Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
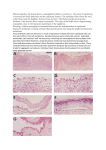
CHUKWUEMEKA JULIET- 17CP022984 TYPES OF CELLS IN THE AVEOLI SPACE 1. TYPE 1 CELLS Type I cells are the larger of the two cell types; they are thin, flat epithelial lining cells (membranous pneumocytes), that form the structure of the alveoli. They are squamous (giving more surface area to each cell) and have long cytoplasmic extensions that cover more than 95% of the alveolar surface. Type I cells are involved in the process of gas exchange between the alveoli and blood. These cells are extremely thin – sometimes only 25 nm – the electron microscope was needed to prove that all alveoli are lined with epithelium. This thin lining enables a fast diffusion of gas exchange between the air in the alveoli and the blood in the surrounding capillaries. The nucleus of a type I cell occupies a large area of free cytoplasm and its organelles are clustered around it reducing the thickness of the cell. This also keeps the thickness of the bloodair barrier reduced to a minimum. The cytoplasm in the thin portion contains pinocytotic vesicles which may play a role in the removal of small particulate contaminants from the outer surface. In addition to desmosomes, all type I alveolar cells have occluding junctions that prevent the leakage of tissue fluid into the alveolar air space. The relatively low solubility (and hence rate of diffusion) of oxygen, necessitates the large internal surface area (about 80 square m [96 square yards]) and very thin walls of the alveoli. Weaving between the capillaries and helping to support them is an extracellular matrix, a meshlike fabric of elastic and collagenous fibres. The collagen fibres, being more rigid, give the wall firmness, while the elastic fibres permit expansion and contraction of the walls during breathing. Type I pneumocytes are unable to replicate and are susceptible to toxic insults. In the event of damage, type II cells can proliferate and differentiate into type I cells to compensate. 2. TYPE 2 CELLS Type II cells are cuboidal and much smaller than type I cells. They are the most numerous cells in the alveoli, yet do not cover as much surface area as the squamous type I cells. Type II cells (granulous pneumocytes) in the alveolar wall contain secretory organelles known as lamellar bodies or lamellar granules, that fuse with the cell membranes and secrete pulmonary surfactant. This surfactant is a film of fatty substances, a group of phospholipids that reduce alveolar surface tension. The phospholipids are stored in the lamellar bodies. Without this coating, the alveoli would collapse. The surfactant is continuously released by exocytosis. Reinflation of the alveoli following exhalation is made easier by the surfactant, which reduces surface tension in the thin fluid lining of the alveoli. The fluid coating is produced by the body in order to facilitate the transfer of gases between blood and alveolar air, and the type II cells are typically found at the blood–air barrier. Type II cells start to develop at about 26 weeks of gestation, secreting small amounts of surfactant. However, adequate amounts of surfactant are not secreted until about 35 weeks of gestation – this is the main reason for increased rates of infant respiratory distress syndrome, which drastically reduces at ages above 35 weeks gestation. Type II cells are also capable of cellular division, giving rise to more type I and II alveolar cells when the lung tissue is damaged. MUC1, a human gene associated with type II pneumocytes, has been identified as a marker in lung cancer. 3. BASEMENT MEMBRANES The basement membrane is a thin, pliable sheet-like type of extracellular matrix, that provides cell and tissue support and acts as a platform for complex signalling. The basement membrane sits between epithelial tissues including mesothelium and endothelium, and the underlying connective tissue. The basement membrane is organized as follows: i. Epithelial/mesothelial/endothelial tissue (outer) ii. Basement membrane iii. Basal lamina iv. Lamina lucida v. laminin vi. integrins (hemidesmosomes) vii. nidogens viii. dystroglycans ix. Lamina densa x. collagen IV (coated with perlecan, rich in heparan sulfate) xi. Attaching proteins (between the basal and reticular laminae) xii. collagen VII (anchoring fibrils) xiii. fibrillin (microfibrils) xiv. Lamina reticularis xv. collagen III (as reticular fibers) xvi. Connective tissue (Lamina propria) The primary function of the basement membrane is to anchor down the epithelium to its loose connective tissue (the dermis or lamina propria) underneath. This is achieved by cell-matrix adhesions through substrate adhesion molecules (SAMs). The basement membrane acts as a mechanical barrier, preventing malignant cells from invading the deeper tissues. Early stages of malignancy that are thus limited to the epithelial layer by the basement membrane are called carcinoma in situ. The basement membrane is also essential for angiogenesis (development of new blood vessels). Basement membrane proteins have been found to accelerate differentiation of endothelial cells. The most notable examples of basement membranes is the glomerular basement membrane of the kidney, by the fusion of the basal lamina from the endothelium of glomerular capillaries and the podocyte basal lamina, and between lung alveoli and pulmonary capillaries, by the fusion of the basal lamina of the lung alveoli and of the basal lamina of the lung capillaries, which is where oxygen and CO2 diffusion occurs (gas exchange). As of 2017, other roles for basement membrane include blood filtration and muscle homeostasis. Fractones may be a type of basement membrane, serving as a niche for stem cells. 4. CAPILLARY ENDOTHELIAL CELLS The capillary endothelium is 0.1 μm thick and composed of an area of 126 m2. The capillaries are approximately 7 μm in diameter and are short, resulting in a large surface area. Red blood cells traverse the capillary beds and 2–3 alveoli in approximately 0.75 s. The pulmonary bed receives all of the CO at a low pressure of 15 mmHg. Endothelial cells (ECs) constitute small capillary blood vessels and contribute to delivery of nutrients, oxygen and cellular components to the local tissues, as well as to removal of carbon dioxide and waste products from the tissues. Besides these fundamental functions, accumulating evidence indicates that capillary ECs form the vascular niche. In the vascular niche, ECs reciprocally crosstalk with resident cells such as epithelial cells, mesenchymal cells, and immune cells to regulate development, homeostasis, and regeneration in various organs. Capillary ECs supply paracrine factors, called angiocrine factors, to the adjacent cells in the niche and orchestrate these processes. Although the vascular niche is anatomically and functionally well-characterized in several organs such as bone marrow and neurons, the effects of endothelial signals on other resident cells and anatomy of the vascular niche in the lung have not been well-explored.