Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
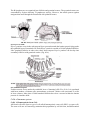
C-9: IMMUNOLOGY Syllabus UNIT-I Immune Response - An overview, components of mammalian immune system, molecular structure of Immuno-globulins or Antibodies, Humoral & Cellular immune responses, Tlymphocytes & immune response (cytotoxic T-cell, helper T-cell, suppressor T-cells), T-cell receptors, genome rearrangements during B-lymphocyte differentiation, Antibody affinity maturation class switching, assembly of T-cell receptor genes by somatic recombination. NOTES (Part-I) Immune Response - An overview The B and T-lymphocytes that are yet to encounter an antigen are called naive B and naive Tcells. The naive B-cells that encounter the antigen, proliferate and differentiate into two types of cells: the antibody-secreting plasma cells and the memory B-cells. The plasma cells form the basis of primary immune response, which is the response mounted by the immune system to an antigen that the animal encounters for the first time. The primary response has a characteristic lag phase, during which naive B-cells proliferate and differentiate into plasma cells and memory cells. Following this, serum antibody level increases logarithmically, reaches the peak at about day 14, remains at a plateaus for some time, then begins to drop off as the plasma cells begin to die. The memory cells remain in G0 phase, and have a much longer life than plasma cells; some memory cells persist for the life of the individual. Therefore, when the animal encounters the same antigen a second time, the population of memory cells responds rapidly to begin antibody secretion. The antibody levels peak in about 7 days, and the level of antibody is about 100 to 1,000-fold higher than that in the primary response. The immune response mounted by the animal to an antigen, which it encounters a second time is called secondary immune response. The population of memory B-cells specific for a given antigen is considerably larger than the population of corresponding naive B-cells; this accounts for some of the differences between primary and secondary immune responses (Fig. 41.4). In a similar manner, the recognition of an antigen-MHC complex by a specific mature Tlymphocyte induces its proliferation and differentiation into TH cells and CTLs (the effector cells) and into memory cells. The effector cells bring about the primary immune response, which is relatively slower; it takes about 10-14 days in mouse for rejection of a skin graft in the first instance. But when skin tissue from the same source is grafted the second time, it is rejected in about 7-9 days due to the faster action of memory T-cells. Components of immune system Organs of immune system Primary or Central Lymphoid Organs: Immature lymphocytes generated in hematopoiesis, the process of formation and development of blood cells, mature and become committed to a particular antigenic specificity within the primary lymphoid organs, namely, thymus, bursa of Fabricius (in birds) and bone marrow (in mammals). A lymphocyte becomes immuno-competent, i.e., capable of mounting an immune response only after it matures within a primary lymphoid organ. 1. Thymus: Thymus is a greyish, flat, bilobed lymphoid organ situated above the heart and extending into the neck on the front and sites of trachea. It develops from the epithelium of third and fourth pharyngeal pouches and, on maturity, acts as the site of development and maturation of lymphocytes named thymus-derived lymphocytes or T-lymphocytes or T-cells. The thymus reaches peak activity in childhood and attains its largest size at puberty. Thereafter, the thymus begins to atrophy without any apparent effect on T-lymphocyte function and is extremely small in old age. For convenience, the average weight of the thymus is 70 g in infants and its age- dependent involution leaves the thymus with an average weight of 3 g in the old age. This is probably due to the fact that T-lymphocytes are very long-lived and can circulate in the resting state for long periods of time. Each lobe of thymus is surrounded by a capsule and is divided into a series of lobules, which are separated from each other by strands of connective tissue called trabeculae. Each lobule is organized into two compartments-outer and inner. The outer component is called cortex, whereas the inner component is called medulla (Fig. 42.2). The cortex is densely packed with thymocytes, whereas the medulla is sparsely populated with thymocytes. Thymocytes develop from prothymocytes. The latter are produced in bone marrow, migrate through blood stream, enter the cortex of the thymus, and act as thymocytes. Thymocytes divide rapidly in the cortex and give rise to T-lymphocytes. Of the T-lymphocytes produced in thymus only 5% leave the thymus as viable cells. Though the reason for this apparent wasteful process is not known, some believe that it is the elimination of T-lymphocyte clones that react against self. Both the cortex and the medulla of the thymus are criss-crossed by a three dimensional network consisting of epithelial cells, dendritic cells, and macrophages, which make up the framework of the organ and contribute to the growth and maturation, of thymocytes. Some epithelial cells of the outer cortex possess long membrane extensions that surround as many as 50 thymocytes. These cells are called nurse cells. Other epithelial cells of the cortex have long interconnecting cytoplasmic extensions that form a network and have been found to interact with many of the thymocytes when they traverse the cortex. The function of the thymus is to generate T-lymphocytes and to confer immunological competence on to them during their stay in the organ. T-lymphocytes so educated in the thymus become capable of mounting cell-mediated immune response against appropriate antigen. This is effected under the influence of the thymic microenvironment and several hormones such as thymosin and thymopentin produced by the epithelial cells of the thymus. The competent T-lymphocytes immediately move from thymus to the secondary or peripheral lymphoid organs. Surgical removal of bursa (bursectomy) from newly hatched chickens destroys their subsequent ability to produce antibodies. The B-cells mature, proliferate, and differentiate into bursa and then migrate from it and reach outer or superficial cortex of the germinal follicles and medullary cords of peripheral lymph nodes and lymphoid follicles of spleen where, following appropriate antigenic stimulation, transform into plasma cells and secrete antibodies. Like thymus, the bursal of Fabricius starts to shrink or atrophy at puberty. 3. Bone Marrow: Bone marrow is the site of origin and development of B-lymphocytes or B-cells (bone marrow derived lymphocytes) in mammals particularly in humans and mice after birth. Before birth, the yolk sac, foetal lever, and total bone marrow are the major sites of B-lymphocyte maturation. Bone marrow, therefore, is the mammalian equivalent of the bursa of Fabricius in birds. Development of B-lymphocytes (B-cells) begins with the differentiation of lymphoid stem cells into the earliest distinctive progenitor B cells (pro-B cell), which proliferate within the bone marrow filling the extravascular spaces between large sinusoids in the shaft of a bone. Proliferation and differentiation of pro-B cells into precursor B cells (pre-B cells) requires the microenvironment provided by the bone marrow stromal cells. The stromal cells within the bone marrow: (1) Interact directly with the pro-B and pre-B cells and (2) Secrete various cytokines that are required for development. Bone marrow is not the site of origin and development of B-lymphocytes (B-cells) in all mammals. In cattle and sheep, the fietal spleen is the primary lymphoid tissue wherein the maturation, proliferation, and diversification of B-cells take place during early gestation. During later gestation this function is performed by ideal Peyer’s patch, a patch of tissue embedded in the wall of the intestine. In rabbit, gut-associated tissues (e.g.. appendix) act as primary lymphoid tissue for maturation, proliferation, and diversification of B-cells. Secondary or Peripheral Lymphoid Organs: As stated earlier, the lymphocytes mature, proliferate, and differentiate in the primary or central lymphoid organs. These lymphocytes migrate therefrom via circulation to the secondary or peripheral lymphoid organs. Here they bind appropriate antigens and undergo further antigendependent differentiation. Once in the secondary lymphoid organs, the lymphocytes do not remain there but move from one lymphoid organ to another through the blood and lymphatic’s. The passage of lymphocytes facilitates the induction of an immune response. Lymph nodes and the spleen are the most highly organized secondary or peripheral lymphoid organs, whereas mucosa-associated lymphoid tissue (MALT) is the less organized lymphoid tissue. 1. Lymph Nodes: Lymph nodes are small, encapsulated, bean-shaped structures clustered at junctions of the lymphatic vessels which are distributed throughout the body. Lymph nodes contain a reticular network packed with lymphocytes, macrophages and dendritic cells, and filter out pathogenic microorganisms and antigens from the lymph. As the lymph percolates through a lymph node, any pathogen or antigen that is brought in with the lymph is trapped by the phagocytic cells and dendritic cells. A lymph node consists of three regions: the cortex, the paracortex, and the medulla (Fig. 42.3). Cortex is the outermost region and contains several rounded aggregates of lymphocytes (mostly B-lymphocytes), macrophages, and follicular dendritic cells arranged in primary follicles. Each follicle has a pale-staining germinal centre surrounded by small dark-staining lymphocytes. The deeper region lying beneath the cortex is the paracortex. It is the zone between the cortex and the medulla. Paracortex possesses large number of T-lymphocytes and also contains interdigitating dendritic cells thought to have migrated from tissues to the lymph node. Because of the presence of large number of T-lymphocytes in it. the Para-cortex is also referred to as a thymus-dependent area in contrast to the cortex which is a thymus-independent area. Medulla, the inner most region of lymph node, is more sparsely populated with lymphoidlineage cells. Of the lymphoid-lineage cells present, many are plasma cells actively secreting antibody molecules. Each lymph node has a number of lymph vessels called afferent lymphatic vessels, which pierce the capsule of a lymph node at numerous sites and empty lymph into the sub-capsular sinus. The lymph now percolates slowly inward through the cortex, paracortex, and medulla, allowing phagocytic cells and dendritic cells to trap pathogens and antigens carried by the lymph. The lymph then is drained into a single large lymph vessels called efferent lymphatic vessel that carries the lymph to the thoracic duct, which empties into a large vein in the neck. 2. Spleen: The spleen, which is about 5 inches long and 200 g in weight in adults, is an ovoid encapsulated, and the largest secondary or peripheral lymphoid organ. Spleen is specialized for trapping blood-borne antigens and is present high in the left abdominal cavity and being encapsulated, its capsule extends a number of projections, called trabeculae, into the interior resulting in the formation of compartments. These compartments are filled by two types of tissues, the red pulp and white pulp, which are separated by a diffuse marginal zone (Fig. 42.4). The red pulp consists of a network of sinusoids populated by large number of erythrocytes (red blood cells) and macrophages and few lymphocytes. In fact, red pulp is the region where old and defective erythrocytes are destroyed and eliminated. The white pulp consist of the branches of the splenic artery that make a periarteriolar lymphoid sheath (PALS) populated heavily by T-lymphocytes. Periarteriolar lymphoid sheath (PALS) is attached with primary lymphoid follicles that are rich in B-lymphocytes. The marginal zone separating the red pulp from white pulp is populated by lymphocytes and macrophages. When the blood-borne antigens enter the spleen the B- and T-lymphocytes present in periarteriolar lymphoid sheath (PALS) are initially activated. Here interdigitating dendritic cells capture antigen and present it combined with class II MHC molecules (major histocompatibility molecules) to TH cells (T helper cells). Once activated, these TH cells can then activate B- lymphocytes (B-cells). The activated B-lymphocytes, together with some TH cells then migrate to primary follicles in the marginal zone. When the primary follicles are challenged by antigen, they differentiate into characteristic secondary follicles. The latter contain germinal centres (similar to those occurring in lymph nodes) where rapidly dividing B-lymphocytes and plasma cells are surrounded by dense clusters of concentrically arranged lymphocytes. 3. Mucosal-Associated Lymphoid Tissue (MALT): The mucous membranes lining the alimentary, respiratory, and genitourinary systems have a very large combined surface area (about 400 m2; nearly the size of a basketball court), which is constantly exposed to numerous antigens and is the major site of entry for most pathogens. These vulnerable membrane surfaces possess a group of organized lymphoid tissues which defend it from pathogens and antigens. The group of organized lymphoid tissues is known collectively as mucosal-associated lymphoid tissue (MALT). There are several types of MALT; the most studied one is the gut-associated lymphoid tissue (GALT) which includes tonsils, Peyer’s patch, appendix, and loosely organised clusters of lymphoid cells in the lamina propria of intestinal villi. Mucosal-associated lymphoid, tissue (MALT) is functionally very significant in immune system of the body because of the presence of large number of antibody-producing plasma cells in it. The number of plasma cells in MALT for exceeds that of the total of the number of plasma cells present in spleen, lymph nodes, and bone marrow. (i) Tonsils: (a) Palatine tonsils are largest sized tonsils present on either side of oropharynx. (b) Pharyngeal tonsils are present on the posterior wall of the pharynx. (c) Lingual tonsils are present on the dorsum of the posterior part of tongue.(Fig. 42.5). All the aforesaid tonsil groups are nodule-like and consist of a meshwork of reticular cells and fibres interspersed with lymphocytes, macrophages, granulocytes, and mast cells. The B-lymphocytes are organised into follicles and germinal centres. The germinal centres are surrounded by regions showing T-lymphocyte activity. However, the tonsils protect against antigens that enter through the nausal and oral epithelial routes. (ii) Peyer’s Patch: Peyer’s patches occur in the sub-mucosal layer present beneath the lamina propria lying under the epithelial layer of intestinal villi. Each Peyer’s patch is a nodule of 30-40 lymphoid follicles. Like lymphoid follicles in other sites, those that compose Peyer’s patches can develop into secondary follicles with germinal centres (Fig. 42.6). (iii) Lamina Propria: Lamina propria occurs under the epithelial layer of intestinal villi (Fig. 42.6). It is populated with large number of plasma cells, macrophages, activated T helper cells (activated TH cells) in loose clusters. More than 15,000 lymphoid follicles have beer, reported within the lamina propria of a healthy child. Cells of immune system Cell # 1. Hematopoietic Stem Cell: All blood cells arise from a type of cell called hematopoietic stem cell (HSC) (or stem cell). The stem cells are self-renewing, maintain their population by cell division, and differentiate into other cell types. This process of formation and development of blood cells (red and white blood cells) is called hematopoiesis. It is remarkable that every functionally specialised, mature blood cell is derived from the same type of hematopoietic stem cell. In contrast to a unipotent cell, which differentiates into a single cell type, a hematopoietic stem cell is multi-potent or pluripotent as it is able to differentiate in various ways and thereby gives rise to various type of blood cells. In humans, the formation and development of blood cells begins in the embryonic yolk sac during the first weeks of development. The hematopoietic stem cells differentiate into primitive erythroid cells that contain embryonic haemoglobin. In the third month of gestation, hematopoietic stem cells migrate from the yolk sac to the foetal liver and then to the spleen. Liver and spleen play major roles in hematopoiesis from the third to the seventh months of gestation. In later months, hematopoietic stem cells differentiate in the bone marrow and play major role in hematopoiesis, and by birth there is little or no hematopoiesis in the liver and spleen. Multi-potent hematopoietic stem cell (or stem cell) in the bone marrow differentiates to form two lineages: (1) Common-lymphoid progenitor cell and (2) Common myeloid progenitor cell (Fig. 42.7). The progenitor cells, unlike hematopoietic stem cell that is self-renewing, loss the capacity for self-renewal, and are committed to their specific cell linkage. The common lymphoid progenitor cells give rise to B-lymphocytes (B-cells) that differentiate into antibody secreting plasma cells. T-lymphocytes (T-cells) that become activated T-cells. natural killer (NK) cells, and some dentritic cells. The common myeloid progenitor cells give rise to erythroblasts that produce erythrocytes (red blood cells), megakaryoblasts that produce platelets (thrombocytes), myeloblasts that produce granulocytes (eosinophils, basophils, neutrophils), monoblasts that differentiate into monocytes which give rise to macrophages and dendritic cells, and an unknown precursor that produces mast cells. However, B-lymphocytes (B-cells) T-lymphocytes (T-cells) and natural killer (NK) cells produced by lymphoid progenitor cell lineage and eosinophils, basophils, neutrophils, macrophages, and dendritic cells produced by myeloid progenitor cell lineage are collectively called white blood cells or leucocytes (Gk. leucos = white, kytos = cell). White blood cells or leucocytes are the cells that are responsible for nonspecific and specific immunity in the body. Cell # 2. Lymphocytes: Lymphocytes (L. lympha = water, cyte = cell) are the most important effector cells of many cells involved in specific immune response. These cells are small, round and 5-15 μm in diameter. They are found in peripheral blood, lymph, lymphoid organs, and in many other tissues. Lymphocytes constitute 20% – 40% of the white blood cell (leucocyte) population in the body and 99% of the cells in the lymph. They may be small (5-8 μm), medium (8-12 μm). and large (12-15 μm). The small lymphocytes are more numerous and may be short-lived with a life-span about two weeks or long-lived with a life-span of three years or more or even for life. Short-lived lymphocytes act as effector cells in immune response, while long-lived ones function as memory cells. Long-lived lymphocytes are mainly thymus derived. The formation and development of lymphocytes, i.e.. lymphopoiesis takes place in bone marrow, primary or central lymphoid organs, and secondary or peripheral lymphoid organs. Lymphocytes are approximately 1011 in number in a human body; their number ranges from 1010 to 1012 depending on body size and age. Lymphocytes can be broadly subdivided into three populations: B-lymphocytes or B-cells, T-lymphocytes or T-cells, and null cells (natural killer cells or NK cells are included in this group). 1. B-Lymphocytes or B-Cells: B-lymphocytes or B-cells derive their name from their site of maturation. They are so named since they were first detected in the bursa of Fabricius of birds and later from bone marrow of a number of mammalian species, including humans and mice. In birds, the multi-potent hematopoietic stem cells originating in the bone marrow migrate to the bursa of Fabricius and differentiate there into antibody synthesizing cells. The bursa is a small pouch-like organ in the embryonic hind-gut of birds and is absent in mammals. In a number of mammalian species including humans and mice, the B-cells originate in the foetal lever and later migrate to the bone marrow which becomes the site for production of B-cells after embryonic life. B-lymphocytes do not have the ability to synthesize antibody molecule during undifferentiated stage. During differentiation, each lymphocyte acquires the ability to synthesize antibody molecules when provoked by antigens. 2. T-Lymphocytes or T-Cells: T-Lymphocytes or T-cells derive their name from their site of maturation in the thymus. They are major players in the cell-mediated immune response and also have an important role in Bcell activation. T-cells themselves do not secrete antibodies (immunoglobulin) like B-cells. They are immunologically specific and are directly involved in cell-mediated immune responses, can carry a vast repertoire of immunologic memory, and can function in a variety of effector and regulatory way. The main effector functions include tuberculin reaction (delay-ed hypersensitivity response), destruction of tissue grafts, secretion of soluble chemical mediators called lymphokines and their ability to perform killer functions of other cells. The regulatory functions involve their cooperation with B-lymphocytes to produce antibodies. In additions to these functions, some subpopulations of T-cells contribute immune responses such as cytotoxicity, suppression, and killer properties. Like B-lymphocytes, T-lymphocytes have specific receptors on the plasma membrane surface for antigen. The receptors on T-cell membrane are called T-cell receptors (TCRs). Although T-cell receptor (TCR) is structurally distinct from immuno-globulin (the membrane receptor of B-lymphocyte), it does share some common structural features with the immunoglobulin molecule, most notably in the structure of its antigen- binding site. Unlike the membrane-bound antibody on B-cells that recognise free antigen, the T-cell receptor (TCR) does not recognize free antigen instead it recognizes the hound one to particular class of a self-molecule (e.g., major histocompatibility complex molecule or MHC molecule) displayed on self-cells (e.g., antigen presenting cells or APCs, virus-infected cells, cancer cells, and grafts). It is the T-cell system that helps eliminating these altered self-cells that threaten the normal functioning of the body. Cell # 3. Monocytes: Monocytes (G. monos = single; cyte = cell) are mononuclear phagocytic leucocytes possessing an oval or kidney-shaped nucleus and granules in the cytoplasm that stain grey-blue (Fig. 42.8). Monocytes are produced in bone marrow. During hematopoiesis in bone marrow, granulocytemonocyte progenitor cells differentiate into pro-monocytes, which-leave the bone marrow and enter the blood where they further differentiate into mature monocytes. Mature monocytes circulate in the blood stream for about eight hours, enlarge in size, migrate into the tissues, and differentiate into specific tissue macrophages or into myeloid dendritic cells. Cell # 4. Macrophages: Macrophages (G. macros = large; phagein = to eat), as noted above, are differentiated from monocytes into the tissues of the body. Differentiation of a monocyte into a tissue macrophage (Fig. 42.9) involves a number of changes: (i) The monocyte enlarges five- to ten-fold, (ii) Its intracellular organelles increase in both number (especially lysosomes and phagolysosomes) and complexity, (iii) The cell acquires increased phagocytic ability, (iv) Produces higher levels of hydrolytic enzymes, (v) Begins to secrete a variety of soluble factors, and (vi) Develops ruffles or microvilli on the surface of its plasma membrane. Macrophages are transported throughout the body. Some macrophages reside in particular tissues and become fix macrophages. Others remain motile by amoeboid movement throughout the body and are called free or wondering macrophages. Macrophages serve different functions i different tissues and are named according to their tissue location, e.g., histiocytes in connective tissues, osteoclasts in bone, microglial cells in the brain, alveolar macrophages in the lung, kupffer cells in the liver, and mesangial cells in the kidney. Macrophages normally remain in a resting state and are activated for effective functioning. They are activated by a variety of stimuli such as interferon gamma (IFN-γ) secreted by activated T helper (TH) cells, mediators of inflammatory response, components of bacterial cell walls, etc. Activated macrophages secrete different types of cytotoxic proteins that help them eliminate large number of pathogens including vims-infected cells, malignant cells, and intracellular bacteria. Activated macrophages also display class II MHC molecules that allow them to act more effectively as antigen-presenting cells (APCs). Thus, macrophages and T helper (TH) cells facilitate each other’s activation during the immune response. Macrophages are highly phagocytic and they are capable of ingesting and digesting exogenous antigens (e.g., whole microorganisms and insoluble particles) and exogenous matter (e.g., injured or dead host cells, cellular debris, activated clotting factors). Cell # 5. Granulocytes: Granulocytes (Fig. 42.10) are those white blood cells (leucocytes) which have irregular-shaped nuclei with two to five lobes and granulated cytoplasmic matrix. Granules of cytoplasmic matrix contain reactive substances that kill microorganisms and enhance inflammation. Granulocytes are also called polymorphonuclear leucocytes (PMNs). Three types of granulocytes are recognised in the body and they are: basophils, eosinophils, and neutrophils. 1. Basophils: Basophils (G. basis = base; philein = to love) possess bi-lobed irregular-shaped nucleus and cytoplasmic matrix granules that stain bluish-black with basic dyes (e.g., methylene blue). These granulocytes are non-phagocytic cell that function by releasing pharmacologically active substances (e.g., histamine, prostaglandins, serotonin, and leucotrienes) from their cytoplasmic granules upon appropriate stimulation. Since these pharmacologically active substances influence the tone and diameter of blood vessels, they are collectively termed vasoactive mediators. Basophils possess high-affinity receptors for immunoglobulin-E (IgE) antibody and thereby become coated with these antibodies. Once coated, antigens trigger the basophil cells to secrete vasoactive mediators which are inflammatory and play a major role in certain allergic responses (e.g., eczema, hay fever, and asthma). Basophils, however, comprise less than 1 % of white blood cells, are non-motile, and remain confined to the blood stream. 2. Eosinophils: Eosinophils (G. eos = dawn; philein = to love) have a bi-lobed nucleus connected by a slender thread of chromatin and prominent acidophilic granules in cytoplasmic matrix. Eosinophils, like neutrophils, are motile cells that migrate from bloodstream into tissue spaces. These granulocytes are considered to play a role in the defence against parasitic organisms (protozoans and helminth parasites) by phagocytosis. They release mainly cationic proteins and reactive oxygen metabolites into the extracellular fluid. These substances damage the plasma membrane of the parasite. Eosinophils constitute only 3-5% of the white blood cells and their acidophilic granules stain red with acidic dyes. 3. Neutrophils: Neutrophils (L. neuter – neither; philein = to love) possess a three- to five-lobed nucleus connected by slender threads of chromatin, and contain fine primary and secondary granules in cytoplasmic matrix. Neutrophils, like eosinophils, are motile cells that migrate from bloodstream into the tissue. These granulocytes circulate in the bloodstream for 7 to 10 hours before their migration into the tissues where they enjoy a life span of only a few days. Approximately 60% of the circulating white blood cells (leucocytes) in human are the neutrophils. Like macrophages, the’ primary function of neutrophils is phagocytosis of foreign or dead cells and pinnocytosis of pathological immune complexes. Phagocytosis by neutrophils is similar to that operated by macrophages except that the lytic enzymes and bactericidal substances in neutrophils are contained within primary and secondary granules instead of lysosomes in macrophages. The primary granules are larger and denser and contain peroxidase, lysozyme, and various hydrolytic enzymes. The secondary granules are smaller and contain collagenase, lactoferrin, and lysozyme. Both primary and secondary granules fuse with phagosome, whose contents are then digested and the remains excreted much as they are in macrophages. Neutrophils, like macrophages, also use oxygen-dependent and oxygen-independent pathways to generate antimicrobial substances and defensins to kill ingested microorganisms. Neutrophils generate more reactive oxygen intermediates and reactive nitrogen intermediates and express higher levels of defensins than macrophages do. Cell # 6. Dendritic Cells: Dendritic cells constitute only 0.2% of WBCs (leucocytes) in the blood and are present in even smaller numbers in skin and mucous membranes of the nose, lungs, and intestines. They derive their name due to long membrane extensions resembling the dendrites of nerve cells. Dendritic cells arise from hematopoietic stem cells in the bone marrow via different pathways and in different locations (Fig. 42.11); they descend through both the myeloid and lymphoid lineages. Stem cell-originated dendritic cells are of four types: Langerhans cells, interstitial dendritic cells, myeloid dendritic cells, and lymphoid dendritic cells. Despite differences, all the stem cell-originated mature dendritic cells perform the same major function of presenting antigen to T helper (TH) cells by expressing high levels of both class II MHC molecules and members of the co-stimulatory B-7 family, and thereby play an important accessory role in the specific immune response. This pattern of functioning makes dendritic cells more potent antigen-presenting cells (APCs) than macrophages and B-lymphocytes, both of which need to be activated before they can function as antigen-presenting cells (APCs). In addition to dendritic cells originated in bone marrow, there are another type of dendritic cells, the follicular dendritic cells, that do not arise in bone marrow and perform their function in a different ways as they do not express class II MHC molecules and do not act as antigenpresenting cells (APCs). Follicular dendritic cells express high levels of membrane receptors for antibody; which allows the binding of antibody complexes. The interaction of B-lymphocytes with this bound antigen can have important effects on B-lymphocyte responses. Cell # 7. Mast Cells: Mast cell precursors originate in the bone marrow and are released into the blood as undifferentiated cells. Mast cells are not differentiated from their precursors until the latter leave the blood and enter the tissues. Mast cells are found in a variety of tissues including the skin, connective tissues of various organs, and mucosal epithelial tissue of the respiratory, genitourinary, and digestive tracts. These cells, like basophils, possess large numbers of granules in cytoplasmic matrix. The granules in cytoplasm contain histamine and other pharmacologically active substances that contribute to the inflammatory response. Mast cells, together with basophils, play an important role in the development of allergies and hypersensitivities. Cell #8.Natural Killer Cells: These cells are mostly derived from the large granular lymphocytes. Most surface antigen of NK are shared with T cells or cells of the myelomonocytic series. NK cells are able to kill certain tumour cells and are also cytotoxic for virus infected cells. NK cells may also release interferon-7 and other cytokines (immunological mediators) which may be important in the regulation of hemopoiesis and immune responses. NK cells have other important surface molecules which are common to all leucocytes. These surface molecules are important for cell adhesion and intercellular communication. Structure of Antibody (Ab) Molecule Polypeptide chains: Antibody molecules have a common structure of four polypeptide chains, having two different sizes. These are a pair of identical high molecular weight chains called Heavy chains (H-chains) and a pair of identical low molecular weight chains called Light chains (L-chains). Each light and heavy chain may be subdivided into homologous regions termed domains. H-chains: H-chains have a molecules weight of 50-55 kd. Each H-chain has disulfide linkages (-S-S-) and also contains carbohydrate molecule attached to its asparagine residue. Each H-chain has N-terminal (NH3+) and C-terminal (Coo–) respectively. L-chains: L-chains have a molecular weight of 20-25kd. L-chains are covalently linked with H-chains by disulfide bridge (-S-S-). Each L-chain also has N-terminal and C-terminal respectively. Hinge region: Electron microscopy of purified immunoglobulin (IgG) molecules, after negative staining reveals the Y-shape of the molecules indicating a flexible “hinge region” at about the middle of the H-chains where two H-chains are connected by disulfide bridge (-S-S-). Antigen combinding site (ACS): One variable region of a heavy chain (VH) and one variable domain of light chain (VL) together constitute an antigen combinding site (ACS) or in other words determine the antibody specificity. Structure of Immunoglobulin Molecule (Ab) based on Amino Acid sequencing Studies: Polypeptide chains: Amino acid sequencing study reveals that each heavy and light chain in an immunoglobulin molecule contains an amino terminal variable (V) regions (VH → variable region of H-chain; VL → → variable region of L-chain) that consist of 100-110 amino acids and differ from one antibody to the next. The remainder of the molecule—the constant region (CH → constant region of Heavy-chain; CL → constant region of Light-chain) exhibits limited variation that defines two Light chain subtypes (k & λ) and five Heavy chain subtypes (γ, α, μ, δ, or ε). Heavy and Light chains are folded into domain, each containing about 110 amino acid residues and an inter-chain disulfide bonds that forms a 60 amino acids loop. Hinge region: Amino acid analysis of the hinge region has indicated an unusual feature—a large no. of proline residues present. Because of its structure, proline prevents the peptide chains from assuming an α-helix conformation and thus the hinge region remains extended. And the peptide bonds of the hinge region are accessible to the proteolytic enzymes shown below: Antigen binding site: Within the variable region of both Light and Heavy chain (VL and VH), amino acids at reversal positions are often substituted and at certain points, the substitution is at a notably higher frequency. These high frequency-regions are termed as “hyper-variable regions” or “hot spots”. Three regions on the Light (VL) chain and three on the Heavy (VH) chain lie relatively close to each other to form the antigen binding site. Each hyper-variable region consists of five to ten amino acid residues. As the hyper-variable regions determine the combinding sites for antigenic determinant, that’s why they are also termed as complementarity-determining regions (CDRs). The variable sequences on either side of the hyper-variable regions termed as framework regions (FRs). Thus each variable (V) region consists of three CDRs and four FRs. Humoral immune response The immune system protects the body from potentially harmful substances by recognizing and responding to antigens. Antigens are foreign particles, normally large or small molecules on the surface of cells, viruses, fungi or bacteria. Some non-living substances such as toxins, chemicals, drugs and foreign particles can also be antigens. Substances that contain these antigens are recognized and destroyed by the immune system. One of the most important immune response is humoral effector response. The effector functions in humoral immunity are mainly mediated by secreted antibodies. It protects body from extra-cellular pathogenic agents by combining with them to form antigen-antibody complex and gradually leads towards their elimination. Humoral immunity combats extracellular bacteria, fungi and even obligate intra-cellular microbes e.g. viruses; before they infect their target T-cells. Any defect in humoral immunity results in increased susceptibility to infection with bacteria and fungi. Ways involved in Humoral Immunity: Humoral effector functions facilitate effective elimination of foreign pathogens from a host animal in a variety of ways. Antibodies play vital role in elimination of antigenic agents: (i) The antibody can bind to the surface epitopes of the antigen making it more susceptible to phagocytosis—known as opsonization. (ii) The antibody molecule can bind to the antigen forming an antigen-antibody complex, which then combines with the complement in a step-wise manner to initiate and facilitate phagocytosis of the antigen. (iii) The antibody can bind to toxin molecules elaborated by microbes making them nontoxic. (iv) Antibodies can inactivate free virus particles by combining with the epitopes on viral particles to make them incapable of attachment to host cell membranes. (v) Binding to potential pathogens at mucous membrane surfaces, preventing colonization. (vi) Binding to Fc (fragment crystalized) receptors on NK cells or macrophages in antibody dependent cell mediated cytotoxicity (ADCC), confirming specificity for antigen. Humoral Immune Responses: Most defenses that are mediated by antibody present in the plasma, lymph and tissue fluids are called humoral immune responses. It protects against extra-cellular bacteria and foreign macromolecules. Transfer of antibodies confers this type of immunity on the recipient. Humoral immune responses have an activation phase and an effector phase. These phases occur as follows (Fig. 10.1): 1. The antigen is taken up by phagocytosis and degraded in a lysosome in an APC, such as a macrophage. 2. A T-cell receptor recognizes processed antigen bound to a class II MHC protein on the macrophage. 3. Cytokines released by the TH cell and IL-1 released by macrophage stimulate the TH cell to produce a clone of differentiated cells capable of interacting with B-cells. Activation phase occurs in lymphatic tissue. 4. B-cells are also antigen presenting cells. Binding of antigen to a specific IgM receptor triggers receptor mediated endocytosis, degradation and display of the processed antigen on class II MHC proteins. 5. When a TH cell receptor binds to the displayed antigen—MHC II complex on the B cell, it releases cytokines. 6. These cytokines cause the B-cell to produce a clone of B-cells. 7. Now, these B-cells produce antibody secreting plasma cells. 5 Major Classes of Immunoglobulin | Immunology Class # 1. IgG: (a) It constitutes 75% of the total serum immunoglobulin in human. (b) During the secondary immune response, it is the major Ig to be synthesized. Hence, it plays a vital role in the defense against infection.: (c) It is the only Ig class that can cross the placenta. Hence it is responsible for the protection of the neonate during the first few months of life. (d) It diffuses readily into extra-vascular spaces and hence provides a major defense against bacterial toxins and other blood born infectious agents. (e) Organisms coated with IgG attract macrophages via their FC region receptors thus enhancing phagocytosis. (f) The complement binding site on the IgG molecule appears to be on the CH-II region. (g) Ig is unable to bind onto the mast cells but has the ability to bind guinea-pig skin— the significance remains unclear. (h) The property of Fc portion of IgG to bind to protein-A on the surface of Staphylococcus aureus has been greatly exploited for use in diagnosis and research. Class # 2. IgA: (a) It is actively secreted by mucosal associated lymphoid tissue (MALT). (b) It appears selectively in sero-mucous secretions such as saliva, tears, nasal fluids and in the secretions of the lung and also in GI tracts and in UG system. (c) It is present in fluids as a dimer stabilized against proteolysis by combination with another protein, the secretory compound (J-chain) which is synthesized by local epithelial cells and has a single peptide chain of MW. 60 kd. (d) The IgA chain is synthesized locally by plasma cells and dimerized intra-cellularly before secretion with the help of J-chain. (e) It is actively endocytosed and transported within the endocytic vacuole and the mucosal surface. Cleavage of the receptor releases the IgA, still attached to the part of receptor termed the secretory piece, into sero-mucous secretion. (f) IgA is the most abundant in body secretions. It performs the role of defending the exposed external surfaces of the body against the attack of the microorganisms. (g) IgA activates complements by the alternative pathway. (h) It, the prime functional units of MALT, facilitates passage through epithelial cells and protects the secretory molecule from proteolytic degestion. Class # 3. IgD: (a) It is present in serum in trace amounts. (b) Because of an extended hinge region it is relatively liable to degradation by proteolytic enzymes. (c) The main functions of IgD has not yet been determined with IgM, it is found abundantly on the surface of B-lymphocytes. It has been suggested that they may operate as antigen receptors and in the control of lymphocytic activation and suppression. Class # 4. IgM: (a) It is the largest Ig in science and exists as a pentamer of the basic 4-chain subunit, held together by disulfide bond. (b) A relatively small molecule, the J-chain participates in the polymerization of IgM via SH residue near the C-terminal. (c) The heavy-chain of IgM are designated as μ-chains. (d) Electron microscopic study reveals that it is shaped like a star but when it is touched to a bacterium, its antigen binding sites (Fab) are bound to the bacterial surface. This changes the appearance of IgM to a crab like form, and causes cross linking of the different antigenic determinants (epitopes) on the bacterial cell-surface by polyvalent IgM molecule. (e) IgMs appear early in response to injection and because of their size are largely confined to their blood stream. (f) They are an important defense mechanism against bacteria. (g) The size and valency of IgM makes it a very effective, agglutinating and cytolytic agent. (h) Since it does not cross the placenta its presence in blood vessels indicates active foetal injection. (i) Since IgM response is short lived, its presence may be helpful in establishing an acute injection. Class # 5. IgE: (a) It is the present in very low concentration in serum. (b) IgE antibody has a very high affinity for mast cells and binding occurs via Fc portion of the Ig molecule. (c) On contact with specific antigens called allergens, the mast cells undergo degranulation with release of vasoactive amines (Histamine). This process is responsible for skin reaction in allergy for the symptoms of Hay fever and an extrinsic asthma. (d) IgE also have the ability to attach to human skin where they probably bound to mast cells. (e) It is found mainly in lining of the respiratory and GI tracts where they form constituents of MALT. (f) The main physiological role of IgE appears to be the protection of external mucosal surface. (g) Infection agents penetrating the IgA defenses, combined with specific IgE on the mast cell surface to trigger the release of vasoactive agents and other factors, chemotactic for granulocyte. (h) It is possible that IgE acts in the way as a defense against helminth infection which is characterised by an extra-aggregated IgE response. Cell-Mediated Immune Responses The cell-mediated or cellular immunity is that where the T-lymphocytes destroy other cells having antigens on their surface without any mediation by antibodies. The precursors of Tlymphocytes produced by stem cells of bone marrow pass through liver and spleen before reaching the thymus where they are processed, hence called thymus-dependent (T) lymphocytes. These lymphocytes come under the influence of the hormone “thymosin” and become immunologically competent and are called lymplioblasts. When stimulated by an appropriate antigen, the lymphoblasts divide and differentiate into cytotoxic T-lymphocyte (killer TIymphocytes), helper T-cells, and suppressor T-cells. The cytotoxic T-lymphocytes, in addition with other T-lymphocytes, release biologically active soluble factors collectively called lymphokines which act as a biochemical mediators of cellular immunity. Unlike B-lymphocytes which are normally stimulated by free antigens in the circulatory system of the body, the cytotoxic T-lymphocytes possess specific cell surface proteins, called T-cell receptors, on their surface and respond to only major histocompatibility complex antigens (MHC-antigens) bound to the surface of other cells. After the interaction between T-cell receptor and MHC-antigen is established and the cytotoxic T-lymphocyte cells binds the MHC-antigen containing cell, the latter undergoes lysis and is phagocytised (Fig. 41.2). The cell-mediated immunity is important in controlling those infections where the pathogens are intracellular and reproduce within the infected cells (e.g., viruses, rickettsia, chlamydia, some protozoans like Trypanosomes, etc.). In such infections the antibodies (hence the antibody-mediated or humoral immunity) prove to be ineffective because the antibodies are unable to penetrate and attack intracellular pathogens multiplying within the host cells. In addition, the cellular immunity is considered to play an important role in monitoring and regulating the proliferation of abnormal type of cells, (e.g., would be tumor cells), and thus, inhibiting the tumor development. Types of T cell T cells are grouped into a series of subsets based on their function. CD4 and CD8 T cells are selected in the thymus, but undergo further differentiation in the periphery to specialized cells which have different functions. T cell subsets were initially defined by function, but also have associated gene or protein expression patterns. Depiction of the various key subsets of CD4-positive T cells with corresponding associated cytokines and transcription factors. Helper CD4+ T cells T helper cells (TH cells) assist other lymphocytes, including maturation of B cells into plasma cells and memory B cells, and activation of cytotoxic T cells and macrophages. These cells are also known as CD4+ T cells as they express the CD4 on their surfaces. Helper T cells become activated when they are presented with peptide antigens by MHC class II molecules, which are expressed on the surface of antigen-presenting cells (APCs). Once activated, they divide rapidly and secrete cytokines that regulate or assist the immune response. These cells can differentiate into one of several subtypes, which have different roles. Cytokines direct T cells into particular subtypes. CD4+ Helper T cell subsets Cell type Cytokines Produced Key Transcription Factor Role in immune defence Role in autoimmunity Th1 IFNγ Tbet Produce an inflammatory response, key for defense MS, Type against intracellular bacteria, diabetes viruses and cancer. Th2 IL-4 GATA-3 Aid the differentiation and Asthma and other antibody production by B allergic diseases cells 1 Th17 IL-17 RORγt Defense against gut Rheumatoid pathogens and at mucosal Arthritis, Psoriasis barriers Th9 IL-9 IRF4, PU.1 Defense against helminths Multiple Sclerosis (parasitic worms) Tfh IL-21, IL-4 Bcl-6 Help B antibody cells produce Asthma and other allergic diseases Cytotoxic CD8+ T cells Cytotoxic T cells (TC cells, CTLs, T-killer cells, killer T cells) destroy virus-infected cells and tumor cells, and are also implicated in transplant rejection. These cells are defined by the expression of CD8+ on the cell surface. These cells recognize their targets by binding to short peptides (8-11AA) associated with MHC class I molecules, present on the surface of all nucleated cells. CD8+ T cells also produce the key cytokines IL-2 and IFNγ, which influence the effector functions of other cells, in particular macrophages and NK cells. Memory T cells Antigen-naïve T cells expand and differentiate into memory and effector T cells, after they encounter their cognate antigen within the context of an MHC molecule on the surface of a professional antigen presenting cell (e.g. a dendritic cell). Appropriate co-stimulation must be present at the time of antigen encounter for this process to occur. Historically, memory T cells were thought to belong to either the effector or central memory subtypes, each with their own distinguishing set of cell surface markers (see below). Subsequently, numerous new populations of memory T cells were discovered including tissue-resident memory T (Trm) cells, stem memory TSCM cells, and virtual memory T cells. The single unifying theme for all memory T cell subtypes is that they are long-lived and can quickly expand to large numbers of effector T cells upon re-exposure to their cognate antigen. By this mechanism they provide the immune system with "memory" against previously encountered pathogens. Memory T cells may be either CD4+ or CD8+ and usually express CD45RO. Memory T cell subtypes: Central memory T cells (TCM cells) express CD45RO, C-C chemokine receptor type 7 (CCR7), and L-selectin (CD62L). Central memory T cells also have intermediate to high expression of CD44. This memory subpopulation is commonly found in the lymph nodes and in the peripheral circulation. (Note- CD44 expression is usually used to distinguish murine naive from memory T cells). Effector memory T cells (TEM cells and TEMRA cells) express CD45RO but lack expression of CCR7 and L-selectin. They also have intermediate to high expression of CD44. These memory T cells lack lymph node-homing receptors and are thus found in the peripheral circulation and tissues. TEMRA stands for terminally differentiated effector memory cells reexpressing CD45RA, which is a marker usually found on naive T cells. Tissue resident memory T cells (TRM) occupy tissues (skin, lung, etc..) without recirculating. One cell surface marker that has been associated with TRM is the intern αeβ7, also known as CD103. Virtual memory T cells differ from the other memory subsets in that they do not originate following a strong clonal expansion event. Thus, although this population as a whole is abundant within the peripheral circulation, individual virtual memory T cell clones reside at relatively low frequencies. One theory is that homeostatic proliferation gives rise to this T cell population. Although CD8 virtual memory T cells were the first to be described, it is now known that CD4 virtual memory cells also exist. Regulatory CD4+ T cells Regulatory T cells are crucial for the maintenance of immunological tolerance. Their major role is to shut down T cell-mediated immunity toward the end of an immune reaction and to suppress autoreactive T cells that escaped the process of negative selection in the thymus. Two major classes of CD4+ Treg cells have been described — FOXP3+ Treg cells and FOXP3− Treg cells. Regulatory T cells can develop either during normal development in the thymus, and are then known as thymic Treg cells, or can be induced peripherally and are called peripherally derived Treg cells. These two subsets were previously called "naturally occurring", and "adaptive" or "induced", respectively.[18] Both subsets require the expression of the transcription factor FOXP3 which can be used to identify the cells. Mutations of the FOXP3 gene can prevent regulatory T cell development, causing the fatal autoimmune disease IPEX. Several other types of T cell have suppressive activity, but do not express FOXP3. These include Tr1 cells and Th3 cells, which are thought to originate during an immune response and act by producing suppressive molecules. Tr1 cells are associated with IL-10, and Th3 cells are associated with TGF-beta. Recently, Treg17 cells have been added to this list. Natural killer T cell Natural killer T cells (NKT cells – not to be confused with natural killer cells of the innate immune system) bridge the adaptive immune system with the innate immune system. Unlike conventional T cells that recognize peptide antigens presented by major histocompatibility complex (MHC) molecules, NKT cells recognize glycolipid antigen presented by CD1d. Once activated, these cells can perform functions ascribed to both Th and Tc cells (i.e., cytokine production and release of cytolytic/cell killing molecules). They are also able to recognize and eliminate some tumor cells and cells infected with herpes viruses. Mucosal associated invariant MAIT cells display innate, effector-like qualities. In humans, MAIT cells are found in the blood, liver, lungs, and mucosa, defending against microbial activity and infection. The MHC class I-like protein, MR1, is responsible for presenting bacterially-produced vitamin B metabolites to MAIT cells. After the presentation of foreign antigen by MR1, MAIT cells secretes pro-inflammatory cytokines and are capable of lysing bacterially-infected cells. MAIT cells can also be activated through MR1-independent signaling. In addition to possessing innate-like functions, this T cell subset supports the adaptive immune response and has a memory-like phenotype. Furthermore, MAIT cells are thought to play a role in autoimmune diseases, such as multiple sclerosis, arthritis and inflammatory bowel disease, although definitive evidence is yet to be published. Gamma delta T cells Gamma delta T cells (γδ T cells) represent a small subset of T cells which possess a γδ TCR rather than the αβ TCR on the cell surface. The majority of T cells express αβ TCR chains. This group of T cells is much less common in humans and mice (about 2% of total T cells) and are found mostly in the gut mucosa, within a population of intraepithelial lymphocytes. In rabbits, sheep, and chickens, the number of γδ T cells can be as high as 60% of total T cells. The antigenic molecules that activate γδ T cells are still mostly unknown. However, γδ T cells are not MHC-restricted and seem to be able to recognize whole proteins rather than requiring peptides to be presented by MHC molecules on APCs. Some murine γδ T cells recognize MHC class IB molecules. Human γδ T cells which use the Vγ9 and Vδ2 gene fragments constitute the major γδ T cell population in peripheral blood, and are unique in that they specifically and rapidly respond to a set of nonpeptidic phosphorylated isoprenoid precursors, collectively named phospho antigens, which are produced by virtually all living cells. The most common phosphor antigens from animal and human cells (including cancer cells) are isopentenyl pyrophosphate (IPP) and its isomer dimethylallyl pyrophosphate (DMPP). Many microbes produce the highly active compound hydroxy-DMAPP (HMB-PP) and corresponding mononucleotide conjugates, in addition to IPP and DMAPP. Plant cells produce both types of phosphor antigens. Drugs activating human Vγ9/Vδ2 T cells comprise synthetic phosphor antigens and amino bisphosphonates, which upregulate endogenous IPP/DMAPP. T-Cell Receptor T lymphocytes or T cells respond only to peptide fragments of protein antigens that are displayed by self-MHC molecules (major histocompatibility complex). T cell receptor or TCR differs from the B cell receptor in two important ways. First, the T cell receptor is membrane bound and does not appear in a soluble form as the B cell receptor does; second, the T cell receptor is specific not for antigen alone but for antigen combined with a molecule encoded by MHC. Further, the T cell receptor remains associated on the membrane with a signal-transducing complex CD3 which is non-covalently linked to the receptor to form the TCR complex. The TCR is a clonally distributed receptor, meaning that clones of T cells with different specificities express different TCRs. The biochemical signals that are triggered in T cells by antigen recognition are transduced not by the TCR itself but by TCR complex. T cells also express other membrane receptors that do not recognise antigen but participate in responses to antigens; these are collectively called accessory molecules. These molecules deliver signals to the T cell that function in concert with signals from the TCR complex to fully activate the cells. Antigen recognition by T cells is specific not only for antigen but also for an MHC molecule. T cells were shown to recognise antigen only when presented on the membrane of APC (antigen presenting cell) by a self-MHC molecule. This attribute called self-MHC restriction, distinguishes of antigen recognition by T cells from that by B cells. Two models were proposed to explain the MHC restriction of T cell receptor. The dual-receptor model proposed that a T cell has two separate receptors, one for antigen and one for class I or class II MHC molecules. The altered self-model proposed that T cell possesses a single receptor capable of recognising foreign antigen, bound to a self-MHC molecule. However, the elegant experiments by J. Kappler and P. Marrack provided supports for the altered self-model. Unlike the dual-receptor model, in which an antigen and MHC molecule are recognised separately, the altered self-model predicts that a single receptor recognises an alteration in self-MHC molecules induced by their association with foreign antigens. Structure of T Cell Receptors: The antigen receptor of MHC restricted CD4+ helper T cells and CD8+ cytotoxic T cells is a heterodimer consisting of two trans-membrane polypeptide chains. These chains are designated as α and β which are covalently linked to each other by disulfide bonds (Fig. 6.58). Another group of TCR, found on a small subset of T cells, has y and δ chains. In the amino terminal, each a chain and β chain consists of the Ig-like variable domain (V), one constant domain (C), hydrophobic trans-membrane region and a short cytoplasmic region. Thus, the extracellular portion of αβ heterodimer in TCR is structurally similar to the antigen binding fragment (Fab) of an Ig molecule. The α and β chains of V regions of TCR contain short stretches of amino acids where the hypervariable or complementarity determining regions (CDRs) are located. Three such CDRs in the α chain are juxtaposed to three similar regions in the β chain to form the part of the TCR that specifically recognises peptide-MHC complexes. The P chain V domain contains a fourth hyper-variable region, which does not appear to participate in antigen recognition but is the binding site for microbial products called super antigens. The C regions of both α and β chains continue into short hinge regions, which contain cysteine residues that contribute to a disulfide bond linking the two chains. The hinge is followed by a hydrophobic trans membrane portion of 21 or 22 amino acids. In a chain, positively charged amino acid residues like lysine and in (3 chain lysine or arginine residues are present in the trans membrane portion. Both α and β chains have carboxyl terminal cytoplasmic tails that are 5 to 12 amino acids long. Each TCR chain, like Ig heavy and light chains, is encoded by multiple gene segments that undergo somatic rearrangements during the maturation of the T lymphocytes. In α and β chains of the TCR, the third hyper-variable regions (CDR3) are composed of sequences encoded by V and J (joining) gene segments (in the α chain) or V, D (diversity) and J segments (in the β chain). The CDR3 regions also contain sequences that are not present in the genome but are encoded by different types of nucleotide additions, so-called N regions and P nucleotides. Thus, most of the sequence variability in TCRs is concentrated at CDR3. CDRs of T Cell Receptor and their Role in MHC-associated Peptide Recognition: Different experimental studies have established that both α and β chains of TCR form a single hetero-dimeric receptor that is responsible for both antigen specificity and MHC restriction of a T cell. The antigen binding site of the TCR is a flat surface formed by the CDRs of α and β chains. The TCR contacts the peptide-MHC complex in a diagonal orientation, fitting between the high points of MHC α helices. In general, the CDRl loop of the TCR α and β chains are positioned over the ends of the bound peptide. The CDR2 loops are over the helices of the MHC molecule and the CDR3 loop is positioned over the centre of the MHC-associated peptide. In fact, the side chain of only one or two amino acid residues of the MHC bound peptide make contact with the TCR. T cells have very remarkable ability to distinguish among diverse antigens on the basis of very few amino acid differences. The affinity of the TCR for peptide-MHC complex is very low. Such low affinity of specific antigen binding is the likely reason that accessory molecules are needed to stabilise the adhesion of T cells to APCs. The TCR and accessory molecules in the T cell plasma membrane move coordinately with their ligands in the APC membrane to form a transient supra-molecular structure that is referred to as immunological synapse. This structure regulates the TCR-mediated signal transduction. Virtually all αβ TCRexpressing T cells are MHC restricted and express either the CD4 or the CD8 co-receptors. A small population of T cells also expresses markers that are found on NK (natural killer) cells; these are called NK-T cells. T Cell Receptor Complex: TCR CD3: Experiments by J. P. Allison and L. Lanier and others demonstrated that T cell receptor and another protein CD3 are located quite close together in the T cell plasma membrane (within 11.5 nm of each other). The expression of CD3 molecule is required for membrane expression of αβ and yδ T cell receptors; thus each heterodimer forms a complex with CD3 on the T cell membrane. Loss of genes encoding either CD3 or TCR chains results in the loss of the entire molecular complex from the membrane. CD3 is a complex of five invariant polypeptide chains that associate to form three dimers: a heterodimer of gamma and epsilon chains (yɛ), a homodimer of delta and epsilon chains (δɛ), and a heterodimer of two zeta chains (ϚϚ) or a heterodimer of zeta and eta chain (Ϛƞ) (Fig. 6.59). The Ϛ and ƞ chains, though encoded by the same gene, may differ in their carboxyl terminal ends because of differences in RNA splicing of the primary transcript. About 90% of the CD3 complexes examined to date have ϚϚ homodimer than heterodimer Ϛƞ as possessed by rest 10% of CD3 complexes. In CD3 complex, the y, δ and ɛ chains belong to immunoglobulin superfamily, each containing an extracellular domain followed by a trans membrane region and a cytoplasmic domain of more than 40 amino acid residues. The zeta and eta (Ϛ and ƞ) chains have quite different structure, each with a very short extracellular region of only 9 amino acids, a trans membrane region and a long cytoplasmic tail of 113 amino acids in Ϛ chain and 155 amino acids in ƞ chain. The trans membrane segment of all CD3 polypeptide chains contains a negatively charged amino acid residue of aspartic acid. Such residues enable the CD3 complex to interact with one or two positively charged amino acid residues in the trans membrane segment of each TCR chain. The cytoplasmic domains of CD3 Ƴ, ɛ and δ chains contain one copy of a conserved sequence motif called the immuno-receptor tyrosine-based activation motif (ITAM) in each chain. An ITAM contains two copies of the sequence tyrosine-X-X-leucine (X is an unspecified amino acid) separated by six to eight residues. ITAM plays a central role in signaling by TCR complex. They are also found in Ϛ chain of the TCR complex, Iga and IgP proteins associated with membrane Ig molecules of B cells. Functions of CD3 complex: The CD3 and Ϛ chains link antigen recognition by the TCR to the biochemical events that lead to functional activation of the T cells. The earliest intracellular event that occurs in T cells after antigen recognition is the phosphorylation of tyrosine residues within the ITAMs in the cytoplasmic tails of CD3 and Ϛ proteins. This phosphotyrosines then become the docking sites for adapter proteins and for tyrosine kinase with a kinase called ZAP-70 that binds to the Ϛ chain. Another kinase that also docks at phosphotyrosine is Fyn that binds to CD3. Subsequent activation of these kinases triggers signal transduction pathway that ultimately lead to changes in gene expression in the T cells. yδ TCR: In a few T cell populations, a second type of diverse, disulfide linked heterodimer yδ TCR receptor is expressed instead of αβ TCR. This receptor also remains associated with CD3 and Ϛ proteins. (This yδ TCR should not be confused with the y and δ chains of CD3 complex). The TCR y and δ chains contain extracellular V and C domains, short connective or hinge regions, hydrophobic trans-membrane segments and short cytoplasmic tails. The constituents of these segments are almost similar to those of α and β chains. Furthermore, TCR – mediated signaling events typical of αβ expressing T cells are also observed in yδ T cells. However, majority of yδ T cells do not express CD4 or CD8. The percentages of yδ TCR expressing T cells vary widely in different tissues and species, but overall, less than 5% of all T cells express this receptor. T cells with yδ TCR are a lineage distinct from the αβ-expressing MHC- restricted T cells. Many δy T cells present in different organs, may have different V regions, indicating that these subsets may be specific for different ligands. One intriguing feature of yδ T cells is their abundance in epithelial tissue of certain species (e.g., small bowel mucosa of mice and chicken). In human only about 10% of intestinal intra-epithelial T cells express the yδ receptors. The function of the yδ T cells is not clear. yδ T cells do not recognise MHC-associated peptide antigens and are not MHC restricted. Some can recognise small phosphorylated molecules, alkyl amines or lipids that are commonly found in association with “non-classical” class I MHC-like molecules in mycobacteria and other microbes. Others may recognise protein or nonprotein antigens that do not require processing or any particular type of APCs for their presentation. Some suggest that they may initiate immune responses to a small number of common microbes that frequently encounter at epithelial boundaries between the host and the external environment. Accessory Molecules of T Cell Receptor: T cells express several integral membrane proteins that play important role in antigen recognition and T cell activation. Some of these molecules strengthen the interaction between T cells and antigen presenting cells or target cells; some act in signal transduction and some do both. These protein molecules are often collectively called accessory molecules. (i) CD4 and CD8: Mature αβ T cells express either CD4 or CD8 membrane protein, but not both. CD4 and CD8 interact with class II and class I MHC molecules, respectively, when the antigen receptor of T cells specifically recognise peptide-MHC complexes on APCs. Both CD4 and CD8 are trans membrane glycoprotein members of the Ig superfamily, with similar functions but different structures. CD4 is a 55-kDa monomeric protein that contains four extracellular domains (D1-D4), a hydrophobic trans membrane segment and a long cytoplasmic tail of 38 basic amino acids (Fig. 6.60). It binds through its two N-terminal domains to non-polymorphic β2 domain of the class II MHC molecule. Most CD8 molecules exist as disulfide-linked heterodimers composed of two related chains called CD8α and CD8β. Both the a and 8 chains have a single extracellular domain, a hydrophobic trans membrane region and a highly basic cytoplasmic tail of about 25 amino acids long (Fig. 6.60). The extracellular domain of CD8 binds to the non-polymorphic α3 domain of class I MHC .molecules. Some T cells express CD8αα homodimers, but this form appears to function like the more common CD8αβ heterodimers. Functions of CD4 and CD8: CD4 binds to class II MHC molecules and is expressed on T cells whose TCRs recognise complexes of peptide and class II MHC molecules. Most CD4+ class II-restricted T cells are cytokine-producing helper cells (TH cells) and function in host defence against extracellular microbes. CD8 binds to class I MHC molecules and is expressed on T cells whose TCRs recognise complexes of peptide and class I MHC molecules. Most CD8+ class I-restricted T cells are CTLs (cytotoxic T lymphocytes) which serve to eradicate infections by intracellular microbes. However, in humans, some CD4+ T cells may function as CTLs, but even these are class II restricted. Thus, expression of CD4 or CDS determines the MHC restriction of the T cells and not their functional capabilities. CD4 and CD8 participate in the early signal transduction events that occur after T cell recognition of peptide MHC complexes on APCs. This signal transduction is mediated by a T cell specific Src family tyrosine kinase called Lak that is non-covalently but tightly associated with cytoplasmic tails of both CD4 and CD8. This kinase is also required for T cell maturation and activation. CD4 and CDS promote the adhesion of MHC-restricted T cells to APCs or target cells expressing peptide MHC complexes. However, in such strengthening function, both coreceptors need the help of other accessory molecules. The CD4 act as a receptor for the human immunodeficiency virus. (ii) CD28 and CTLA-4: CD28 is a homodimer membrane protein that is expressed on more than 90% of CD4+ T cells and on 50% of CD8+ T cells in humans. Binding of B7 molecules on APCs to CD28 delivers signals to the T cells that induce the expression of anti-apoptotic proteins, stimulate production of growth factors and other cytokines and thus promote T cell proliferation and differentiation. Thus CD28 is the principal receptor for delivering signals for T cell activation. A second receptor for B7 molecule was discovered later and called CTLA-4. It is structurally homologous to CD28 but is expressed on recently activated CD4+ and CD8+ T cells. It inhibits T cell activation by counteracting signals delivered by CD28. Thus, CTLA-4 is involved in terminating T cell responses. (iii) CD45, CD2: CD45, a cell surface glycoprotein is believed to play a role in T cell activation. Various forms of CD45 are expressed on immature and mature leucocytes like T and B cells, thymocytes, mononuclear phagocytes and polymorphonuclear leucocytes. CD2 is a glycoprotein present on more than 90% of mature T cells, 50% of thymocytes and on NK cells. It functions both as an intercellular adhesion molecule and as a signal transducer. (iv) Adhesion molecules: Some accessory molecules of T cells function as intercellular adhesion molecules and play important roles in the interactions of T cells with APCs. These molecules also help in the migration of T cells to sites of infection and inflammation. The major adhesion molecules of T cells include integrins, selectins and CD44. The major functions of integrins are to mediate adhesion of T cells to APCs, endothelial cells and extracellular matrix proteins. The T cell selecting mediates the migration of naive T cells into lymph nodes, where antigens are concentrated and immune responses are initiated. Selectins also help in the migration of effector and memory T cells to sites of inflammation. CD44 is responsible for the retention of T cells in extravascular tissues at sites of infection. It also causes rapid binding of activated and memory T cells to endothelium at sites of inflammation and in mucosal tissues.