Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
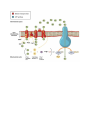
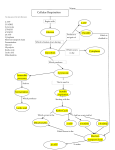
Activity # 5 Metabolism of carbohydrates , fats ,proteins with oxygen availability: Carbohydrates: Carbohydrates are organic molecules composed of carbon, hydrogen, and oxygen atoms. The family of carbohydrates includes both simple and complex sugars. Glucose and fructose are examples of simple sugars, and starch, glycogen, and cellulose are all examples of complex sugars. The complex sugars are also called polysaccharides and are made of multiple monosaccharide molecules. Polysaccharides serve as energy storage (e.g., starch and glycogen) and as structural components (e.g., chitin in insects and cellulose in plants). During digestion, carbohydrates are broken down into simple, soluble sugars that can be transported across the intestinal wall into the circulatory system to be transported throughout the body. Carbohydrate digestion begins in the mouth with the action of salivary amylase on starches and ends with monosaccharides being absorbed across the epithelium of the small intestine. Once the absorbed monosaccharides are transported to the tissues, the process of cellular respiration begins (Figure 1). This section will focus first on glycolysis, a process where the monosaccharide glucose is oxidized, releasing the energy stored in its bonds to produce ATP. Glycolysis: Glycolysis is the process of breaking down a glucose molecule into two pyruvate molecules, while storing energy released during this process as ATP and NADH. Nearly all organisms that break down glucose utilize glycolysis. Glucose regulation and product use are the primary categories in which these pathways differ between organisms. In some tissues and organisms, glycolysis is the sole method of energy production. This pathway is common to both anaerobic and aerobic respiration. Glycolysis consists of ten steps, split into two phases. During the first phase, it requires the breakdown of two ATP molecules. During the second phase, chemical energy from the intermediates is transferred into ATP and NADH. The breakdown of one molecule of glucose results in two molecules of pyruvate, which can be further oxidized to access more energy in later processes. Glycolysis can be regulated at different steps of the process through feedback regulation. The step that is regulated the most is the third step. This regulation is to ensure that the body is not over-producing pyruvate molecules. The regulation also allows for the storage of glucose molecules into fatty acids. There are various enzymes that are used throughout glycolysis. The enzymes upregulate, downregulate, and feedback regulate the process. Krebs Cycle/Citric Acid Cycle/Tricarboxylic Acid Cycle The pyruvate molecules generated during glycolysis are transported across the mitochondrial membrane into the inner mitochondrial matrix, where they are metabolized by enzymes in a pathway called the Krebs cycle (Figure 4). The Krebs cycle is also commonly called the citric acid cycle or the tricarboxylic acid (TCA) cycle. During the Krebs cycle, high-energy molecules, including ATP, NADH, and FADH2, are created. NADH and FADH2 then pass electrons through the electron transport chain in the mitochondria to generate more ATP molecules. To start the Krebs cycle, citrate synthase combines acetyl CoA and oxaloacetate to form a six-carbon citrate molecule; CoA is subsequently released and can combine with another pyruvate molecule to begin the cycle again. The aconitase enzyme converts citrate into isocitrate. In two successive steps of oxidative decarboxylation, two molecules of CO2 and two NADH molecules are produced when isocitrate dehydrogenase converts isocitrate into the five-carbon α-ketoglutarate, which is then catalyzed and converted into the four-carbon succinyl CoA by α-ketoglutarate dehydrogenase. The enzyme succinyl CoA dehydrogenase then converts succinyl CoA into succinate and forms the high-energy molecule GTP, which transfers its energy to ADP to produce ATP. Succinate dehydrogenase then converts succinate into fumarate, forming a molecule of FADH2. Fumarase then converts fumarate into malate, which malate dehydrogenase then converts back into oxaloacetate while reducing NAD+ to NADH. Oxaloacetate is then ready to combine with the next acetyl CoA to start the Krebs cycle again (see Figure 4). For each turn of the cycle, three NADH, one ATP (through GTP), and one FADH2 are created. Each carbon of pyruvate is converted into CO2, which is released as a byproduct of oxidative (aerobic) respiration. Oxidative Phosphorylation and the Electron Transport Chain The electron transport chain (ETC) uses the NADH and FADH2 produced by the Krebs cycle to generate ATP. Electrons from NADH and FADH2 are transferred through protein complexes embedded in the inner mitochondrial membrane by a series of enzymatic reactions. The electron transport chain consists of a series of four enzyme complexes (Complex I – Complex IV) and two coenzymes (ubiquinone and Cytochrome c), which act as electron carriers and proton pumps used to transfer H+ ions into the space between the inner and outer mitochondrial membranes (Figure 5). The ETC couples the transfer of electrons between a donor (like NADH) and an electron acceptor (like O2) with the transfer of protons (H+ ions) across the inner mitochondrial membrane, enabling the process of oxidative phosphorylation. In the presence of oxygen, energy is passed, stepwise, through the electron carriers to collect gradually the energy needed to attach a phosphate to ADP and produce ATP. The role of molecular oxygen, O2, is as the terminal electron acceptor for the ETC. This means that once the electrons have passed through the entire ETC, they must be passed to another, separate molecule. These electrons, O2, and H+ ions from the matrix combine to form new water molecules. This is the basis for your need to breathe in oxygen. Without oxygen, electron flow through the ETC ceases. The electrons released from NADH and FADH2 are passed along the chain by each of the carriers, which are reduced when they receive the electron and oxidized when passing it on to the next carrier. Each of these reactions releases a small amount of energy, which is used to pump H+ ions across the inner membrane. The accumulation of these protons in the space between the membranes creates a proton gradient with respect to the mitochondrial matrix. Also embedded in the inner mitochondrial membrane is an amazing protein pore complex called ATP synthase. Effectively, it is a turbine that is powered by the flow of H+ ions across the inner membrane down a gradient and into the mitochondrial matrix. As the H+ ions traverse the complex, the shaft of the complex rotates. This rotation enables other portions of ATP synthase to encourage ADP and Pi to create ATP. In accounting for the total number of ATP produced per glucose molecule through aerobic respiration, it is important to remember the following points: A net of two ATP are produced through glycolysis (four produced and two consumed during the energy-consuming stage). However, these two ATP are used for transporting the NADH produced during glycolysis from the cytoplasm into the mitochondria. Therefore, the net production of ATP during glycolysis is zero. In all phases after glycolysis, the number of ATP, NADH, and FADH2 produced must be multiplied by two to reflect how each glucose molecule produces two pyruvate molecules. In the ETC, about three ATP are produced for every oxidized NADH. However, only about two ATP are produced for every oxidized FADH2. The electrons from FADH2 produce less ATP, because they start at a lower point in the ETC (Complex II) compared to the electrons from NADH (Complex I) The Fatty Acid Oxidation Pathway Intersects the TCA Cycle In 1904, Knoop, in a classic experiment, decisively showed that fatty acid oxidation was a process by which two-carbon units were progressively removed from the carboxyl end fatty acid molecule. The process consists of four reactions and generates acetyl-CoA and the acyl-CoA molecule shortened by two carbons, with the concomitant reduction of FAD by enzyme acyl-CoA dehydrogenase and of NAD+ by β-hydroxyacyl-CoA dehydrogenase. This pathway is known as βoxidation because the β-carbon atom is oxidized prior to when the bond between carbons β and α is cleaved The four steps of β-oxidation are continuously repeated until the acyl-CoA is entirely oxidized to acetyl-CoA, which then enters the TCA cycle. In the 1950s, a series of experiments verified that the carbon atoms of fatty acids were the same ones that appeared in the acids of TCA cycle. Amino Acid Transamination/Deamination Contributes to the TCA Cycle Two points must be considered regarding the use of amino acids as fuels in energy metabolism. The first is the presence of nitrogen in amino acid composition, which must be removed before amino acids become metabolically useful. The other is that there are at least twenty different amino acids, each of which requires a different degradation pathway. For our purpose here, it is important to mention two kinds of reactions involving amino acid: transamination and deamination. In the first kind of reaction, the enzymes aminotransferases convert amino acids to their respective αketoacids by transferring the amino group of one amino acid to an α-ketoacid. This reaction allows the amino acids to be interconverted. The second kind of reaction, deamination, removes the amino group of the amino acid in the form of ammonia. In the liver, the oxidative deamination of glutamate results in α-keto-glutarate (a TCA cycle intermediate) and ammonia, which is converted into urea and excreted. Deamination reactions in other organs form ammonia that is generally incorporated into glutamate to generate glutamine, which is the main transporter of amino groups in blood. Hence, all amino acids through transamination/deamination reactions can be converted into intermediates of TCA cycle, directly or via conversion to pyruvate or acetyl-CoA (b) Exersices that burn fat: (1) Shadow Boxing (To Burn Fat) it’s one of the cardio exercises you can do at home or maybe anywhere when you want. It doesn’t require any equipment and it can help you burn a ton of calories! you can burn up to 400 calories within 30-40 minutes of shadowboxing. Obviously, if you have a budget to buy a good punching bag and/or an opponent against whom you can box, then even better. This way you can increase the number of calories burned by about 30-50%. But the best part about it is Shadow-boxing helps you work on your abs! Boxing, in general, is a very compound exercise because it essentially activates almost every muscle in your body. Certain movements such as the uppercut and hook really work on the upper-body rotation have an extreme impact on your obliques and upper abs. You can increase your calorie expenditure by adding movements that are of higher intensity, such as knee kick, elbow hit (which also works your abs), spinning-backfist, and more. (2) Rowing (The Fat Burning Exercise) Rowing is one of the fat killer cardio exercises that speed up your heart rate, entangling almost every muscle in the body. The main downside is that you need a Rowing Machine or an actual boat and river where you can, you know, row. Reasons to love rowing machine exercise You can burn up to 400 calories in an interval of 30 minutes. It’s a multi-joint compound exercise that activates almost every muscle in the body. It gives a very good pump for the feet, back and biceps. Improves core strength. Boosts muscle growth. It improves muscle coordination. It is perfect for muscle hydration (pump). What is also really comfortable is that the machine allows you to adjust the force and thus alter the intensity of the rowing. This is perfect because it allows you to do very explosive HIIT in the gym. As I have already mentioned, exercise kills three large muscle groups – the back, legs, and abdomen. How is that of any importance? Essentially, large foundation muscles such as the legs, abs, chest, and back require far more calories to move and recover than other smaller muscle groups like the biceps and triceps. (3) Burpee (Best Cardio To Burn Fat) One of the simplest exercises that can be done anywhere at any time. That leaves your heart thumping against your chest, your lungs on fire, and every muscle completely exhausted. Burpees are estimated to burn approximately 355 calories within a 30-minute timeframe. The Burpee is definitely one of those cardio exercises that don’t just help you burn fat but it will also help you build some muscle definition. From a standing position, you go down into a squat. You put your hands to the ground and keep them extended and you kick your feet behind you – you are now in a plank position. Instead of stopping there drop yourself to the ground and do a push-up. From there as you push yourself back and you return back to your starting position as you jump in the air you grab on a pull-up bar and you do a pull-up. This is going to leave your body completely exhausted after a couple of repetitions. There are also a couple of other really good variations of the classic Burpee that can help you increase the intensity and burn some extra calories.