, N itsNstarting material is methionine), methyl group, aldehyde group, etc. (THF – tetrahydrofolate, its starting material is P–O–P–O–P–CH2 O folic acid i.e. vitamin B9 – earlier vitamin B10; C2 – acetaldehyde (TPP – thiamine H H i.e. vitamin B ), acetyl group in a pyrophosphate, its staring material is aneurine, 1 H H macroerg thiolester bond (coenzyme A, its starting material is pantothenic acid i.e. vitamin B5; and lipoic acid that is connected to the -amino group of a lysine as a OH OH prosthetic group therefore it is often called lipoamide); and other groups: phosphate group (ATP or other nucleoside triphosphate molecules), amino group (PAL – Az ATP átadható csoportjai pyridoxal phosphate, its reacted form is PAM – pyridoxamine phosphate, and its starting material is pyridoxine i.e. vitamin B6). 20 O HN C ATP O ADP HOOC NH CH2 S CH2 biotin (H-vitamin) CH2 CH2 N C NH CO2 COOH CH2 CH2 CH2 CH2 COOH S karboxi-biotin A biotin keletkezése és formái Transfer coenzyme – carbon dioxide – biotin NH2 H2N COOH CH CH2 O P P P –O–CH2 O CH2 H OH CH3 Met Pi PPi N H H + OH ATP COOH N N CH2 S CH3 NH2 CH CH2 H3C N O H S H2N N N CH2 H H N N O H2N COOH NH2 CH CH2 CH2 S N N CH2 H H H OH OH S-adenozil-metionin (SAM) N N O H H OH H OH S-adenozil-homocisztein (SAH) A SAM keletkezése és különbözô formái Transfer coenzyme – methyl group – SAM 21 CH2 O H 4 5 6 8 7 1 N H2N CH2–NH N HN 3 2 C1 CH 4-aminobenzoesav N COOH COHN H C1: CHO CH2 CH3 CH2 CH2OH COOH tetrahidro-folsav (THF) Glu O N H2N CH2–NH N HN COOH COHN CH CH2 N CH2 folsav (B10 vitamin) COOH A C1 részleteket szállító koenzim és prekoenzim vitaminja Transfer coenzyme – C1 – THF H3C CH2CH2O N H3C CH2 N NH2 O P H3C H3C CH2 N C N tiamin-pirofoszfát (TPP) H3C O S CH2CH2OH CH2 N CH N P NH2 H3C C OH H "aktív acetaldehid" N H3C CH2CH2O N S CH N P S NH2 tiamin (aneurin) B1-vitamin Az acetaldehidet szállító koenzim és prekurzor vitaminja Transfer coenzyme – acetaldehyde – TPP O HN C S ATP O ADP HOOC NH CH2 CH2 biotin (H-vitamin) CH2 CH2 N C CO 2 COOH S NH CH2 CH2 CH2 CH2 COOH karboxi-biotin A biotin keletkezése és formái P H3C H3C N piridoxin (B6-vitamin) H3C N N piridoxamin-foszfát (PAM) piridoxál-foszfát (PAL) 22 Az aminocsoportot szállító koenzim és prekurzor vitaminja NH2 N P O P O CH2 CH3 H CH–OH O C=O NH P CH2 P O P O H H C CH3 H OH CH3 O H C CH3 CH–OH C=O NH C=O NH C=O NH OH CH3 C CH3 CH–OH C=O NH CH2 CH2 S–C–CH3 CH2 O CH3–CO–SKoA acetil koenzim A "aktív ecetsav" COOH pantoténsav (régen B9-vitamin) SH Koenzim-A (újabban B5 vitamin) A koenzim-A különbözô formái Transfer coenzyme – acetyl group – coenzyme A COOH COOH S HS S liponsav H CH2OH CH2 ciszteamin H H 2,4-dihidroxiO 3,3-dimetilvajsav P CH2 -alanin CH2 CH2 O CH2 CH2 CH2 CH2 N N O CH2 N N N N O NH2 N SH dihidro-liponsav COOH HS S C CH3 O acetil-dihidro-liponsav A liponsav koenzim különbözô formái Transfer coenzyme – acetyl group – lipoic acid 23 CH2OH HO H3C piridoxin (B6-vitamin) CH2O– P HO H3C N CH2NH2 CHO CH2OH N piridoxál-foszfát (PAL) CH2O– P HO H3C N piridoxamin-foszfát (PAM) Az aminocsoportot szállító koenzim és prekurzor vitaminja Transfer coenzyme – amino group – PAL Lipids There are two types of lipid (apolar – fatty soluble) biomolecules. Simple lipids cannot be hydrolyzed by sodium hydroxide and complex lipids can be hydrolyzed by sodium hydroxide. The two main types of simple lipids are the fatty acids and the terpenes. Fatty acids (C16 and C18) are building blocks of complex lipids (neutral triglycerides and phospholipids). Saturated fatty acids are palmitate (CH3(CH2)14–COOH) and stearate (CH3(CH2)16–COOH). Unsaturated fatty acids are the unsaturated versions of stearate (C18): oleate, linoleate and linolenate. The essential linoleate (-6-fatty acid) and linolenate (-3-fatty acid) are known as PUFA (polyunsaturated fatty acids) or vitamins F. Formulas of oleate, linoleate and linolenate Terpenes can be derived from isoprene (methylbutadiene) CH2=C(CH3)–CH=CH2 (C5H8). Monoterpenes contain two (C5H8)2 (C10), diterpenes four (C5H8)4 (C20), triterpenes six (C5H8)6 (C30) and tetraterpenes eight isoprene units (C5H8)8 (C40). The branched end of isoprene is called the head (fej in Hungarian) and the other part is called the tail (láb in Hungarian). There are different variations for connecting the isoprene units. Most frequent are head-to-tail connections, while and tail-to-tail and head-to-head variations are rare. 24 H2C C CH CH2 CH3 izoprén fej fej-fej láb láb-láb fej-láb Az izoprén egységek kapcsolódási fajtái Different variations of connecting isoprene units: head-to-head, head-to-tail and tail-to-tail Only two representatives of polyisoprenoids are shown here: chloresterol as triterpene (C30) and -carotene as tetraterpene (C40). Triterpenes and tetraterpenes generally contain two chains with head-to-tail connection and these chains are CH3 CH3 CH3of the molecule. The chain of the connected in a tail-to-tail combination in the middle O triterpene squalene is cyclized to cholesterol. The ring system of cholesterol is called the sterane skeleton (withoutCHmethyl 3 CH3 groups gonane skeleton). Cholesterol can be the H starting material for different kinds of steroids – among them sexual hormones. CH2 CH3 OH Vitamin D formed from cholesterol by uv light plays an important role in calcification mentol kámfor limonén of cartilage and bone. Néhány monterpén képlete CH3 CH3 CH3 C A D CH3 B A C CH3 CH3 CH3 D B HO gonánváz szteránváz koleszterol cikloalkánok A gonánváz, a szteránváz és a koleszterol képlete Formulas ofCH gonane and sterane skeletons, and of cholesterol CH3 3 CH 3 CH3 CH3 CH3 Tetraterpene carotenoides areCHorganic pigments that are naturally occurring CH3 in h 3 CH3 C D the chloroplasts and chromoplasts of plants. There areCH two classes of carotenoides: 2 carotenes Aare hydrocarbons and xanthophylls contain oxygen. Because of B polyconjugated double h bond system, carotenoids can absorb light energy for use in HO HO photosynthesis, and as antioxidants they protect chlorophyll from photodamage. D3-vitamin (kolekalciferol) Antioxidants can eliminate free radicals by reduction. In humans -carotene and other carotenoids can be converted to retinol (vitamin A) by an oxidative CH3splitting. Retinal CH CH CH CH 3 3 3 synthesized3 from retinol is essential for vision. -jonon CH3 CH3 CH3 CH3 ox. -karotin CH3 CH3 CH3 CH2OH CH3 CH3 CH3 -jonon CH3 CH3 CH3 CH3 CH3 25 The conjugated double bond system of -carotene Complex lipids have generally ester group(s) (sometimes carboxamides) therefore they can be attacked by nucleophilic reagents e.g. sodium hydroxide. There are four categories of complex lipids: fruit esters, waxes, neutral triglycerides (fats and oils) and phospholipids (membrane lipids that can form the lipid bilayers of cell membranes). Fruit esters (synthesized from short-chained carboxylic acids and short-chained alcohols) are flavour components of fruits, e.g. aroma of pineapple is methyl butyrate (CH3CH2CH2–COOCH3). Waxes (synthesized from long-chain carboxylic acids and long-chain alcohols) are not only water-repellent materials on the surface of leaves and fruits but bees use beeswax (H3C(CH2)14–COO(CH2)29CH3 myricyl palmitate) to form the walls and caps of the comb. Neutral triglycerides (triacylglycerols)are triesters of glycerol with fatty acids (C16-C18)- They serve as are highly concentrated energy stores in fat cells (adipose cells), as water-repellent materials (e.g. on the skin) and as heat-insulators in humans and animals. Fats are solid and their fatty acid parts are palminate, stearate and oleate. Oils are liquids and their major fatty acid part is linoleate. Surfactant (detergent) soaps (sodium salts of fatty acids) can be produced by the hydrolysis of fats with sodium hydroxide. RCOOCH2 RCOOCH + 3 NaOH 3 R C CH2 O H O + CH O H O Na szappan RCOOCH2 CH2 O H glicerol A szappanfôzés összfolyamata Hydrolysis of triglycerides by sodium hydroxide Surfactant molecules (o) contain both polar (o) and apolar () parts that are suitable for selective adsorption. In this way the apolar surface of the fat (zsír in Hungarian) can be changed to quasi-polar and fats can produce an emulsion in water. O O CH3 (CH2)14–C O nátrium-palmitát O O Na zsír O poláros apoláros O O O O A mosóhatás O O poláros felület 26 P Selective adsorption of surfactants to fat P P P There are different types of phospholipids and type of phosphoglycerides P is P P their major class. Phosphoglycerols can be derived from phosphatidate (phosphatidic acid) that is a phosphate ester of diacylglycerols producing phosphatidyl P ethanolamines, phosphatidyl serines, and phosphatidyl cholines. The acyl groups are from fatty acids (C16-C18). Phosphoglycerides are surfactants and lecitines are the major component of animal cell membranes (in plants the major membrane lipid components are glycolipids). RCOOCH2 RCOOCH2 RCOOCH RCOOCH CH2 O CH2–O– P foszfatidsavak NH2 O–CH2–CH szerin-kefalinok COOH P RCOOCH2 RCOOCH CH2 O RCOOCH2 RCOOCH CH2 O apoláros rész P O–CH2CH2 NH2 kolamin-kefalinok P O –CH2CH2 N(CH3)3 poláros rész lecitinek A foszfolipidek származtatása a foszfatidsavból Formation of different kinds of phosphoglycerols: phosphatidyl ethanolamines (szerin-kefalinok in Hungarian), phosphatidyl serines (szerin-kefaninok in Hungarian) and phosphatidyl cholines (lecithines) (lecitinek in Hungarian) from phosphatidate There many biomolecules containing phosphate in ester (e.g. phosphatidate acid) or anhydride (e.g. ATP) form, therefore their abbreviations are used. Abbreviations of phosphates in esters and anhydrides Membrane transport processes A membrane is a layer of material which serves as a selective barrier between its two sides, and remains impermeable to specific particles, molecules, or substances. The structure of membranes can be illustrated by the fluid mosaic model. Through the bimolecular layer surfactant phospholipids membrane-integrated (transmembrane) 27 proteins can make passage possible for specific molecules. Surface proteins can bind different regular molecules as receptors. áthatoló fehérje felületi fehérje irányított, bimolekuláris foszfolipid réteg A membránok felépítése Fluid mosaic model of the structure of membranes – place of a transmembrane (membrane-integrated) protein (áthatoló fehérje in Hungarian) and a surface protein (felületi fehérje in Hungarian) The scheme of an Eukaryotic cell (from book of Ádám and Fehér) Some compounds are allowed to pass through the membrane, whereas others are retained. The driving force for the passage is a difference in the concentrations of 28 the molecule on the two sides of the membrane and the molecules pass from the higher to lower concentration without the investment of energy (called passive diffusion). The transfer can be carried out in a simple way for small molecules (e.g. water) or by a facilitated diffusion by special transfer molecules. There are different kinds of passive diffusion with the aid of transmembrane proteins. In symport transport there are two molecules bound to the same part of membrane and the concentration gradient from higher to lower concentration is valid for the sum of the concentrations of both molecules. In antiport transport there are two molecules bound to the opposite sides of membrane and the concentration gradient from higher to lower concentration has to be valid for both of them. In this way the position of two molecules is exchanged during the passive diffusion. The name of this kind of proteins is translocases. Passage from a lower to a higher concentration needs a change in the conformation of the integrated protein by energy investment (by the hydrolysis of a macroerg bond). The process is similar to the antiport because generally the position of two molecules is exchanged. Nucleic acids Nucleic acids are biopolymers consisting of nucleotide units connected by 3’,5’-phosphodiester bonds. A nucleotide unit contains a nucleic acid base either containing a pyrimidine ring (thymine (T) and cytosine (C) for DNA or uracil (U) and cytosine (C) for RNA) and or a purine skeleton (adenine (A) and guanine (G) for both DNA and RNA). The connection of sugars i.e. D-2’-deoxyribose for DNA and Dribose for RNA to nucleic bases is shown by an arrow (). This point is 3-N for pyrimidine and 9-N for purine bases. Nucleic acids are N-glycosides. The numbering of sugar is distinguished from that of the base with comma. The phosphate is connected to the 5’-hydroxyl group of sugars forming ester group. 1 6 HN O N H timin (DNS) H uracil (RNS) O NH2 7 5 N 6 1 N N N H citozin 3 4 N 9 H2N ) feltüntetésével 5' 5' HOCH2 O 4' H 3' 1' OH H 2' H OH OH - D-ribofuranozid HOCH2 4' H O H 3' 1' OH 2' H H OH H - D-2'-dezoxiribofuranozid A nukleinsavak építô elemei The structure of nucleoside units N guanin purin bázisok a kapcsolódási hely ( H N H N HN adenin pirimidin bázisok pentózok O 8 2 4 3N CH3 HN 5 2 O NH2 O O N H 29 The unit consisting only of a nucleic base and sugar is called nucleoside. The name for a nucleoside monophosphate is nucleotide. The name of nucleosides are uridine, thymidine, cytidine, adenosine, guanosine. Nucleotides of DNA are distinguished from RNA by the abbreviation of ‘deoxy’: UMP, dTMP, CMP, dCMP, AMP, dAMP, GMP, dGMP. ROCH2 O N ROCH2 O H CH3 HN HN O H H H OH O ROCH2 H N O H H OH H R = H dezoxitimidin (dT) R = H uridin (U) R = P uridin monofoszfát R = P dezoxitimidin-monofoszfát (dTMP) (UMP) OH N N O H H NH2 O O H H OH H Q R = H, Q = H dezoxicitidin (dC) R = P O P , Q = OH citidin-difoszfát (CDP) Néhány pirimidinvázas nukleozid, nukleotid, nukleozid-difoszfát és nukleozid-trifoszfát képlete The formulas of some nucleosides, nucleotides, nucleoside diphosphates and nucleoside triphosphates containing a pyrimidine ring The formulas of some nucleosides, nucleotides, nucleoside diphosphates and nucleoside triphosphates containing a purine skeleton The nucleotide units are connected by 3’, 5’-phosphodiester bonds. In each unit there is one acidic hydrogen atom at the phosphate part therefore the biopolymer 30 itself is called nucleic acid. The 5’ terminal of the polymer chain (strand) contains a phosphate ester and its 3’ terminal contains a hydroxyl group. The 3’,5’-phosphodiester bond in the polynucleotide chain (R=H DNA, R=OH RNA) P O CH2 O 5'-láncvég H H H 3' O B bázis Bázis H P 5' OH 3' cukor egység (ribóz, dezoxiribóz) A nukleozid egység sematikus ábrázolása P 5' O CH2 H H A Bázis O H H O OH 3'-láncvég A nukleinsavak elsôdleges szerkezete (a P a megfelelô foszforsav egységet jelenti) 5'-láncvég P 5' G 3' P 5' 3' 3'-láncvég A nukleinsav szekvencia sematikus ábrázolása 5'-láncvég AGC ......... 3'-láncvég A nukleinsav szekvenciájának legegyszerûbb ábrázolása A polinukleotid lánc ábrázolási lehetôségei Various modes to represent the polynucleotide chain In DNA two of antiparallel polynucleotide strands form a double helical structure described by the Watson - Crick Model. On the surface of helix is the chain containing the sugar and phosphate. Inside the helix two nucleic bases (a pyridine and a purine base) form a pair connected by hydrogen bonds (two for T-A and three for CG) and the character of one base determines the another base, therefore they are called complementary base pairs. 31 O H3C H N N R N N N O N R T O NH2 H2N R A N H N N N N N O C R H2N G A komplementer bázispárok Complementary pairs of bases.) In most of the cases RNA contains only one strand, but it can form a double helix with a DNA during its biosynthesis. There can be a double helical section in an RNA chain when it contains an antiparallel complementary sequence – this folded form is called palindromic structure (e.g. in tRNA). There can be double palindromic structures in DNA, as well. Nucleic acids are in the nucleus of the cell, therefore the double helix needs to assume various more compact forms. The double helix of DNA is wrapped around clusters of histones (small proteins with a basic character) by lefthanded superhelical turns to form nucleosomes, which are coiled to form solenoids that are further compact formations for DNA. Solenoids are able to become increasingly even more packed formations – these are chromosomes. Scheme of the Watson-Crick Model of DNA The biological function of DNA is to preserve genetic information for the biosynthesis of proteins. Different types of RNA include tRNA (transfer), mRNA (messenger), rRNA (ribosomal) and snRNA (small nuclear) provide the conditions for the biosynthesis of proteins (translation). Details are given in the section of the metabolism of the biomolecules of the genetic information. mRNA transports information from DNA about a protein sequence to the ribosome (the site of protein synthesis in the cell); tRNA transports amino acids to the ribosome connected to its 3’ terminal as esters; 32 rRNA is the catalytic component of the ribosome containing not only RNA but several proteins; snRNA makes ripening of different types of RNA by splicing. Macroerg bonds The phosphoric acid anhydride (pyrophosphate) derivatives of nucleotides are the nucleoside diphosphates (NDP) and nucleoside triphosphates (NTP). Their anhydride bonds (one in NDP and two in NTP) are called macroerg bonds (they have a high phosphoryl-transfer potential) because their synthesis requires energy while their hydrolysis generates an energy of about 30,6 kJ/mol. The most important NTP is adenosine triphosphate. There are different types of macroerg bonds. They are formed from an acid and a compound with acidic character. Anhydrates can be synthesized from two molecules of phosphates (phosphoric acid anhydrides e.g. ATP) or from a carboxylate and a phosphate (mixed anhydrides e.g. glycerate 1,3-bisphosphate). There are other compounds with acidic character that can form esters with an acid. An ester from phosphate and an enol (e.g. phosphoenolpyruvate – PEP) or a thiolester from a carboxylate and a thiol (e.g. acetyl coenzyme A – acetyl-CoA) (H3C–COSCoA) contain also macroerg bonds. NH2 N N P O P O N N O P O CH2 H H H H OH OH adenozin-trifoszfát (ATP) Az ATP képlete Adenosine triphosphate (ATP) 33 O a) savanhidridek – foszforsavanhidrid (pl. ATP) O P O P O H O O vegyes savanhidrid O pl. P O P O O H P O C O O C O P O O H C O H CH2 O O H P glicerinsav 1,3-difoszfát b) különleges észterek – enolészter pl. PEP (foszfo-enol-piruvát) COOH CH O – tiolészter pl. acetil-koenzim-A CH3 P CH2 O C S KoA A makroerg kötések Compounds containing macroerg bonds: phosphoric acid anhydride, mixed anhydrides e.g. glycerate 1,3-bisphosphate, enolester e.g. phosphoenolpyruvate, thiolester e.g. acetyl coenzyme A Metabolism The whole range of organic reactions of biomolecules in the living organisms are called metabolism. Primary metabolism – the metabolism of biomolecules. Phases of primary metabolism: Catabolism – oxidative degradation of biomolecules combined with the generation of energy in form of macroerg bonds (e.g. ATP). The intermediates of catabolism are the starting materials of anabolism. The terminal products of the catabolism are CO2 and H2O. Anabolism – reductive biosynthesis of biomolecules by means of the energy produced during catabolism. In autotrophic plants glucose molecules are synthesized from CO2 and H2O by the energy of the light. The other biomolecules are synthesized from ammonia and the metabolites of glucose degradation. The starting materials of anabolic reactions of heterotrophic living organisms (animals and human beings) derive from the oxidative degradation of the nutritive materials (foods). Secondary metabolism: the metabolism of different molecules of the living organisms which are generally needed for their functioning. Secondary metabolites are synthesized from the different intermediates of biomolecules. The most important types of secondary metabolites are coenzymes, regulating (e.g. hormones), attracting (e.g. the sweet sucrose, the fruit esters as scent agents etc.) and repelling agents (e.g. alkaloids and toxins). The processes of basic biochemistry take place inside the cell: in the cytosol (cytoplasm), mitochondria and ribosomes – and we are focused on human biochemistry (except photosynthesis). Only the metabolism of nutritive materials is 34 discussed because the metabolism of nucleic acids is in nucleus of the cell and its role is peripheral in the nutrition. Scheme of the biosynthesis of biomolecules Catabolism The first phase of the degradation of nutrients (and nucleic acids) is the hydrolysis of the combined functional groups of biopolymers in the cytosol. Then in the second phase (at first in the cytosol then in the mitochondria) the oxidative degradation of the intermediates (sugars – especially glucose, amino acids, fatty acids and glycerol) leads to a synthesis of a common intermediate acetyl coenzyme A (H3C-COSCoA with a macroerg thiolester) (the intermediates of some amino acids are the members of citric acid cycle). In the third phase (in the mitochondria) oxidative degradation of acetyl coenzyme A by oxygen to CO2 (citric acid cycle) and H2O (with the form of macroerg bonds) (terminal oxidation – respiratory chain) takes place. 35 Biomolecule Polysaccharides Enzyme Glycosidases/ Phosphorylases Proteases/Peptidases Lipases Intermediate Sugars/ Sugar phosphates Amino acids Fatty acids and glycerol Proteins Neutral triglycerides (triacylglycerols) Nucleic acids Nucleases Nucleotides The first phase of the degradation of biomolecules In the hydrolytic phase of the degradation of biopolymers different types of hydrolase enzymes take part. Exohydrolase enzymes start the hydrolysis at one end of the biopolymers (e.g. with polysaccharose chains at the non-reducing end; in proteins amino peptidase enzymes at N-terminal and carboxypeptidase enzymes at C-terminal). Endohydrolase enzymes start hydrolysis in the middle of the biopolymers (in polysaccharide chains in a random way, in proteins between special amino acid units: pepsin – before aromatic amino acids; trypsin – after basic amino acids; chymotrypsin – after aromatic amino acids). Hydrolase enzymes often need an activating step (phosphorylation or hydrolysis) before action. Hydrolases of amylose (one of the two components of starch): -amylase is an endohydrolase; -amylase an exohydrolase; that starts the hydrolysis of amylose at the non-reducing end splitting off maltose molecules; maltase hydrolyzes maltose to two glucose molecules. Cellulose is hydrolyzed by cellulase (only in certain microorganisms) to cellobiose, that is hydrolyzed by -glucosidase to glucose. Lactose is hydrolyzed by lactase (-galactosidase) to glucose and galactose, sucrose is hydrolyzed by invertase to glucose and fructose. The addition of a phosphate group from an inorganic phosphate (phosphoric acid) to a substrate can be catalyzed by phosphorylases. The name ‘phosphorylase’ is generally used for glycogen phosphorylase that catalyzes the release of glucose-1phosphate from the reducing end of glycogen molecule with an inorganic phosphate (Pi). Glucose-1-phosphate is converted to glucose-6-phosphate to enter glycolysis. The details of this reaction are given in the section of biosynthesis of polysaccharides. The degradation of carbohydrates to acetyl coenzyme A After the hydrolysis of carbohydrates the product is mostly glucose. Other sugars (except fructose) can be formed directly or in NDP-sugar phase to glucose (e.g. galactose). The first phase of the degradation of glucose is glycolysis from glucose (C6 stage) to pyruvate (pyruvic acid) (C3 stage) in the cytosol. Pyruvate is a common starting material for the two kinds of anaerobic degradations (alcoholic and lactic acid fermentations) of glucose in the cytosol and its aerobic degradation (at first to acetyl coenzyme A then to carbon dioxide and water) in the mitochondria. Different biochemical processes are characterized not only by their participants but also by their stoichiometry. Stoichiometry (stoichiometry of reactions) is the quantitative relationships of the reactants and products in a balanced chemical reaction. Glycolysis The steps of glycolysis are reversible – except three of them. The first and the third reactions are catalyzed by kinases (hexokinase and phosphofructokinase – PFK). A kinase (transferase) can phosphorylate a molecule coupled with the hydrolysis of a 36 macroerg phosphoric acid anhydride bond of a NTP (mostly ATP) but the product has not a macroerg bond (in the case of sugars the product is a phosphate ester of a sugar), therefore this kind of reaction is irreversible. The details of the third irreversible step of glycolysis will be given later. The phosphorylation of glucose to glucose 6-phosphate by hexokinase is often called the ‘activation of glucose’. It is not a real activation step, because – as it was mentioned earlier – this ester does not contain a macroerg bond. But the phosphoric acid unit of sugars can help the formation of a connection between sugars and enzymes by ionic interactions. The isomerization of glucose 6-phosphate to fructose 6-phosphate catalyzed by an isomerase is a double oxo-enol-oxo tautomerism that was presented earlier. The last step of C6 stage of glycolysis is the synthesis of fructose 1,6bisphosphate from fructose 6-phosphate catalyzed by PFK. PFK is the key enzyme in the control of glycolysis because its activity can be inhibited by ATP that is the end product of oxidative phosphorylation part of the respiratory chain. In the case of high ATP concentration glycolysis is stopped but at low ATP concentration glycolysis is started. This control system is called feedback and its mechanism is called allosteric mechanism. There are also other regulating agents of PFK. In the second stage (C3) of glycolysis the cleavage of fructose 1,6-bisphosphate catalyzed by aldolase results in a mixture of glyceraldehyde 3-phosphate and dihydroxyacetone phosphate (DHAP). Glyceraldehyde 3-phosphate is on the direct pathway of glycolysis. The conversion of DHAP to glyceraldehyde 3-phosphate is catalyzed by an isomerase. It is noticed that at equilibrium 96% of triose phosphate is DHAP. 37 Glycolysis and the possibilities for the further degradation of pyruvate The oxidative step of the glycolysis is the formation of 1,3bisphosphoglycerate (its earlier name is 1,3-diphosphoglycerate) containing a macroerg mixed anhydride bond from glyceraldehyde 3-phosphate catalyzed by glyceraldehyde 3-phosphate dehydrogenase with a covalent catalysis. The coenzyme is NAD that is reduced to (NADH+H). This step is one of the two energy producing steps of glycolysis. In the course the synthesis of the macroerg mixed anhydride bond during the oxidation at first a macroerg thiolester intermediate is formed in the enzyme-substrate complex (ES) that is attacked by an inorganic phosphate. The stoichiometry of glycolysis The energy of 1,3-bisphosphoglycerate is converted to the synthesis of an ATP molecule from ADP and an inorganic phosphate (Pi) accompanied by the formation of 38 3-phosphoglycerate. The rearrangement of 3-phosphoglycerate to 2-phosphoglycerate is in fact a combination of two transfer reactions by means of a cofactor 2,3bisphosphoglycerate, and catalyzed by a mutase that is a transferase. During the elimination of a water molecule from 2-phosphoglycerate to phosphoenolpyruvate (PEP) a macroerg thiolester bond is formed (the catalyst is enolase). The energy of this thiolester bond is converted to the energy of a phosphoric acid anhydride (ADP+PiATP), and enolpyruvate is transformed immediately to pyruvate by an irreversible oxo-enol tautomerism (the catalyst is pyruvate kinase). This reaction is the third irreversible step of glycolysis because direct conversion of pyruvate to enolpyruvate is impossible it can be carried out only in a roundabout way (see gluconeogenesis). The entrance of fructose to the glycolysis Fructose can enter the glycolysis in an alternative way called fructose 1phosphate pathway because the affinity of hexokinase is about twenty times lower than that of glucose, and the glucose concentration is generally higher in cells than that of fructose. The phosphorylation of fructose by ATP to fructose 1-phosphate is catalyzed by fructosekinase and the elimination of fructose 1-phosphate to DHAP and glyceraldehyde is catalyzed by fructose 1-phosphate aldolase. DHAP is the intermediate of the glycolysis but glyceraldehyde can enter the glycolysis only after a phosphorylation glyceraldehyde 1-phosphate by ATP catalyzed by triose kinase. It is noticed that there are some tissues (e.g. adipose tissue) in which hexokinase can phosphorylate fructose to fructose 6-phosphate because of the high fructose concentration in them. The possibilities for the further degradation of pyruvate According to the stoichiometry of glycolysis during the degradation of one molecule of glucose to two molecules of pyruvate one molecule reduced coenzyme (NADH+H) and two molecules of ATP (exactly two macroerg bonds) are formed. In 39 a fermentation process only the reduced coenzyme is used for the reduction of pyruvate (to ethanol and CO2 in alcoholic fermentation and to L-lactate in lactic acid fermentation). That means that the benefit of the combination of glycolysis and fermentation (that is the anaerobic degradation of glucose) is only 2ATP/glucose. The stoichiometry of the lactic acid and alcoholic fermentations of glucose Ethanol is formed from pyruvate in yeast and several other microorganisms in two steps. The decarboxylation of pyruvate to acetaldehyde is catalyzed by pyruvate decarboxylase then the reduction of acetaldehyde by (NADH+H) formed in glycolysis to ethanol is catalyzed by alcohol dehydrogenase. L-Lactate from pyruvate can be formed by the reduction of (NADH+H) not only in certain microorganisms but in the muscle of animals and human beings. It is a fast but ‘non-economic’ method for releasing of energy during the degradation of glucose. There is another possibility for the further degradation of pyruvate. This takes place in mitochondria and the result of this complex process using oxygen (6 O2/glucose) is 36-38ATP/glucose. This process is called the aerobic degradation of glucose. After the irreversible penetration of pyruvate to the mitochondria the first step of this complex process is the formation of acetyl coenzyme A from pyruvate catalyzed by pyruvate dehydrogenase complex. The structure of mitochondria Mitochondria have an outer membrane (in Hungarian külső membrán) and a highly folded inner membrane (in Hungarian belső membrán) with a large surface. The intermembrane space is between the folds of the membranes. The inside of the mitochondria is the matrix that is bound by the inner membrane. The outer membrane is permeable to small molecules and ions. In contrast penetration through the inner membrane is only possible by membrane transport processes of transmembrane proteins. The site of the dehydrogenation of pyruvate, the citric acid cycle and the fatty acid oxidation occurs in the matrix. The site of terminal oxidation (respiratory chain) is in the inner membrane. Membranes of mitochondria 40 The oxidative decarboxylation of pyruvate to acetyl coenzyme A catalyzed by pyruvate dehydrogenase multienzyme complex The oxidative decarboxylation of pyruvate to acetyl coenzyme A with a macroerg thiolester bond is catalyzed by the pyruvate dehydrogenase multienzyme complex. Coenzyme is NAD. The reaction catalyzed by pyruvate dehydrogenase There are different steps, three enzyme subunits (pyruvate dehydrogenase, dihydrolipoyl transacetylase, dihydrolipoyl dehydrogenase), three coenzymes (TPP, coenzyme A, NAD) and two prosthetic groups (lipoic acid, FAD) participating in the oxidative decarboxylation of pyruvate. The first step is the decarboxylation of pyruvate combined with the synthesis of active acetaldehyde. Active acetaldehyde is derivative of coenzyme TPP that seems to be a secondary alcohol but really it can react as a carbonyl compound (it can be oxidized to acyl group), because the carbon atom between N+ and S is a reactive carbon atom with acidic (dissociable) proton. Active acetaldehyde is oxidized by lipoic acid to an acetyl group that is immediately connected to one of the thiol groups of the reduced lipoic acid (acetyl dihydrolipoic acid with a macroerg thiolester bond). The acetyl group is transferred to a coenzyme A and lipoic acid is regenerated from dihydrolipoic acid by a cascade reductive system containing FAD as prosthetic group and the coenzyme NAD producing (NADH+H). The product acetyl coenzyme A with a macroerg thiolester bond is the starting material of the citric acid cycle. 41 The stoichiometry of the reaction catalyzed by pyruvate dehydrogenase The degradation of lipids to acetyl coenzyme A Degradation in the cytosol After the hydrolysis of complex lipids by lipases the ways of the oxidative degradation of fatty acids and glycerol are different. The oxidative degradation of alcohols is related to the degradation of carboxylic acids Glycerol 3-phosphate (its earlier name is -glycerol phosphate) is formed by the phosphorylation of glycerol with ATP catalyzed by a kinase. Glycerol 3-phosphate can be oxidized by NAD and the product is DHAP which is an intermediate of glycolysis. The decomposition of triglycerides and glycerol Fatty acids are activated before oxidative degradation by transformation to acyl coenzyme A with the aid of a macroerg thiolester bond. The first step is the reaction of the fatty acid and ATP to acyl-AMP that contains a macroerg mixed acid anhydride bond. The reaction needs the energy of another macroerg phosphoric acid anhydride provided by the hydrolysis of the by-product pyrophosphate to two molecules of phosphoric acid. Acyl coenzyme A is formed from the reaction of acyl-AMP and coenzyme A. 42 The activation of fatty acids to acyl coenzyme A Acyl coenzyme A enters the matrix of mitochondria in form of acyl carnitine formed in a reaction catalyzed by carnitine acyltransferase I at the cytoplasmic side of the inner membrane then it takes part in an antiport membrane transport process against L-carnitine that transfers the acyl group to coenzyme A in a reaction catalyzed by carnitine acyltransferase II at the matrix side of the inner membrane The membrane transport of fatty acids from cytosol to the matrix of mitochondria -Oxidation of fatty acids to acetyl coenzyme A molecules Degradation in the mitochondrial matrix – -oxidation The oxidative degradation of the saturated acyl CoA named -oxidation contains a recurring sequence of four reactions and the first three of these reactions closely resembles the last reactions (from succinate to oxaloacetate) of the citric acid cycle. The coenzyme (prosthetic group) of the oxidation of acyl CoA to enoyl CoA containing a trans double bond between C-2 and C-3 is FAD. The coenzyme of the second oxidation step after the hydration of enoyl CoA to L-3-hydroxyacyl CoA is 43 NAD and the product is 3-ketoacyl CoA (its earlier name is -ketoacyl CoA; that is the origin of the name of the degradation). The last step of the -oxidation is the cleavage 3-ketoacyl CoA by the thiol group of a molecule of coenzyme A resulting in a molecule of acetyl-CoA and an acyl CoA shortened by two carbon atoms. The reaction is called thiolysis and is catalyzed by -ketothiolase (or simply thiolase). At the end of -oxidation each carbon atom of the original fatty acids is transformed to acetyl coenzyme A. At the end product of the -oxidation of fatty acids having an odd number of carbon atoms (these are minor species) is, instead of acetyl-CoA, one molecule of propionyl CoA. This is converted to succinyl CoA (an intermediate of citric acid cycle) in two steps (a carboxylation followed by an intramolecular rearrangement). Unsaturated fatty acids contain cis double bonds. In the case of one double bond (oleate) an isomerization of cis double bond to a trans double bond is catalyzed by an isomerase. When there is another double bond in the fatty acid (linoleate and linolenate) the result of the hydration step is a D-hydroxyl derivative that is converted to L- hydroxyl derivative by an epimerase. The oxidative degradation of terpenes to acetyl-CoA is carried out by similar processes as described for -oxidation. Degradation of L-amino acids to acetyl coenzyme A or intermediates of citric acid cycle After the hydrolysis of proteins by different kinds of peptidases and proteases there are four levels of the degradation of amino acids. Three of them (transamination, oxidative deamination of glutamate, urea cycle) are connected with the removing of the -amino groups (the details are given later) and produce -keto carboxylic acids while the fourth level is the oxidative degradation of -keto carboxylic acids to pyruvate (Ala, Thr, Gly, Ser, Cys, Trp), acetyl-CoA (Trp, Leu, Ile), acetoacetyl-CoA (Phe, Tyr, Trp, Leu, Lys), -ketoglutarate (Glu, Gln, His, Arg, Pro), succinyl CoA (Met, Val, Ile), fumarate (Tyr, Phe, Asp) and oxaloacetate (Asp, Asn). There are also other direct degradation modes (oxidative deamination or elimination of an ammonia or water molecule). Transamination In transamination reactions (catalyzed by aminotransferases) the amino group of amino acids is transported to -ketoglutarate (that is an intermediate of citric acid cycle) – the products are -keto carboxylic acids (from the amino acid) and glutamate (from -ketoglutarate) by means of the coenzyme pyridoxal phosphate (PAL) that is in fact a prosthetic group. The first step of this process is the formation of aldimines (Schiff bases). Then by the rearrangement of aldimines ketimines are synthesized by an aldimine-ketimine tautomerism. -Keto carboxylic acids and pyridoxamine phosphate (PAM) are produced by the hydrolysis of ketimines. The regeneration of PAM to PAL is carried out by a transformation of -ketoglutarate to glutamate. In this way the amino groups from all of the amino acids are transferred to glutamate molecules. Aldimines are the starting materials of biogenic amines by decarboxylation followed by hydrolysis. Aldimines from some amino acids (e.g. serine) transfer their side chain to a tetrahydrofolate (THF) coenzyme producing glycine from the amino acids. 44 Transamination and oxidative deamination of glutarate Other reactions from aldimines derived from amino acids Oxidative deamination of glutarate Glutarate molecules produced from amino acids are oxidized to an imino acid derivative which is hydrolyzed to -ketoglutarate and ammonia. The reaction is catalyzed by glutamate dehydrogenase accompanied by NAD (NADH+H) conversion. In plants the reverse process (reductive amination) of this reaction gives a possibility for the assimilation of ammonia using (NADPH+H). The urea cycle A part of ammonia (as NH4+) is used for the biosynthesis of nitrogen derivatives. Since ammonia is a toxic for animals and human beings therefore surplus 45 ammonia is converted to urea (in Hungarian karbamid) in the urea cycle (another name is ornithine cycle) connected to the biosynthesis of arginine (Arg). The urea cycle and its connection with the citric acid cycle The stoichiometry of the urea cycle From five reactions of the urea cycle two are mitochondrial and three cytoplasmic. In the mitochondria CO2, NH3 and 2ATP produce carbamoyl phosphate, 2ADP and Pi. Then carbamoyl phosphate reacts with ornithine and produces citrulline and Pi. The cycle is continued in the cytosol. The second amino group of urea is derived from aspartate that reacts with citrullin by means of converting an ATP to AMP and PPi and the product is argininosuccinate. From argininosuccinate arginine and fumarate are formed in an elimination reaction. Aspartate can be recovered from fumarate in citric acid cycle (fumarate malate oxaloacetate followed by a transamination to aspartate). Arginine can be hydrolyzed to ornithine (being the final step of the cycle) and urea (in two steps with isourea as intermediate). According to the stoichiometry of the urea cycle the synthesis of an urea molecule needs the energy of three macroerg bonds. Degradation of the common intermediate (acetyl coenzyme A) to CO2 and H2O 46 During the further degradation of acetyl coenzyme A the formation of carbon dioxide and water molecules takes place in separate reactions. These biochemical processes are localized in mitochondria. In the citric acid cycle two molecules of carbon dioxide and a CoA molecule are formed from one acetyl-CoA molecule, meanwhile four reduced coenzymes: 3 (NADH+H), FADH2 and one macroerg bond (GDPGTP) are produced in the mitochondrial matrix. In the respiratory chain (terminal oxidation) in the inner membrane of mitochondria the reduced coenzymes reduce an oxygen molecule to water by electron transport and at the same time the energy of nutrients are built into macroerg bonds of ATP (ADP + Pi ATP). Finally all of the carbon atoms of the nutrients are oxidized to CO2, at the same time the hydrogen (formed during the regeneration of coenzymes) and oxygen atoms of nutrients produce water molecules. The stoichiometry of the citric acid cycle Citric acid cycle The other names of this cycle are: tricarboxylic acid cycle (TCA cycle), Krebs cycle, or sometimes Szentgyörgyi-Krebs cycle. Acetyl-CoA reacts with the staring material (oxaloacetate) of the cycle in an addition reaction catalyzed by citrate synthase followed by the hydrolysis of the intermediate (citryl CoA) to citrate and CoA. In this reaction the nucleophilic attack of the acetyl group at the carbonyl group of oxaloacetate is carried out by the methylen part of the enol tautomer that is temporarily formed in the reaction under the influence of the strong hydrogen bond of a His of the enzyme. Nucleophilic attack of the enol tautomer of acetyl-CoA at the carbonyl group of oxaloacetate It was proved by a labeled compound that in the further reaction citric acid reacts as an asymmetric molecule because of the asymmetric association with the enzyme. That is the reason why the carbon dioxide molecules are derived from carbon atoms of oxaloacetate and the regenerated oxaloacetate contains the carbon atoms of acetyl-CoA. The problem is shown by the illustration of both forms of oxaloacetate in the scheme of citric acid cycle. The next step of the citric acid cycle is a rearrangement of citrate to L-isocitrate that is carried out by a combination of a water elimination followed by water addition. The enzyme is a lyase (aconitase after the name of the unsaturated 47 intermediate cis-aconitate). The products of the oxidation of isocitrate combined with decarboxylation are -ketoglutarate, carbon dioxide and (NADH+H) in a reaction catalyzed by isocitrate dehydrogenase. The second carbon dioxide molecule is produced in the oxidative decarboxylation of -ketoglutarate to succinyl CoA with a macroerg thiolester bond catalyzed by -ketoglutarate dehydrogenase multienzyme complex. The structure of this enzyme is similar to that of pyruvate dehydrogenase using three coenzymes (TPP, coenzyme A, NAD) and two prosthetic groups (lipoic acid, FAD). The reduced coenzyme is again (NADH+H) and CoA comes from the acetyl-CoA. The citric acid cycle Succinyl CoA is hydrolyzed by succinate CoA synthetase that is a ligase. The energy of the macroerg bond is transmitted to a GDP+Pi GTP conversion. The next three steps of the cycle are similar to the first steps of -oxidation of fatty acids. Succinate is oxidized to fumarate by succinate dehydrogenase accompanied by a FAD FADH2 conversion. In a hydration reaction catalyzed by fumarase fumarate is converted to L-malate which is oxidized to oxaloacetate (the staring material of the cycle) by malate dehydrogenase. The reaction is accompanied with a NAD (NADH+H) conversion. The enzymes of the citric acid cycle are working as large enzyme complexes attached to the inner membrane of mitochondria. The cycle is regulated by feedback. The essential regulating effect is the ATP/ADP ratio but other intermediates play also an important role. The citric acid cycle is in close cooperation with other biochemical processes. Its intermediates participate as starting materials in many biochemical 48 reactions therefore it is often called ‘the pool of biointermediates’. Glycolysis can work under both anaerobic and aerobic condition while the citric acid cycle can work only in an aerobic mode, because the regeneration of the reduced coenzymes takes place in the terminal oxidation which needs the presence of oxygen. There are many bacteria and plants that can synthesize glucose from acetylCoA in a biosynthetic process called the glyoxylate cycle. As far as the synthesis of isocitrate the steps of glyoxylate cycle are the same as those of the citric acid cycle. The details are given later. Terminal oxidation (Respiratory chain) The name ‘terminal oxidation’ means that this metabolic pathway is the last oxidation step in the catabolic pathway (in the oxidation of the carbon atoms of biomolecules or other organic compounds). Now this name is preferred. The name ‘respiratory chain’ means that this metabolic pathway can be connected directly to respiration, the direct oxygen consumption of living organisms. This name is preferred in medical biochemistry. There are two partial processes in terminal oxidation. The electron transport chain is a special chain to transfer electrons from a higher-energy molecule (the donor) with lower standard oxidation-reduction potential (E’0) to a lower-energy molecule (the acceptor) with higher standard oxidation-reduction potential in order to regenerate reduced coenzymes. Oxidative phosphorylation is the metabolic pathway that uses energy released by the oxidation of nutrients to produce macroerg bonds (ADP+Pi ATP). Instead of terminal oxidation the name ‘oxidative phosphorylation’ is often used for the whole metabolic pathway. The electron transport chain The enzyme complexes of the terminal oxidation are in the inner membrane of mitochondria. There are three large enzyme complexes (NADH-Q reductase, cytochrome reductase and cytochrome oxidase) and two small ones (coenzyme Q and cytochrome c). Coenzyme Q (ubiquinone) can move between the two walls of the inner membrane, therefore it can receive hydrogen atoms from the intermembrane space through the outer part of the inner membrane of mitochondria (e.g. glycerate 3phosphate redox shuttle). NADH-Q reductase contains as prosthetic group FMN. The driving force of electron transport is the electron-transfer potential of reduced coenzymes to oxygen. The direction of the electron transport is determined by the standard oxidation-reduction potential values (E’0) of the participants. The direction is from negative to positive values of the standard oxidation-reduction potentials. It means that the direction is from participants with high-potential electrons to those with low-potential electrons. The standard oxidation-reduction potentials are given in V (volts). The standard oxidation-potential of H:H2 is defined to be 0 V. The standard oxidation-reduction potentials of the coenzymes are: (NADH+H) (-0.32 V) and (NADPH+H) (-0.32 V), FADH2 (-0.22). If necessary (NADPH+H) reduces NAD to (NADH+H) catalyzed by NADPH dehydrogenase, in this way it can take part in the electron transport. (NADPH+H) + NAD NADP + (NADH+H) Reaction catalyzed by NADPH dehydrogenase 49 There are three of transitions of electron transport involved in the energy producing oxidative phosphorylation and in these transitions there are large differences between the standard oxidation-reduction potential of electrons of the participants. These differences result in proton pumps (3×3 protons) connected with a change of the conformation in the membrane proteins. The proton pumps transfer protons not only from the inner membrane but from the matrix (via the dissociation of water molecules) to the intermembrane space causing a significant difference in the pH between the intermembrane space (acidic region) and the matrix (basic region). This difference in pH starts the oxidative phosphorylation to produce a macroerg bond by an ADP+Pi ATP conversion (three ATP molecules in the case of NADH+H). These transitions are: NADH-Q reductase – coenzyme Q, cytochrome reductase – cytochrome c, cytochrome oxidase – oxygen molecule. As the standard oxidationreduction potential of FADH2 is more positive than that of NADH-Q reductase, it can join later (at coenzyme Q) in the electron transport therefore it produces only two ATP molecules by the electron transport. From (NADH+H) to coenzyme Q hydrogen atoms (protons and electrons) take part together in the electron transport chain. From coenzyme Q to oxygen molecules only electrons take part in the electron transport chain; and the protons take part in the proton pump. The direction of electron transport: - values in E’0 + values in E’0 NADH-Q reductase (-0.30 V), coenzyme Q (+0.04 V), cytochrome reductase (cytochrome b + 0.07 V and cytochrome c1 +0.23 V), cytochrome c (+0.25 V), cytochrome oxidase (cytochrome a +0.29 V and cytochrome aa3 +0.55 V), oxygen (+0.82 V). The terminal oxidation pathway and the participants of the electron transport with their standard oxidation-reduction potentials (E’0) To avoid forming free radicals four electrons and four protons are connected to oxygen molecules almost simultaneously producing two molecules of water. There are special enzymes to eliminate the toxic byproducts i.e. the superoxide anion by superoxide dismutase and hydrogen peroxide by catalase. Formation of the superoxide anion: O2 + e O2 Ө Formation of the peroxide anion: O2 + 2e O22 Ө Formation of hydrogen peroxide: 2 H + O22Ө H2O2 Elimination of superoxide anion by superoxide dismutase: 2 O2Ө O22Ө + O2 Elimination of hydrogen peroxide by catalase: 2 H2O2 2H2O + O2 Elimination of toxic oxygen-containing byproducts 50 Oxidative phosphorylation According the Mitchell’s chemiosmotic hypothesis in the oxidative phosphorylation the difference in the pH between the intermembrane space (acidic region) and the matrix (basic region) activates the ATP-synthesizing enzyme complex ATP synthase, also known as FOF1-ATPase. The production of macroerg bonds by ADP+Pi ATP conversions is catalyzed by the F1 subunit of FOF1-ATPase and at the same time the difference in the pH values between the intermembrane space and the matrix is eliminated by the contribution of the FO subunit. The stoichiometry of the terminal oxidation There are so called uncoupling materials eliminating the connection between electron transport and oxidative phosphorylation. Such compounds (e.g. 2,4nitrophenol) are able to transfer protons back to the matrix without forming macroerg bonds. The FO subunit of FOF1-ATPase is involved in this uncoupling activity. The O letter in FO subunit means that the antibiotic oligomycin is also an uncoupling agent. Uncoupling agents are toxic materials, because they eliminate the energy producing function of terminal oxidation. Oxidative phosphorylation and uncoupling agents Connection between the catabolic processes There are complex systems regulating connections between catabolic and anabolic pathways as well as within the catabolic processes. Now some processes supporting the well-balanced metabolism are presented. Anaplerotic reactions There are special reactions named anaplerotic reactions serving to maintain the level of oxaloacetate not only for the citrate acid cycle in mitochondria, but also for 51 other metabolic pathways (e.g. gluconeogenesis) in the cytosol. The activity of the enzymes in these reactions can be regulated by the feedback of acetyl-CoA. The processes are activated by a low acetyl-CoA concentration. In the cytosol phosphoenolpyruvate (in a reversible reaction) and in the mitochondria pyruvate (in an irreversible reaction) can be carboxylated by carbon dioxide to oxaloacetate. It was mentioned earlier that both membranes of the mitochondria are permeable for pyruvate. For the activation of carbon dioxide direct carboxylation reactions always need the energy of a macroerg phosphoric acid anhydride bond (ATP for pyruvate and GTP for phosphoenolpyruvate), and the coenzyme biotin to transfer the carboxylic group. Carboxylation of the biotin-enzyme complex always needs the presence of magnesium ions. The reaction of carboxybiotin-enzyme complex with pyruvate needs the presence of manganese ions. Since the membranes of mitochondria are not permeable for oxaloacetate it can pass through only in its reduced form (L-malate). There exist cytoplasmic and mitochondrial malate dehydrogenase (MDH) enzymes to help the transfer and regenerate oxaloacetate. These reactions play an important role in glucose biosynthesis from pyruvate (gluconeogenesis). The details of this process are given later. Oxaloacetate synthesis from pyruvate catalyzed by pyruvate carboxylase Oxaloacetate synthesis from phosphoenolpyruvate catalyzed by PEP carboxykinase and its passing through the membranes of mitochondria There are other mechanisms which regulate the level of acetyl-CoA in mitochondria. In the case of a low oxaloacetate level of so-called ketone bodies (e.g. in the liver in illness diabetes mellitus) can be formed from acetyl-CoA molecules. From two molecules of acetyl-CoA acetoacetyl-CoA is synthesized. This compound takes part in an addition reaction with the third acetyl-CoA to produce 3-hydroxy-3methylgutaryl CoA (its earlier name is -hydroxy--methylgutaryl CoA). This compound is the starting material of the biosynthesis of terpenes – (details see later). In the synthesis of ketone bodies acetoacetate is formed by the elimination of acetylCoA from 3-hydroxy-3-methylgutaryl CoA. The ketone bodies are: acetoacetate, 52 acetone (produced by the irreversible decarboxylation of acetoacetate) and D-3hydroxybutyrate that is produced by the reversible reduction of acetoacetate accompanied with a NAD (NADH+H) conversion. The biosynthesis of ketone bodies In the case of a low concentration of acetyl-CoA ketone bodies can be a source of acetyl-CoA. In this way they can be used as energy sources (by means of the citrate acid cycle and terminal oxidation). The macroerg thiolester bond of acetoacetyl-CoA is derived from a succinyl CoAsuccinate conversion. Acetoacetyl-CoA can react with acetyl-CoA according to the last step of -oxidation of fatty acids catalyzed by thiolase producing two molecules of acetyl-CoA. Regeneration of acetyl-CoA from acetoacetate Redox shuttles Redox shuttles can regenerate coenzyme NAD from the reduced (NADH+H) formed in the cytosol (e.g. in glycolysis). These hydrogen atoms can be transported to the mitochondria, to the terminal oxidation for producing energy by redox shuttles. These cytoplasmic (cytosolic) coenzymes reduced in this way are often called extra mitochondrial (NADH+H) molecules. There are two redox shuttles for transferring the extra mitochondrial (NADH+H) molecules: the glycerol 3-phosphate (its earlier name is -glycerolphosphate) and the malate-aspartate redox shuttle. 53 The glycerol 3-phosphate shuttle The starting material of glycerol 3-phosphate shuttle is dihydroxyacetone phosphate (DHAP) that is an intermediate of the glycolysis. This compound is reduced by (NADH+H) in a reaction catalyzed by cytoplasmic glycerol 3-phosphate dehydrogenase to glycerol-3-phosphate that can pass the outer membrane of mitochondria. In the intermembrane space glycerol-3-phosphate is oxidized by mitochondrial glycerol 3-phosphate dehydrogenase to DHAP, but the coenzyme of this reaction is FAD. DHAP returns to the cytosol and can repeat this process. At the outer side of the inner membrane the hydrogen atoms of the reduced FADH2 coenzyme enter the electron transport by coenzyme Q – that means the formation of only two instead of three ATP molecules from ADP. This seems to be a loss in transporting of reducing equivalents, but in reality it is an active membrane transport, since the hydrogen atoms of the reduced FADH2 coenzyme can enter the electron transport even at high mitochondrial concentration of reduced coenzyme (NADH+H). Malate-aspartate redox shuttle In the case of the malate-aspartate redox shuttle oxaloacetate is reduced by extra mitochondrial (NADH+H) to L-malate by cytoplasmic malate dehydrogenase (MDH) for which the membranes of mitochondria are permeable. In the matrix mitochondrial MDH regenerates oxaloacetate forming reduced (NADH+H). It means that there is no loss in transporting of reducing equivalents in this case. Since the inner membrane of mitochondria is not permeable for oxaloacetate only for aspartate and ketoglutarate, therefore oxaloacetate leaves the mitochondria in a roundabout way. 54 Oxaloacetate and glutamate give -ketoglutarate and aspartate in a transamination reaction. They leave the mitochondria by antiport membrane transport processes by the help of translocases. Oxaloacetate is regenerated by the transamination reaction of -ketoglutarate and aspartate producing glutamate for which membranes of mitochondria is permeable by an antiport membrane transport process. The pairs in the antiport membrane processes are aspartate and glutamate, as well as ketoglutarate and oxaloacetate. This reversible redox shuttle can work only in when the (NADH+H)/NAD ratio is higher in the cytosol than in the mitochondrial matrix (e.g. in tissues of high capacity as the human heart and liver). The stoichiometry of aerobic oxidative degradation of glucose During the oxidative degradation of glucose when the glycerol 3-phosphate shuttle is used, the overall reaction is: C6H12O6 + 6 O2 6 CO2 + 6 H2O accompanied by the reaction: 36 ADP + 36 Pi 36 ATP + 36 H2O. In the case of the malate-aspartate shuttle the number of ATP molecules is 38. The pentose phosphate pathway The pentose phosphate pathway (earlier called the direct oxidation of glucose or the phosphogluconate pathway or the hexose monophosphate shunt) is an alternative cytoplasmic oxidative degradation of glucose resulting in (NADPH+H) from NADP as well as different intermediates (e.g. ribose 5-posphate, ribulose 5phosphate, erythrose-4-phosphate). The reduced coenzyme (NADPH+H) is the coenzyme of reductive biosyntheses for all kinds of living organisms. Except plants (using the energy of photons for glucose biosynthesis) and some microorganisms (using other chemical energy) the living organisms use pentose phosphate pathway for the biosynthesis of (NADPH+H). Moreover (NADPH+H) is an antioxidant reducing agent in living organisms (e.g. it can regenerate hemoglobin containing ferrous ion from methemoglobin containing ferric ion by reduction). Ribose 5posphate is one of the starting materials of the biosynthesis of nucleic acids. Ribulose 5-posphate is the starting material of the synthesis of ribulose 1,5-bisposphate being the starting material of Calvin cycle. Erythrose-4-phosphate is used in the synthesis of aromatic amino acids. Reactions catalyzed by transketolases and transaldolases 55 The starting material of the pentose phosphate pathway is glucose 6-phosphate that is oxidized to 6-phosphoglucono--lactone (the coenzyme is NADP reduced to NADPH+H) catalyzed by glucose 6-phosphate dehydrogenase. After the hydrolysis of this lactone to 6-phosphogluconate this molecule is oxidized by NADP (which is reduced to NADPH+H) and ribulose 5-phosphate and carbon dioxide are produced. From ribulose 5-phosphate by isomerisation ribose 5-phosphate and xylulose 5phosphate (by epimerization that is a change in the chirality) are formed. In the next steps of the pathway by means of transferase enzymes transketolases and a transaldolase from six ribulose 5-phosphate molecules five glucose 6-phosphate molecules are formed in a complicated system. Both transaldolase and transketolases can perform transformations of aldoses to ketoses and inversely by transferring C-2 (transketolases) or C-3 (transaldolase) fragments from a ketose to an aldose. The chirality of the new hydroxyl group from the aldehyde group is L in the case of transketolases and D in the case of transaldolase. The pentose phosphate pathway is regulated by the level of NADP. 56 The pentose phosphate pathway The stoichiometry of the pentose phosphate cycle In the pentose phosphate cycle during the degradation of one glucose molecule twelve reduced coenzymes (NADPH+H) (equivalent to 36 ATP molecules) and six carbon dioxide molecules are produced. On the basis of equivalents between reduced coenzyme (NADPH+H) and macroerg phosphoric acid anhydride bond of ATP the pentose phosphate cycle is equivalent to the aerobic oxidative degradation of glucose 57 via the glycerol 3-phosphate shuttle. From six glucose 6-phosphate molecules twelve reduced coenzymes (NADPH+H), six carbon dioxide molecules and six ribose 5phosphate molecules are produced. From six ribose 5-phosphate molecules (C5) five glucose 6-phosphate molecules (C6) were formed by sugar transformation reactions. Stoichiometry of the pentose phosphate cycle Anabolism In the following the reductive biosynthetic pathways of different biomolecules are presented, at first the anabolic reactions of glucose. Biosynthesis of glucose In animals the maintenance of glucose level can be maintained (beyond the direct use of the glucose content of foods) by the biosynthesis glucose from noncarbohydrate precursors called gluconeogenesis. In plants glucose is synthesized by photosynthesis. During the germination of seeds glucose can be synthesized by different ways. Gluconeogenesis Gluconeogenesis (or ‘de novo’ synthesis of glucose) is a glucose biosynthesis from non-carbohydrate precursors (pyruvate, L-lactate, DHAP, glycerol, some amino acids degradation of which gives pyruvate or an intermediate of the citric acid cycle – these are glucogenic amino acids). Animals cannot synthesize glucose from fatty acids. In the liver glucose is synthesized from lactate formed by glycolysis and lactic acid fermentation in skeletal muscles when the rate of glycolysis exceeds the metabolic rate of the citric acid cycle and the terminal oxidation (respiratory chain) (anaerobe period of muscle activity). This regeneration of glucose used up by muscles is called the Cori cycle. Gluconeogenesis helps to maintain the glucose level in blood and brain. The gluconeogenesis is not a simple reversal of glycolysis. It was shown earlier that there are three irreversible steps of glycolysis: The glucose glucose 6phosphate and fructose 6-phosphate fructose 1,6-bisphosphate reactions catalyzed by kinase enzymes. These reactions utilize the energy of a macroerg phosphoric acid anhydride bond of an ATP molecule, but sugar phosphates do not contain macroerg bonds. The reverse reactions are hydrolyses catalyzed by phosphatases (glucose 6phosphatase and fructose 1,6-bisphosphatase) to glucose and fructose 6-phosphate and Pi. The third irreversible step of glycolysis is the phosphoenolpyruvate pyruvate conversion because of the irreversibility of oxo-enol tautomerism. Pyruvate formed in the cytoplasm and entered to the mitochondria is carboxylated to oxaloacetate (by pyruvate carboxylase) as it was described in the section for anaplerotic reactions. The pass of oxaloacetate from mitochondria to the cytosol in form of malate and the conversion of the regenerated oxaloacetate to PEP is the 58 reverse process catalyzed by PEP carboxykinase which were also described earlier. This reaction is reversible, because during the decarboxylation of oxaloacetate first enolpyruvate is formed that is immediately (before the irreversible tautomerism) phosphorylated by GTP. In the scheme of gluconeogenesis the mitochondrial reactions are in a frame. Gluconeogenesis The summary of the process of gluconeogenesis is given in a separate scheme with the names of intermediates (lactate in Hungarian is tejsav). 59 The process of gluconeogenesis Photosynthesis Practically all energy consumed by living organisms arises from solar energy that is trapped by the process of photosynthesis. Only a few microorganisms can use other chemical energy. For the biosynthesis of one glucose molecule the energy of twelve photons is used: 6 H2O + 6 CO2 12 h C6H12O6 + 6 O2 Water and carbon dioxide molecules are incorporated in separate processes. As a result of photolysis the hydrogen atoms of water are transported separately as protons and electrons in the light phase (electron transport) of photosynthesis to NADP molecules to produce reduced (NADPH+H) molecules by the energy of absorbed light. Photosynthesis in green plants takes place in the chloroplasts. The absorption of four photons is accompanied by the synthesis of two macroerg phosphoric acid anhydride bonds of two ATP molecules and the formation of one oxygen molecule. The stoichiometry of the light phase of photosynthesis The assimilation of carbon dioxide molecules to ribulose 1,5-bisphosphate by the contribution of (NADPH+H) and ATP molecules take place in the dark phase of the photosynthesis (Calvin cycle). This phase can be carried out without the presence of light. 60 The stoichiometry of the dark phase of photosynthesis Photosynthesis in green plants takes place in chloroplasts in which there is a so-called thylakoid membrane system the structure of which resembles to that of the of mitochondria. There are two photosystems in chloroplasts that can trap the energy of photons in a complicated system containing chlorophylls (molecules containing a porphyrin skeleton and a coordinated magnesium ion). The stroma that is similar to the matrix of mitochondria contains all of the enzymes of the dark phase of the photosynthesis. Chlorophyll a and b The light phase of the photosynthesis The light phase of photosynthesis can be illustrated by a Z-scheme that shows the standard oxidation-reduction potentials of the participants of the electron transport from water to the coenzyme NADP. As it was mentioned earlier the direction of the electron transport is determined by the standard oxidation-reduction potential (E’0) of the participants (from the negative to the positive values), the Z-scheme illustrates that the standard oxidation-reduction potential of photosystems can be changed to the 61 negative region by the energy of the photons absorbed. The energy of absorbed photons is the driving force for the electron transport from water molecules (E’0 +0.80) to NADP (E’0 -0.32). The Z-scheme of photosynthesis At first photosystem I (PS-I) absorbs light of 700 nm wavelength (P-700). Affected by the photon the standard oxidation-reduction potential of PS-I changes from about +0.5 V to about -1.3 V. One of the electrons of chlorophyll of PS-I from excited PS-700* enters the electron transport cascade and through different complex iron-sulphur proteins (clusters). One of them is the ferredoxin reducing system (FRS) and the other is ferredoxin (Fd), ferredoxin-NADP reductase (Fd-N-Ox). Finally the electron reduces NADP to (NADPH+H). Clusters are proteins containing sulphur both in covalent (in cysteine) and ionic (between inorganic sulphur and iron) bonds. The electron deficiency of PS-I is eliminated by excitation of photosystem II (PS-II) by light of 680 nm wavelength (P-680). The standard oxidation-reduction potential of PS-II is changed from about +1.0 V to about -0.80 V by the second photon. One of electrons of chlorophyll in PS-II enters electron transport between excited P-680* and P-700 and through several complexes: a primary electron acceptor that contains pheophytin (PEA), different plastoquinones QA, QB, QH2 – (this reduced plastoquinone is called plastoquinone pool PQ), then to the cytochrome bf complex, and plastocyanin (PC). Because of the proton pump between cytochrome b and cytochrome f this electron transport is accompanied by yielding one macroerg bond in an ADP+Pi ATP conversion. This reaction is catalyzed by CFOCF1-ATPase. Electron deficiency in PS-II is stopped by exciting the photolysis of water to protons, electrons and oxygen. The electrons enter PS-II by the mediation of a cluster complex containing manganese. This cluster can prevent the formation of oxygen molecules from the generating of dangerous oxygen radicals. Protons of water can reach the coenzyme reduced by electrons with the help of a proton gradient. This last part of the light phase of the photosynthesis is called the Hill reaction. 62 When the reduced coenzyme level is high, there is an alternative pathway for electrons from P-700* through another ferredoxin to the cytochrome bf complex. In this way the energy of one photon leads to a formation one macroerg bond of ATP: h ATP. This process is the cyclic photophosphorylation. During the light phase from two photons one reduced coenzyme (equivalent to the energy of three macroerg bond of ATP) and one macroerg bond of ATP is formed: 2 h 4 ATP that means: h 2 ATP. Cyclic photophosphorylation provides the synthesis of only a half of ATP molecules, but it is an easy way to produce extra ATP molecules. The dark phase of the photosynthesis (Calvin cycle) In the dark phase of photosynthesis the fixation (assimilation) of carbon dioxide to ribulose 1,5-bisphosphate by the contribution of (NADPH+H) and ATP molecules leads to the biosynthesis of glucose and this process is connected with the regeneration of ribulose 5-phosphate: it is phosphorylated by the extra ATP of cyclic photophosphorylation to restore ribulose 1,5-bisphosphate. The Calvin cycle The enzyme for carbon dioxide assimilation to ribulose 1,5-bisphosphate is ribulose 1,5-bisphosphate carboxylase (its short name is Rubisco) which is also an oxygenase. The addition of CO2 is (through an enediol intermediate) between C-2 and C-3 results in two glycerate 3-phosphate molecules but only one of them contains the assimilated carbon dioxide as a carboxylic group. Similarly to the glycolysis only glycerate 1,3-bisphosphate is able to take part in a oxidation-reduction reaction. In this case glycerate 3-phosphate is phosphorylated by ATP (formed in the light phase) catalyzed by a kinase, then glycerate 1,3-bisphosphate containing a macroerg mixed acid anhydride bond is reduced by (NADPH+H) (formed in the light phase) to 63 glyceraldehyde 3-phosphate. Glucose is synthesized by the steps of gluconeogenesis but only in the ratio of the absorbed carbon dioxide. From the other glyceraldehyde 3phosphate molecules ribulose 5-phosphate is regenerated by similar transformations catalyzed by transketolases and a special aldolase presented earlier in the pentose phosphate pathway. Ribulose 1,5-bisphosphate is synthesized by the phosphorylation by ATP (formed in the cyclic photophosphorylation in the light phase). The traditional photosynthesis is called C3 photosynthesis, because the intermediates of the assimilation of carbon dioxide contain three carbon atoms. Rubisco is also an oxygenase; therefore it catalyzes the addition of an oxygen molecule to ribulose 1,5-bisphosphate resulting in glycerate 3-phosphate and phosphoglycolate (H2O3P-O-CH2–COOH). From phosphoglycolate glycine can be synthesized. Generally the rate of carboxylase reaction is four times that of oxygenase reaction. The name of this disadvantageous reaction is photorespiration. There is another variation of photosynthesis in tropical plans because of the extreme circumstances. The light and dark phases are working separately and carbon dioxide is stored temporarily in L-malate produced by the carboxylation of PEP followed by reduction. Because malate contains four carbon atoms, this variation is called C4 photosynthesis. Glucose biosynthesis in different seedlings The seedlings in the ground are unable to photosynthesize therefore they use the stored biomolecules of the seeds for glucose biosynthesis. There are different kinds of seeds. Cereals are rich in polysaccharides (e.g. starch) and glucose is produced by their hydrolysis. From the seeds that are rich in proteins (e.g. bean) after the hydrolysis of proteins glucose can be synthesized from glucogenic amino acids by gluconeogenesis. From oilseeds (e.g. sunflower seeds) that are rich in oil glucose can be synthesized after the oxidative degradation of fatty acids from acetyl-CoA molecules by the glyoxylate cycle. The glyoxylate cycle There are many bacteria and plants that can synthesize glucose from acetylCoA in a biosynthetic process called the glyoxylate cycle. In plants the glyoxylate cycle occurs in organelles called glyoxysomes. Until the synthesis of isocitrate the steps of glyoxylate cycle are the same that of the citric acid cycle that is degradation process (the addition of acetyl-CoA to the starting material oxaloacetate, rearrangement of citric acid to isocitrate). In the glyoxylate cycle isocitrate takes part in an elimination reaction producing succinate and glyoxylate. From succinate oxaloacetate can be synthesized in three steps following the citric acid cycle which can be then serve as starting material for the biosynthesis of glucose by gluconeogenesis. From glyoxylate oxaloacetate (the starting material of glyoxylate cycle) is regenerated by the addition of another molecule of acetyl-CoA followed by the oxidation of the product L-malate. These reactions are catalyzed first by malate synthase that resembles citrate synthase in the citric acid cycle, then by malate dehydrogenase similarly to the last step of the citric acid cycle accompanied by a NAD (NADH+H) conversion. In the glyoxylate cycle succinate (C4) can be derived from two molecules of acetyl-CoA (C2). 64 The glyoxylate cycle The scheme of the biosynthesis of glucose by the glyoxylate cycle The biosynthesis of polysaccharides After the activation of the glycosidic hydroxyl group of sugars O-glycosides can be synthesized in the cytosol. It is illustrated by the example of glucose. Glucose 6-phosphate (the first intermediate of glycolysis) is converted to glucose 1-phosphate by means of a combination of two transfer reactions assisted by the cofactor glucose 1,6-bisphosphate. The reaction is catalyzed by a mutase (glucose-phosphate mutase or phosphoglucose mutase) a transferase. A reverse process is carried out after the degradation of glycogen by phosphorylase. 65 The reaction catalyzed by glucose-phosphate mutase The active NDP-sugar derivatives (that are sugar donors for the biosynthesis of O-glycosides) are produced from sugar 1-phosphate and nucleoside triphosphate (NTP) molecules. NDP- sugars contain a phosphoric acid anhydride macroerg bond and the energy of the other macroerg bond of NTP is used for the biosynthesis of the active sugar derivative. The hydroxyl component can be an alcoholic or glycosidic hydroxyl group of another sugar or an aglycon molecule. The formation of the glycosidic bond does not need extra energy; the byproduct of the reaction is NDP. Biosynthesis of O-glycosides of glucose In the case of glucose the starting material is NDP-glucose. For sucrose biosynthesis UDP-glucose is the starting material. For the biosynthesis of glucans the starting material for amylose is ADP-glucose and for cellulose that is GDP-glucose. 66 Biosynthesis of glucans The biosynthesis of lipids The staring material of both fatty acids and terpenes is acetyl-CoA. The biosynthesis of fatty acids The biosynthesis of fatty acids till the C16 stage is takes place in the cytosol. Acetyl-CoA is transported from mitochondria to cytosol by a combination of reactions presented earlier. The way of acetyl-CoA from the mitochondria to the cytosol The biosynthesis is catalyzed by a multienzyme complex (fatty acid synthase) that contains six active sites and an acyl carrier protein (ACP). ACP binds the acyl group of the fatty acid of increasing number of carbon atoms by a macroerg thiolester. There are two acetyl-CoA molecules at the start of the biosynthesis. One of the acetylCoA molecules is joined to the ACP, to the site of the prospective long fatty acid 67 (catalyzed by acetyl transacylase). The other acetyl-CoA is connected to the site of the new acetyl-CoA units and carboxylated to malonyl-CoA by the coenzyme biotin (as prosthetic group) aided by the energy of ATP ADP conversion (catalyzed by acetyl-CoA carboxylase) followed by a transacylation reaction from coenzyme A to ACP. ATP plays role in the formation of carboxybiotin. The formation of malonyl-CoA is often named the ‘activation of acetyl-CoA’. In fact it is not a real activation step, because the number of macroerg bonds does not increase. But the electron distribution of malonyl-CoA can help its connection to an acyl-ACP accompanied by a decarboxylation catalyzed by -acyl-ACP-synthase (acylmalonyl-ACP condensing enzyme). The driving force of the synthesis of acetoacetylACP is the elimination of carbon dioxide. The next three steps (reduction of the -keto group, dehydration, and reduction of the unsaturated double bond) of fatty acid biosynthesis are similar to the steps of the -oxidative degradation of fatty acids in the opposite direction, but with several differences: the steps of the biosynthesis are in a chain connected to ACP (instead of CoA); the coenzyme of both reductive reactions is (NADPH+H) (instead of NAD and FAD); in hydration the D-epimer (enantiomer) is formed instead of the Lenantiomer, therefore an epimerization is also needed. The end product of the first elongation cycle is butyryl-ACP that reacts with a second malonyl-CoA. Elongation may continue until palmitoyl-ACP that is hydrolyzed to ACP and palmitate from which palmitoyl-CoA is synthesized in two steps. Further C2 units can be attached to the chain by the fatty acid elongation system in the endoplasmic reticulum membrane (in closed vesicles called microsomes) with reactions similar to those of cytoplasmic ACP. Biosynthesis of fatty acids 68 Unsaturated fatty acids are synthesized from the saturated fatty acids in microsomes catalyzed by a monooxygenase. Monooxygenases can use molecular oxygen for the oxidation but only one of oxygen atoms oxidizes the saturated C–C bonds to double bonds, the other oxygen is reduced by (NADPH+H) using an electron transport chain containing NADH-cytochrome b5 reductase (with FAD/FADH2 content), cytochrome b5 reductase (with Fe2/Fe3 content) and desaturase (with Fe2/Fe3 content). This process cannot be carried out in mammals therefore linoleate and linolenate are essential fatty acids. In mammals these fatty acids are the starting materials for other unsaturated fatty acids of biological importance (e.g. arachidonate for the biosynthesis of prostaglandin hormones) Formation of unsaturated fatty acids The biosynthesis of triacylglycerols and phosphoglycerides Triacylglycerols (triglycerides) and phosphoglycerides are synthesized in the endoplasmic reticulum membrane and their common intermediates are phosphatidates (phosphatidic acids) containing different fatty acid components. The starting material of phosphatidates is glycerol 3-phosphate that is synthesized by the reduction of DHAP. This reaction in the opposite way described in the section of redox shuttles. In the biosynthetic process the phosphoric acid unit of glycerol 3-phosphate can help the formation of a link between the substrate and enzyme by ionic interactions. The hydroxyl groups of glycerol 3-phosphate can react with different acylCoA molecules to produce phosphatidates. In the case of the biosynthesis of triacylglycerols phosphatidates are hydrolyzed by special phosphatases to produce diacylglycerols that react with a third acyl-CoA. The enzymes of this process are associated in a triacylglycerol synthetase complex. The biosynthesis of triglycerides 69 In the case of the biosynthesis of phosphoglycerides there are different possibilities to introduce another ester group to phosphatidates. One of these possibilities is the activation phosphatidates by forming a macroerg phosphoric acid anhydride bond by the reaction with CTP. This reaction is similar to the activation of glucose 1-phosphate to NDP-glucose that was described in the section of the biosynthesis of polysaccharides. CDP-diacylglycerols can produce phosphatidyl ethanolamines with ethanolamine, phosphatidyl serines with serine, and phosphatidyl cholines with choline (lecitines). The byproduct of the reactions is CDP. Another variation is that the hydroxyl compound is activated by phosphorylation followed by a reaction with CTP. In this case the CDP derivative of the hydroxyl compounds is reacted with diacylglycerols (e.g. in the synthesis of lecitines diacylglycerols are reacted with CDP-choline). Variations for the biosynthesis of phosphatidyl cholines (lecitines) The biosynthesis of lecitine The biosynthesis of terpenes The starting material of the cytoplasmic biosynthesis of terpenes is acetylCoA, and the first phase is the synthesis of isopentenyl pyrophosphate from three acetyl-CoA molecules. As it was mentioned in connection with the synthesis of ketone bodies acetoacetyl-CoA is synthesized from two molecules of acetyl-CoA. This compound takes part in an addition reaction with a third acetyl-CoA to produce 3hydroxy-3-methylgutaryl CoA (its earlier name is -hydroxy--methylgutaryl CoA). In an addition reaction a nucleophilic attack of acetyl group on the carbonyl group of acetoacetyl-CoA is carried out by the methylene part of the enol tautomer that is temporarily formed in the reaction under the influence of the strong hydrogen bond of a His of the enzyme. Acetyl-CoA reacted with oxaloacetate similarly in the first step of the citric acid cycle. In the mitochondrial synthesis of ketone bodies 3-hydroxy-3-methylgutaryl CoA takes part in an elimination reaction. In the cytoplasmic synthesis of terpenes 3hydroxy-3-methylgutaryl CoA is reduced by a reductase to mevalonate accompanied 70 by 2 (NADPH+H) 2 NADP conversion in an irreversible reaction. After phosphorylation to 5-pyrophosphomevalonate by 2 ATP molecules accompanied by a decarboxylation isopentenyl pyrophosphate is formed. Isopentenyl pyrophosphate and its isomer dimethylallyl pyrophosphate condense to form the monoterpene geranyl pyrophosphate (C10). The intermediate of this reaction is an allylic carbonium ion (named active isoprene) formed from dimethylallyl pyrophosphate. Geranyl pyrophosphate can then be transformed to other terpenes. Biosynthesis of the monoterpene geranyl pyrophosphate 71 Biosynthesis of amino acids The biosynthesis of proteins from L--amino acids is connected with the metabolism of nucleic acids. There are different pathways for the biosynthesis of amino acids but some common features can be found. The starting materials of the carbon skeletons of amino acids are the intermediates of glycolysis, pentose phosphate pathway and citric acid cycle. On the basis of the starting materials five biosynthetic families of amino acids can be distinguished. The name of these families is based on the compound (generally amino acid) which can be the starting material of the other members of the family. The starting material of the glutamate amino acid family (Glu, Gln, Pro, Arg) is -ketoglutarate (from the citric acid cycle). The starting material of the aspartate amino acid family (Asp, Asn, Met, Lys, Thr, Ile) is oxaloacetate (from the citric acid cycle). The starting material of the serine amino acid family (Ser, Cys, Gly) is 3phosphoglycerate (from the glycolysis). The starting material of the pyruvate amino acid family (Ala, Val, Leu) is pyruvate (from the glycolysis). The starting materials of the aromatic amino acid family (Phe, Tyr, Trp) are phosphoenolpyruvate (from the glycolysis) and erythrose 4-phosphate (from the pentose phosphate pathway). The starting material of histidine is ribose 5-phosphate (from the pentose phosphate pathway). Generally the last step of the biosynthesis of amino acids is the transamination reaction of an -keto carboxylic acids but there are several exceptions. The metabolism of the biomolecules of genetic information As it was mentioned earlier the metabolism of nucleic acids is in separated place from the metabolism of other biomolecules in the nucleus of the cell. The metabolism of nucleic acids is the subject of another science (genetics) therefore now only a short summary is given. Hydrolysis of nucleic acids Nucleases (DNase an RNase enzymes) hydrolyze nucleic acids (DNA and RNA) to nucleotides. There are exonucleases and endonucleases. Exonucleases can be specific for the 3' or 5' end of the strand. From DNase enzymes restriction endonucleases (that cleave DNA chain at specific sites) are often used in genetic engineering. During further hydrolytic processes nucleosides then nucleic bases are formed. Oxidative degradation of pyrimidine nucleic bases During the oxidative degradation of pyrimidine nucleic bases (through the intermediates uracil and then dihydrouracil) ammonia and succinate are formed. 72 Oxidative degradation of pyrimidine nucleic bases Oxidative degradation of purine nucleic bases As first intermediates xanthines (hypoxanthine and xanthine) then uric acid (in Hungarian hugysav) are formed by oxidative deamination of purine nucleic bases followed by an oxidative degradation to urea and glyoxylate. Oxidative degradation of purine nucleic bases Biosynthesis of pyrimidine nucleotides The pyrimidine ring is synthesized from carbamoyl phosphate (formed in the urea cycle) and aspartate through the intermediate orotate (orotic acid) that reacts with PRPP (5-phosphoribosyl 1-pyrophosphate) to form the starting material of pyrimidine nucleotides (orotate monophosphate). The amino group of CTP is synthesized from UTP and glutamine accompanied by an ATP ADP + Pi conversion. Thymine is synthesized by the methylation of uracil (coenzyme is THF) in their nucleoside monophosphate form. 73 The scheme of the biosynthesis of pyrimidine nucleotides Biosynthesis of purine nucleotides The starting material of purine nucleotides is PRPP synthesized from ribose 5phosphate (formed in the pentose phosphate pathway). The first step of this complicated process is the synthesis of 5’-phosphoribosylamine (a -glycoside). It is the amino group of 5’-phosphoribosylamine which is incorporated into the purine skeleton. The sources of the different atoms of the purine skeleton are shown below. The scheme of the biosynthesis of purine nucleotides Biosynthesis of the precursors of DNA The reduction of ribose derivatives to 2’-deoxyribose derivatives proceeds in their nucleoside diphosphate form by (NADPH+H). 74 The formation of 2’-deoxyribose from ribose The biosynthesis of DNA (replication) The genetic information is stored and expressed by DNA. During the semiconservative biosynthesis of DNA a new polynucleotide chain in a 5’ to 3’ direction is synthesized from precursor dNTP molecules (dATP, dGTP, dTTP, dCTP) with the hydrolysis of two macroerg phosphoric acid anhydride bonds to the antiparallel complementary original polynucleotide chain (template strand) in the course of dividing of the cell. Two identical DNA molecules are produced from a single double-stranded helical DNA molecule. DNA replication begins at specific locations in the genome, called origins. Building a new nucleotide unit in replication The process of replication is more or less different in prokaryotes and eukaryotes but common features can be found. The break-down of the superhelical structure of DNA is carried out by the enzyme DNA gyrase (topoisomerase I) and unwinding of the double helix is catalyzed by helicase. Opening of the helical structure demands the energy of phosphoric acid anhydride macroerg bonds (ATP ADP + Pi). The construct formed by two separated strands is named the replication fork and fixed by single strand binding (Ssb) proteins. At first a short RNA primer is created on the template strand catalyzed by an RNA polymerase (primase). Elongation of the DNA chain is continued by DNA polymerase III holoenzyme. At the end of elongation instead of DNA polymerase III another enzyme (DNA polymerase I) continues the biosynthesis of DNA strand by removing the RNA primer and replacing it by a DNA section. This is one of the 3’ (5’->3’) exonuclease activities of DNA polymerase I. At the end of the process the end of two DNA chains are connected by DNA ligase using the energy of phosphoric acid anhydride macroerg bond (formed from NAD in prokaryotes and ATP in eukaryotes). 75 The scheme of the replication As the direction of the biosynthesis is 5’3’, it can be carried out directly only along the 3’5’ template strand (called leading strand). On the other strand (called lagging strand 5’3’) the direction of biosynthesis is the opposite to the direction of the sequence, therefore the synthesis of RNA primers are started at several places to the opposite direction (5’3’) followed by elongation of DNA. When the short DNA chains (called Okazaki fragments) reach the next RNA primers the elimination of primers, they replace them by DNA sections and the connection of DNA sections is repeated several times. In the double helix there are double replication forks forming a bubble. The reaction catalyzed by DNA-ligase During the replication all of the new nucleotide units is controlled (and corrected if it is necessary) by DNA polymerase I. This is the other 3’ (5’->3’) exonuclease activity of DNA polymerase I. The 5’ terminal nucleotide unit is also controlled (and corrected if it is necessary). This is the 5’ (3’->5’) exonuclease activity of DNA polymerase I. There are also other possibilities for the correction of the DNA strands after replication. The biosynthesis of RNA (transcription) 76 The biosynthesis of RNA with the participation of DNA is called transcription and consists of the copying of DNA into messenger RNA during gene expression. The technique of transcription is similar to the replication but the new strand is RNA. The precursor molecules are ATP, GTP, UTP and CTP. As only one of the strands of DNA is used as template (leading strand 3’5’) the direction of the biosynthesis and the sequence of the strand is the same. The sequence of new RNA is antiparallel and complementary to the original DNA. When the gene transcribed encodes for a protein, the result of transcription is an mRNA, which is used for the biosynthesis of a protein in the process of translation. Alternatively, the transcribed gene can encode other RNA types (rRNA, tRNA, snRNA). In transcription only a part of the DNA takes part therefore there are several potential starting sites on it. In eukaryotes transcription is catalyzed by RNA polymerase. Initiation of transcription requires the presence of a core promoter sequence on the DNA that is recognized by the subunit of the enzyme. The transcription is continued to the terminal signal sequence on the DNA, that is a palindrome sequence followed by a polyA sequence, therefore the new RNA contains a palindrome folded structure and a tail of polyU at 3’ terminal. Specific proteins called transcription factors play important roles in the transcription. In most often the products of transcription are RNA precursors (pre-RNA molecules) that are modified later. The only exceptions are the prokaryotic mRNA molecules on which the translation (the protein biosynthesis) can be started before the termination of transcription. There are different changes in the pre-mRNA structure of eukaryotes. A polyA tail is attached to the 3’ terminal catalyzed by polyadenylate polymerase using ATP molecules. Another reaction is when a cap (7-methylguanosine pyrophosphate) is connected to the 5’ terminal of pre-mRNA to prevent the molecule from the hydrolysis by RNases. Then a splicing is carried out that is the removal of not to be translated sections called introns from the pre-mRNA strand and connecting the remaining fragments called exons by the help of snRNA molecules). The terminals of both pre-tRNA and pre-rRNA molecules in eukaryotes are hydrolyzed to tRNA molecules. The pre-rRNA molecules can also hydrolyze themselves (this is ribozyme function that is some kind of enzyme function). The biosynthesis of proteins (translation) The sites of protein biosynthesis are ribosomses that contain both rRNA molecules and proteins. The information of the amino acid sequence contains the mRNA that forms a complex with ribosome. The information is in the sequence of nucleotide triplets (called codons). A codon is the sequence of three nucleotide units starting from a fixed point. The genetic codon is universal (it can be used in all kinds of living organisms) and there are no commas and no overlapping in it. The genetic codon is degenerate what means that for most of amino acids have not only one codon. The codon AUG is both the start codon and the codon of methionine (Met). Stop codons are UUA, UGA and UAG. Amino acids are transferred to the ribosome in ester form that can react with an amino group (aminoacyl tRNA). At first an amino acid is connected to AMP by a phosphoric acid anhydride macroerg bond of ATP and catalyzed by aminoacyl tRNA synthetase. Then aminoacyl AMP is reacted with the 3’ hydroxyl group (with CCA terminus) of tRNA to produce aminoacyl tRNA. The DHU loop of tRNA is connected to the enzyme aminoacyl tRNA synthetase. The anticodon section of tRNA contains an antiparallel and complementary sequence of the genetic codon of the amino acid to 77 be transferred; therefore it is the template recognizing site of tRNA. The third nucleotide member of the anticodon can be different nevertheless tRNA molecules can recognize such a codon. The phenomenon is called wobble in base pairing. Formation of aminoacyl tRNA The structure of tRNA There are two sites in the initiation complex of ribosome and mRNA: site P (for the growing peptide chain connected to tRNA) and site A (for the new amino acid unit in aminoacyl tRNA form). At the start of the protein biosynthesis (initiation step) a Met-tRNA (in prokaryotes formylMet-tRNA) is connected to site P and the next aminoacyl tRNA to the A site (on the basis of the next codon). The connection of the aminoacyl tRNA to the site needs the energy of a phosphoric acid anhydride macroerg bond (hydrolysis of GTP to GDP and an inorganic phosphate) catalyzed by a GTPase. 78 Formation of peptide bond (O-N acyl transfer reaction) The next step is elongation of the chain that is the formation of a carboxamide (peptide) bond between the ester group of aminoacyl tRNA connected to site P and the amino group of aminoacyl tRNA connected to the A site. This step is often called O-N acyl transfer reaction. The free tRNA leaves site P and tRNA containing dipeptide chain is moved from site A to P site by protein elongation factors using the energy of a phosphoric acid anhydride macroerg bond of the hydrolysis of GTP (to GDP and an inorganic phosphate) catalyzed by a GTPase. At the same time the position of mRNA is changed in the complex, in this way the next codon is moved to site A and the elongation can be continued. Each new amino acid unit in the peptide chain is controlled (and corrected if it is necessary) by a special mechanism during translation. When a stop codon is found at site A elongation stops and the new peptide chain is hydrolyzed from the tRNA by the help by release factors. Proteins biosynthesis is regulated in different ways in prokaryotes and eukaryotes. Parallel with translation the formation of secondary, tertiary and quaternary structures of proteins proceeds. There can be different post-translational modifications on the protein structure e.g. cleavage the N-terminal methionine, different changes in the side chains of amino acid units (e.g. the oxidation of proline to hydroxyproline described earlier), etc. Literature 1. Stryer, L.: Biochemistry (3rd Edition) W.H. Freeman & Company New York 1988. 2. Ádám, G. and Fehér, O. (Eds): Élettan biológusoknak (Physiology for biologists). Tankönyvkiadó, Budapest, 1990, pp 20. Topics in Biochemistry 1. Definition of biomolecules. Proteins – structures, biochemical functions and figures of some representatives. 2. Definition of biomolecules. Carbohydrates – structures, biochemical functions and figures of some representatives. 3. Definition of biomolecules. Simple lipids – structures, biochemical functions and figures of some representatives. 4. Definition of biomolecules. Complex lipids – structures, biochemical functions and figures of some representatives. 5. Definition of biomolecules. Nucleic acids – structures, biochemical functions and figures of some representatives. 79 6. Enzymes – structure and function (equations) 7. Degradation of biomolecules. The first step – hydrolysis. 8. Degradation of biomolecules. Glucose to the general intermediate (scheme). 9. Degradation of biomolecules. Fatty acids to the general intermediate (scheme). 10. Degradation of biomolecules. Amino acids to the general intermediate (scheme). 11. Degradation of biomolecules. Formation of CO2 during the degradation of the general intermediate (scheme). 12. Degradation of biomolecules. Formation of H2O during the degradation of the general intermediate (scheme). 13. Degradation of biomolecules. The alternative degradation of glucose – biological functions in autotrophic and heterotrophic living organisms (scheme of some starting steps and the stoichiometry of the reaction). 14. The principle of redox shuttles and gluconeogenesis (discussion on the scheme of glycolysis). 15. Photosynthesis. The role of photons (Z-scheme). 16. Photosynthesis. Assimilation of CO2 (scheme of some starting steps and the stoichiometry of the reaction). 17. Biosynthesis of glucose in seeds (wheat, bean, sunflower). The glyoxylate cycle. 18. Biosynthesis of the glycosidic bond. 19. Biosynthesis of fatty acids and triglycerides. 20. Biosynthesis of DNA and RNA 21. Biosynthesis of proteins.