Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
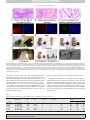
ARTICLE IN PRESS YGENO-08037; No. of pages: 8; 4C: 2, 3 Genomics xxx (2008) xxx–xxx Contents lists available at ScienceDirect Genomics j o u r n a l h o m e p a g e : w w w. e l s ev i e r. c o m / l o c a t e / y g e n o Differential methylation status of imprinted genes in nuclear transfer derived ES (NT-ES) cells Gang Chang a,b,c, Sheng Liu b, Fengchao Wang b, Yu Zhang b, Zhaohui Kou b, Dayuan Chen a,⁎, Shaorong Gao b,⁎ a b c State Key Laboratory of Reproductive Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, PR China National Institute of Biological Sciences, Beijing 102206, PR China Graduate School, Chinese Academy of Sciences, Beijing 100049, PR China a r t i c l e i n f o Article history: Received 28 July 2008 Accepted 17 September 2008 Available online xxxx Keywords: Embryonic stem cells Somatic cell nuclear transfer DNA methylation Imprinting a b s t r a c t Compared with fertilized embryo derived ES (F-ES) cells, somatic cell nuclear transfer (SCNT) produced ES (NT-ES) cells were proposed appropriate for cell transplantation based therapies. Although previous studies indicated that NT-ES cells and F-ES cells were transcriptionally and functionally indistinguishable, characterization of DNA methylation patterns of imprinted genes in NT-ES cells is lacking. Here, we show that DNA methylation patterns in the differentially methylated region (DMR) of paternally imprinted gene, H19, displayed distinct abnormalities in certain NT-ES and F-ES cell lines after long-term culture in vitro. DNA methylation profiles of H19 appeared very dynamic in most ES cell lines examined, either hypermethylation or hypomethylation could be observed in specific ES cell lines. In contrast to H19, maternally imprinted genes, Mest and Peg3, showed relatively stable methylation patterns in ES cells, especially Peg3, which displayed better capability in enduring long-term culture in vitro. Our results indicate that abnormal methylation profiles of certain imprinted genes could be observed in both NT-ES and F-ES cell lines after long-term culture in vitro although these cell lines were proved to be pluripotent with germline transmission competent. Stringent screening of epigenetically normal NT-ES cells might be potentially necessary for further therapeutic application of these cells. © 2008 Elsevier Inc. All rights reserved. Introduction Embryonic stem cells were isolated from the inner cell mass of a blastocyst and could be passaged indefinitely while maintaining selfrenewal in vitro [1–3]. Under appropriate culture conditions, ES cells could differentiate into all types of somatic cells belonging to three embryonic germ layers [4]. Given the capability of differentiation into all types of somatic lineages, ES cells have been considered as suitable seed cells for transplantation based treatment of many genetic and degenerative diseases. However, the developmental potential of ES cells is prone to differentiation, and might be influenced by long-term culture and in vitro manipulation, raising biological safety concerns on the application of ES cells for therapeutic purpose [5]. Moreover, F-ES cells were not appropriate for cell transplantation based therapy because immune rejection might occur following transplantation. Autogenic ES cells have been successfully established by SCNT in mouse [6,7] and more recently in primate rhesus monkey [8]. In ⁎ Corresponding authors. S. Gao is to be contacted at National Institute of Biological Sciences, Beijing 102206, PR China. Fax: +86 10 80727535. D. Chen, State Key Laboratory of Reproductive Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, PR China. Fax: +86 10 64807099. E-mail addresses: [email protected] (D. Chen), [email protected] (S. Gao). theory, NT-ES cells carried the same genome as the donor somatic cells; after directed induction, the differentiated cells could rescue the damaged tissues without immune rejection. In a proof of principle experiment, haematopoietic stem cells (HSCs) have been successfully differentiated from the genetically corrected NT-ES cells, and the immune deficient mice could be rescued successfully by transplantation of the induced HSCs [9]. However, unlike F-ES cells, NT-ES cells are derived from SCNT embryos in which erasing the old epigenetic marks of somatic genome and subsequently re-establishing new epigenetic marks of early embryo occur within a short time. Therefore, the NT-ES cells might inherit certain aberrant epigenetic modifications from the incompletely reprogrammed somatic genome. Results from two independent studies have shown that NT-ES cells were comparable to F-ES cells on the global transcriptional level [10,11]. However, results from previous studies also indicated that methylation patterns of imprinted genes were frequently disrupted in cloned embryos and animals [12,13]. Therefore, it is necessary to investigate whether the methylation profiles in the DMRs of imprinted genes in NT-ES cells derived from SCNT embryos are comparable to those of F-ES cells. DNA methylation is critical for gene expression, especially the monoallelic expression of imprinted genes [14,15]. The specific DNA methylation patterns in the DMRs of imprinted genes provide a convenient way to distinguish the paternal allele from the maternal 0888-7543/$ – see front matter © 2008 Elsevier Inc. All rights reserved. doi:10.1016/j.ygeno.2008.09.011 Please cite this article as: G. Chang, et al., Differential methylation status of imprinted genes in nuclear transfer derived ES (NT-ES) cells, Genomics (2008), doi:10.1016/j.ygeno.2008.09.011 ARTICLE IN PRESS 2 G. Chang et al. / Genomics xxx (2008) xxx–xxx one [16,17]. Disruption of normal imprinting has been reported leading to lethality in early embryonic development and susceptibility to many diseases in adult tissues [18–21]. In human, improper DNA methylation patterns in the DMRs of IGF2 and H19 always associated with many diseases, such as Beckwith–Wiedemann Syndrome and Wilms' tumors [18]. To provide further insights into the DNA methylation patterns of imprinted genes in NT-ES cells, we investigated the methylation status in the DMRs of two maternally imprinted genes and one paternally imprinted gene in four NT-ES cell lines using bisulfite genomic sequencing and restrictive enzyme digestion assays, comparing in parallel to two F-ES cell lines derived from the same strain of mice as NT-ES cells. Results Then we investigated whether ES cells could form chimeric mice or support full-term development of tetraploid complemented embryos, which is the most stringent method to evaluate the pluripotency of ES cells. As shown in Figs. 2G–K and in Table 1, one F-ES cell line, CL11, exhibited high chimerism and germline transmission capability. The other F-ES cell line, D1, and all four NT-ES cell lines, NT1, S5, S8 and S16, could form tetraploid blastocyst complemented fetuses, and most fetuses could survive to adulthood. In addition, tetraploid mice with malformed phenotypes were also observed, which were derived from NT1 and S5 ES cell lines. One pup had only one eye, and the other one was overgrown. As marked by a red arrow in Figs. 2H, I, these two abnormal tetraploid mice died soon after C-section. Based on the results described above, we could conclude that all ES cell lines used in our experiments were pluripotent and germline transmission competent. However, some underlying defects may exist in certain ES cell lines. Pluripotency of ES cell lines Differential methylation status of H19 in NT-ES and F-ES cells Four NT-ES cell lines and two matching F-ES cell lines derived from two different mouse strains were used in the following experiments to eliminate the sample bias. Pluripotency of each ES cell line was investigated independently by pluripotent genes expression, teratoma formation, chimeric mice generation or tetraploid blastocyst complementation. As shown in Fig. 1A, the ES cells specific genes, including Pou5f1, Sox2, Zfp42, Fut4 and Nanog all expressed in NT-ES (NT1 and S5 were used representatively) and F-ES (CL11 was used representatively) cells. To further confirm the RT-PCR results, immunocytochemistry staining of Pou5f1, Sox2 and Nanog was performed. As shown in Fig. 1B, all these three important transcriptional factors were positively expressed in both NT-ES and F-ES cells. Teratoma formation was further performed to investigate the pluripotency of ES cells used. All the ES cell lines we used could form teratomas in SCID mice. We selected the teratoma derived from NT1 ES cell line as the representative to show the HE staining and immunocytochemistry staining results. Similar results could be observed in other ES cell lines we used. The HE staining results of teratoma sections showed that immature neural tube, glia cells, skeleton muscle, column like epithelium and leprose like epithelium, belonging to three different germ layers respectively, all could be induced (Figs. 2A–C). Immunocytochemistry staining of the paraffin packaged sections of teratoma was further processed, and the results showed that the expression of Tubb3, Acta2 and Afp were all detected. Thus, the ES cells we used possessed in vivo differentiation potential (Figs. 2D–F). The methylation profiles of paternally imprinted gene, H19, were firstly investigated in two F-ES cell lines. Surprisingly, abnormal methylation patterns were observed in both F-ES cell lines derived from two different mouse strains, in which hypermethylation of H19 was observed in CL11, whereas hypomethylation of H19 was detected in D1 (Figs. 3A, B; Table 1). Next, the methylation characteristics of H19 were analyzed in NT-ES cell lines. The methylation profiles of H19 exhibited distinct variations in NT-ES cells. As shown in Figs. 3C–F and Table 1, abnormal methylation profiles were observed in all NT-ES cell lines except NT1. As control, DNA methylation profiles of H19 were determined in fertilization produced blastocysts and SCNT produced blastocysts. As shown in Figs. 3G, H, the ratio of methylated alleles to the unmethylated ones was about 1. Therefore, normal DNA methylation patterns of H19 were maintained in both fertilized and SCNT produced blastocysts. Restrictive enzyme digestion assay was further performed to confirm the bisulfite sequencing results using Taq I (Fig. 3I). As shown in Fig. 3J, the methylation level of H19 in D1 was dramatically low, in contrast, hypermethylation was observed in CL11, S5 and S16. Differential methylation status of Mest in NT-ES and F-ES cells Compared to H19, the methylation profiles of Mest maintained relatively better in ES cells examined. Hypermethylation of Mest was Fig. 1. Analyzing the expression of pluripotent genes in NT-ES and F-ES cells using RT-PCR and immunocytochemistry staining. (A) The ubiquitous expression of Pou5f1, Sox2, Zfp42, Fut4 and Nanog in CL11, NT1 and S5 ES cell lines analyzed by RT-PCR. Actin was used as inner control. (B) High expression level of Pou5f1, Sox2 and Nanog proteins in CL11, NT1 and S5 ES cell lines analyzed by immunocytochemistry staining. DNA was labeled by DAPI. Bar = 20 μm. Please cite this article as: G. Chang, et al., Differential methylation status of imprinted genes in nuclear transfer derived ES (NT-ES) cells, Genomics (2008), doi:10.1016/j.ygeno.2008.09.011 ARTICLE IN PRESS G. Chang et al. / Genomics xxx (2008) xxx–xxx 3 Fig. 2. Pluripotency analysis of NT-ES and F-ES cells by teratoma formation, chimera and tetraploid blastocyst complementation. (A–C) Various tissues which belong to three germ layers could be distinguished by HE staining of the teratoma sections. Neuron tube, immature glia cells, skeleton muscle, immature column and leprose like epithelium, all could be observed in the teratoma sections. (D–F) Expressions of Tubb3, Acta2 and Afp, which belong to three germ layers, were detected in the paraffin packaged teratoma sections by immunocytochemistry staining. (G) One F-ES cell line, CL11, had the chimerism and germline transmission ability. (H–K) Viable clonal mice can be produced by tetraploid blastocyst complementation using the four NT-ES cell lines and the other F-ES cell line, D1. Two tetraploid mice with abnormal phenotypes were also observed, which were derived from NT1 and S5 ES cell lines. As marked by a red arrow, one pup had only one eye, and the other one was overgrown. (L) The schematic diagram of blastocyst injection of ES cells. DNA was labeled by DAPI. Bar = 20 μm. observed in one F-ES cell line, CL11; while methylation pattern of Mest appeared normal in the other F-ES cell line, D1 (Figs. 4A, B). Normal methylation patterns of Mest could be observed in NT-ES cell lines including NT1, S5 and S16. However, hypermethylation of Mest was also observed in NT-ES cell line, S8 (Figs. 4C–F; Table 1). Subsequently, Rsa I was used for the restrictive enzyme digestion assay to digest the only recognized site in the DMR of Mest (Fig. 4G). As shown in Fig. 4H, heavily DNA methylation pattern of Mest could be clearly observed in CL11 and S8, while other cell lines were all within the normal arrange. Relatively normal methylation status of Peg3 in NT-ES and F-ES cells Compared to H19 and Mest, the methylation status of Peg3 was relatively stable in the ES cell lines tested. As shown in Figs. 5A–F and Table 1, the methylation profiles of Peg3 in CL11, D1, S8, S5, and S16 appeared normal, whereas hypomethylation was only observed in NT1 ES cell line. Taq I was also used to digest the DMR of Peg3 to confirm the bisulfite genomic sequencing results (Fig. 5G). As shown in Fig. 5H, among these ES cells, hypomethylation of Peg3 was only detected in NT1. Table 1 Genetic background, pluripotency, and DNA methylation status of imprinted genes in NT-ES and F-ES cells ES cell Genetic background Cell type Donor cell Tetraploid formation Germline transmission Chimerism CL11 (XY) NT1 (XX) S5 (XY) D1 (XX) S8 (XY) S16 (XY) C57 BL/6×DBA/2 C57 BL/6×DBA/2 C57 BL/6×DBA/2 C57 BL/6×129/sv C57 BL/6×129/sv C57 BL/6×129/sv F-ES NT-ES NT-ES F-ES NT-ES NT-ES – Cumulus Sertoli – Sertoli Sertoli ND Y Y Y Y Y Y Y Y Y Y Y Y ND Y ND ND ND Methylation level of imprinted genes (%) H19 Mest Peg3 64.1 41.5 69.2 27.2 35.9 66.7 72.9 56.2 58.5 44.1 71.9 44.5 46.7 15.4 46.9 41.1 36.9 48.5 NT: nuclear transfer; Y: determined; ND: not determined. Please cite this article as: G. Chang, et al., Differential methylation status of imprinted genes in nuclear transfer derived ES (NT-ES) cells, Genomics (2008), doi:10.1016/j.ygeno.2008.09.011 ARTICLE IN PRESS 4 G. Chang et al. / Genomics xxx (2008) xxx–xxx Fig. 3. DNA methylation status of H19 in blastocysts and ES cells analyzed by bisulfite genomic sequencing and restrictive enzyme digestion (Taq I). From (A) to (H), the bisulfite genomic sequencing results of CL11, D1, NT1, S8, S5, S16, NT-B and F-B were shown schematically (filled circles represent methylated CpG island). NT-B: nuclear transfer derived blastocyst; F-B: fertilized blastocyst. (I) The recognized site of Taq I in the DMR of H19 was schematically showed. (J) The gel electrophoresis results of Taq I digestion. Both digested PCR products (+) and undigested products (−) were subjected to gel electrophoresis. From left to right, NT1, S8, S5, S16, D1 and CL11 were listed sequentially. The results presented above demonstrated that the methylation profiles in the DMRs of imprinted genes investigated all appeared abnormal more or less in both NT-ES and F-ES cells, and these abnormal imprints observed were not confined to either NT-ES cells or F-ES cells after long-term culture in vitro. Discussion Maintenance of correct epigenetic modifications in NT-ES cells might be potentially important for further therapeutic application of these autogenic NT-ES cells. The results presented in the current study indicate that abnormal methylation patterns of imprinted genes in certain NT-ES cells were observed after long-term culture in vitro, and not incidentally, malformed phenotypes were observed in the tetraploid complemented mice derived from certain NT-ES cells. Epigenetic regulatory circuitry includes the modifications of core histones, incorporation of specific histone variants, DNA methylation and non-coding small RNAs. In vertebrate, DNA methylation plays a critical role in embryonic development, genomic imprinting, cell differentiation and tumorigenesis. It has been shown previously that neither parthenogenetic nor androgenetic embryos could develop to term except the specific imprinted genes being modified [22–25]. Improper imprints of Igf2 and H19 have been reported to correlate with many diseases [18]. And the dysfunction of Mest often led to postnatal growth retardation in human [26]. H19, Mest and Peg3 are well-studied imprinted genes in mouse oocytes and pre-implantation embryos, and the DNA methylation patterns inherited from gametes could be maintained during development [27,28]. Moreover, they all display classical DNA methylation patterns in the DMRs of the parental alleles, in which one allele is hypermethylated and the other one hypomethylated [16,17,29]. Thus this methylation model allowed us a convenient way to distinguish the parental alleles. More attentions have been caught to evaluate the similarities between NT-ES and F-ES cells, which are critical for further application of the NT-ES cells in regenerative medicine. Recent studies indicated that no obvious difference was observed on the global gene expression level between NT-ES and F-ES cells [10,11]. However, whether any epigenetic abnormalities, such as abnormal methylation profiles of specific imprinted genes, could be inherited or raised during culture was poorly understood in NT-ES cells. According to our present results, DNA methylation in the DMR of H19 appeared very unstable, and dynamic methylation patterns were observed among most NT-ES and F-ES cell lines. Aberrant DNA methylation patterns of Mest were also found in S8 and CL11 ES cell lines; however the aberrancy of Mest was not ubiquitous. In contrast to H19 and Mest, the methylation patterns of Peg3 could be maintained better in most ES cell lines tested after long-term culture in vitro, with only NT1 as an exception. Our results coincided with a previous report in which aberrant expression of imprinted gene, H19, was observed in both fertilized embryos derived ES cells and the ES cells produced cloned mice [30]. Based on previous findings and our results, we conclude that there is no significant correlation between ES cells pluripotency and abnormal methylation pattern of any single imprinted gene. However, certain NT-ES cells did cause the malformed phenotypes in its tetraploid complemented mice. Although we could not correlate these defects directly with the abnormal methylation of imprinted genes detected, the methylation abnormalities remain a potential risk for further therapeutic application. Long-term culture with fetal bovine serum (FBS) has been considered as one major factor influencing the stability of DNA methylation of imprinted genes, and it has been reported that epigenetic integrities of ES cells could be affected by long-term culture [5,31,32]. Moreover, results from recent studies have also shown that abnormal methylation patterns of certain imprinted genes could be observed in non-human primate ES cells after long-term Please cite this article as: G. Chang, et al., Differential methylation status of imprinted genes in nuclear transfer derived ES (NT-ES) cells, Genomics (2008), doi:10.1016/j.ygeno.2008.09.011 ARTICLE IN PRESS G. Chang et al. / Genomics xxx (2008) xxx–xxx 5 Fig. 4. DNA methylation status of Mest in ES cells analyzed by bisulfite genomic sequencing and restrictive enzyme digestion (Rsa I). From (A) to (F), the bisulfite genomic sequencing results of CL11, D1, NT1, S8, S5 and S16 were shown schematically (filled circles represent methylated CpG island). (G) The recognized site of Rsa I in the DMR of Mest was schematically showed. (H) The gel electrophoresis results of Taq I digestion. Both digested PCR products (+) and undigested products (−) were subjected to gel electrophoresis. From left to right, NT1, S8, S5, S16, D1 and CL11 were listed sequentially. culture in vitro [33]. Therefore, abnormal DNA methylation of imprinted genes did not occur only in murine ES cells after longterm culture in vitro. We hypothesize that different imprinted genes might possess distinct capacity to endure the culture effects, but the underlying mechanisms need to be further investigated. DNA methyltransferases (DNMTs) are responsible for establishing the differential methylation patterns of imprinted genes. Dnmt3a and Dnmt3b are responsible for the de novo methylation of DNA and formation of hemimethylated patterns, while Dnmt1 is crucial for maintenance of hemimethylated CpG sites [21]. The abnormalities observed in the imprinted genes in the current study might associate with the dysfunctions of DNMTs caused by long-term culture in vitro [34]. The methylation patterns of Igf2r, Peg3, Snrpn, and Grf1 have been found resistant to de novo methylation and overexpression of Dnmt1 [35]. Coincided with previous results, relatively stable imprints of Peg3 were also observed in most NT-ES and F-ES cells in the current study. More recently, induced pluripotent stem (iPS) cells have been generated in both mouse and human by transducing several transcription factors into differentiated somatic cells via viral transfection [36–41]. Although iPS cells can avoid the ethical problems, virus integration and oncogenes application hamper further clinical application. Meanwhile, similar to NT-ES cells we analyzed, the epigenetic characteristics of the iPS cell lines need to be further characterized before further application. In summary, our results demonstrated that abnormal methylation of certain imprinted genes existed in both NT-ES and F-ES cells, although these cell lines were proved to be pluripotent. Therefore, based on our findings, there was no direct correlation between aberrant epigenetic modifications and pluripotency of ES cells. However, detrimental effects might be induced during clinical application of these NT-ES cells in regenerative medicine. Evaluation of the epigenetic characteristics of NT-ES cells, and screening of relatively normal cell lines were proposed indispensable before clinical application of NT-ES cells based cell transplantation therapy. Materials and methods Animals and nuclear transfer The SPF grade mice were housed in the animal facility of the National Institute of Biological Sciences. All studies adhered to Please cite this article as: G. Chang, et al., Differential methylation status of imprinted genes in nuclear transfer derived ES (NT-ES) cells, Genomics (2008), doi:10.1016/j.ygeno.2008.09.011 ARTICLE IN PRESS 6 G. Chang et al. / Genomics xxx (2008) xxx–xxx Fig. 5. DNA methylation status of Peg3 in ES cells analyzed by bisulfite genomic sequencing and restrictive enzyme digestion (Taq I). From (A) to (F), the bisulfite genomic sequencing results of CL11, D1, NT1, S8, S5 and S16 were shown schematically (filled circles represent methylated CpG island). (G) The recognized site of Taq I in the DMR of Peg3 was schematically showed. (H) The gel electrophoresis results of Taq I digestion. Both digested PCR products (+) and undigested products (−) were subjected to gel electrophoresis. From left to right, NT1, S8, S5, S16, D1 and CL11 were listed sequentially. procedures consistent with the National Institute of Biological Sciences Guide for the care and use of laboratory animals. Metaphase II (MII) oocytes were collected from 8–10 weeks old female B6D2F1 (C57BL/6×DBA/2) mice. SCNT was performed as described [42,43]. Briefly, the spindle chromosome complex (SCC) with minimal volume of cytoplasm was removed by the enucleation pipette with an inner diameter of 10 μm attached to the piezo-drill micromanipulator. Either cumulus cells (presumably G1 phase) from ovulated oocytes or freshly prepared sertoli cells were employed as nuclear donors. The reconstructed oocytes were cultured in CZB medium for 1–3 h before activation treatment. The cloned constructs were activated in Ca2+-free CZB medium supplemented with 10 mM SrCl6 and 5 μg/ml of cytochalasin B for 6 h. Cloned embryos were cultured in KSOM (Chemicon) at 37 °C in an atmosphere of 5% CO2 in air. treated with 0.5% Pronase E (Sigma) to remove the zona pellucida (ZP). ZP free blastocysts were transferred into 96 multi-well plate and cultured on feeder layer in ES derivation medium, which contained 15% knock-out serum replacement (KSR, Invitrogen), 1000 U/ml LIF (Chemicon) and 50 μM PD98059 (Promega). Five days after culture, the outgrowths were mechanically separated into several pieces and replated into 96 multi-well dishes with feeder cells. Two F-ES cell lines, CL11 and D1 (genetic background of CL11 is C57BL/6×DBA/2, and D1 is C57BL/6×129/sv), and four NT-ES cell lines (genetic background of NT1 and S5 is C57BL/6×DBA/2, while genetic background of S8 and S16 is C57BL/6×129/sv) were selected for the subsequent experiments in our studies (Table 1), and ES cells used were about 20 passages. RNA extraction, reversed transcript PCR and immunocytochemistry Derivation of NT-ES cells and ES cells culture Derivation of ES cells was performed as previously described with a slight modification [1]. Briefly, expanded blastocysts were In order to investigate the expression of ES cells specific genes in the NT-ES and F-ES cells, reversed transcript PCR (RT-PCR) and immunocytochemistry staining were performed. Trizol (Invitrogen) Please cite this article as: G. Chang, et al., Differential methylation status of imprinted genes in nuclear transfer derived ES (NT-ES) cells, Genomics (2008), doi:10.1016/j.ygeno.2008.09.011 ARTICLE IN PRESS G. Chang et al. / Genomics xxx (2008) xxx–xxx was used to extract total RNA following manufacturer's instruction. Then total RNA was treated with DNase before reverse transcription using the MLV reverse transcriptase system (Promega). For PCR, EX Tag premix (TaKaRa) was used, and 1 μl cDNA was used as the template of 25 μl reaction system. The specific primers for Pou5f1, Nanog, Sox2, Zfp42, Fut4 and Actin were designed as following: Pou5f1, F: 5′ GGTGGAGGAAGCCGACAACAAT 3′; R: 5′ CCAGAGCAGTGACGGGAACAGA 3′, 437 bp; Nanog, F: 5′ AGGGTCTGCTACTGAGATGCTCTG 3′; R: 5′ CAACCACTGGTTTTTCTGCCACCG 3′, 364 bp; Sox2, F: 5′ CTGAACCAGCGCATGGACAGC 3′; R: 5′ CCTGGAGTGGGAGGAAGAGGT 3′, 330 bp; Zfp42, F: 5′ TAGATTTCCACTGTGGCTCTGGG 3′; R: 5′ CCTTCTTGAACAATGCCTATGAC 3′, 333 bp; Fut4, F: 5′ CAGATCGTGCCAACTATGAGCG 3′; R: 5′ ACGCAGCCAAGGCGATGTAG 3′, 310 bp; Actin, F: 5′ CCGGGACCTGACAGACTACCT 3′; R: 5′ ATGCCACAGGATTCCATACCC 3′, 276 bp. Immunocytochemistry staining was further employed to investigate the expression of Pou5f1, Sox2 and Nanog proteins in NT-ES and F-ES cells. ES cells grew on the gelatin coated cover slides were fixed in 4% paraformaldehyde. After permeabilization and blocking treatment, the cells were incubated with the first antibody at a dilution of 1:400. Mouse monoclonal anti-Pou5f1 antibody was obtained from Santa Cruz; Rabbit polyclonal anti-Sox2 antibody was obtained from Abcam; Rabbit polyclonal anti-Nanog antibody was purchased from Cosmo BioCo, Ltd. The appropriate secondary antibody was incubated with the samples after three times of washing. DNA was labeled by DAPI. Stained cells mounted on slides were observed on a LSM 510 META microscope (Zeiss) using Plan Neofluar 63×/1.4 Oil DIC objective and excitation wavelengths of 633 nm, 488 nm and 405 nm. Collection of each channel signal was done sequentially. For each experiment, the same detector gain, amplifier offset and pinhole parameters were used. All collected images were assembled using Adobe Photoshop software (Adobe Systems) without any adjustment of contrast and brightness to the images. Teratoma formation, chimera and tetraploid blastocyst complementation Teratoma formation was performed to evaluate the in vivo differentiation capacities of ES cells. 8 × 106 ES cells were subcutaneously injected into the forelimb of SCID-beige mice. About 4 weeks after injection, teratoma could be derived from all six ES cell lines we used. And the teratoma was examined histologically using standard protocol. In brief, teratomas were dissected, weighed and fixed in 4% formaldehyde. The fixed samples were then embedded in paraffin, and sections were processed with hematoxylin and eosin (HE) staining. Particular cell types were distinguished from the vicinal tissue cells according to their unique morphologies. Moreover, three embryonic layers were distinguished by immunocytochemistry staining of the teratoma sections with specific antibodies, including Tubb3 as the marker of ectoderm, Acta2 as the marker of mesoderm, and Afp as the marker of endoderm. Mouse monoclonal neuron specific Tubb3 antibody was obtained from Abcam; Mouse monoclonal Acta2 antibody was obtained from Sigma; Goat polyclonal Afp antibody was purchased from Santa Cruz. The method used for immunocytochemistry staining of the paraffin sections was similar to the protocols described above with minor modifications. To produce chimeric mice, about 10–15 ES cells were microinjected into the ICR blastocysts [44]. After culture for several hours, the re-expanded blastocysts were transplanted into uteri of pseudo-pregnant mice. To perform tetraploid blastocyst complementation, tetraploid embryos were first produced by the electrofusion of 2-cell stage embryos collected from mated female B6D2F1 mice [45]. ES cells were subsequently injected into the cavity of tetraploid blastocysts using piezo-actuated microinjection pipette. The tetraploid complemented embryos were cultured in KSOM for 2–3 h and then transplanted into the uteri of pseudo-pregnant mice. 7 Bisulfite genomic sequencing analysis of imprinted genes Bisulfite genomic sequencing was performed according to a previous report with a slight modification [29]. Briefly, Phenol: Chloroform:Isoamyl alcohol (25:24:1) was used to extract the DNA of feeder-free ES cells. Equal amount of DNA was used for bisulfite modification treatments using EpiTect Bisulfite Kit (Qiagen). Three independent PCR reactions were performed to eliminate the sampling bias of PCR. Furthermore, PCR products of three reactions were mixed together for sequencing and restrictive enzyme digestion assay. In order to get the mutagenized DNA, two rounds PCR were performed with fully nest primers for H19, Mest and Peg3. Takara Tag HS (TaKaRa) was used for the two rounds PCR. The following conditions were used for the first round of PCR: 6 min at 94 °C followed by 35 cycles of PCR consisting of 1 min at 94 °C, 2 min at 45 °C (H19)/48 °C (Mest)/45 °C (Peg3), 2 min at 72 °C, then 10 min at 72 °C. For the second round of PCR, 3 μl products of the first round PCR were used as the template. The PCR conditions were: 4 min at 94 °C, followed by 35 cycles of PCR consisting of 1 min at 94 °C, 1 min at 45 °C (H19)/50 °C (Mest)/45 °C (Peg3), 1 min at 72 °C, and 10 min at 72 °C. After electrophoresis in 1% agarose, the PCR products were recovered using Wizard SV Gel and PCR Clean-Up System (Promega). Then the PCR products were cloned into pMD18-T vector (TaKaRa) and sequenced using ABI PRISM 3100 Genetic Analyzer (Applied BioSystems). Methylation sensitive restrictive enzyme digestion assay Methylation sensitive restrictive enzyme digestion was conducted to analyze the DNA methylation profiles of imprinted genes. Rsa I (New England Biolabs) was used to recognize the only site (GTAC) at the DMR of Mest. Taq I (TaKaRa) was used to recognize the only site (TCGA) at the DMRs of H19 and Peg3. The final products were loaded onto 2.5% agarose containing ethidium bromide, with control groups containing equivalent amount of undigested PCR products. The electrophoresis condition is 100 V for 1.5 h. After electrophoresis, the agarose gels were visualized on a UV transilluminator. Acknowledgments We thank Jinghe Liu and Jiaoqiao Zhu for their technical assistance. We thank all the lab members for kindly comments on the manuscript. This study was supported by National High Technology Project 863 (2005AA210930). References [1] M.J. Evans, M.H. Kaufman, Establishment in culture of pluripotential cells from mouse embryos, Nature 292 (1981) 154–156. [2] G.R. Martin, Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells, Proc. Natl. Acad. Sci. U. S. A. 78 (1981) 7634–7638. [3] J.A. Thomson, et al., Embryonic stem cell lines derived from human blastocysts, Science 282 (1998) 1145–1147. [4] G. Keller, Embryonic stem cell differentiation: emergence of a new era in biology and medicine, Genes Dev. 19 (2005) 1129–1155. [5] A. Schumacher, W. Doerfler, Influence of in vitro manipulation on the stability of methylation patterns in the Snurf/Snrpn-imprinting region in mouse embryonic stem cells, Nucleic Acids Res. 32 (2004) 1566–1576. [6] M.J. Munsie, et al., Isolation of pluripotent embryonic stem cells from reprogrammed adult mouse somatic cell nuclei, Curr. Biol. 10 (2000) 989–992. [7] T. Wakayama, et al., Differentiation of embryonic stem cell lines generated from adult somatic cells by nuclear transfer, Science 292 (2001) 740–743. [8] J.A. Byrne, et al., Producing primate embryonic stem cells by somatic cell nuclear transfer, Nature 450 (2007) 497–502. [9] W.M. Rideout III, K. Hochedlinger, M. Kyba, G.Q. Daley, R. Jaenisch, Correction of a genetic defect by nuclear transplantation and combined cell and gene therapy, Cell 109 (2002) 17–27. Please cite this article as: G. Chang, et al., Differential methylation status of imprinted genes in nuclear transfer derived ES (NT-ES) cells, Genomics (2008), doi:10.1016/j.ygeno.2008.09.011 ARTICLE IN PRESS 8 G. Chang et al. / Genomics xxx (2008) xxx–xxx [10] T. Brambrink, K. Hochedlinger, G. Bell, R. Jaenisch, ES cells derived from cloned and fertilized blastocysts are transcriptionally and functionally indistinguishable, Proc. Natl. Acad. Sci. U. S. A. 103 (2006) 933–938. [11] S. Wakayama, et al., Equivalency of nuclear transfer-derived embryonic stem cells to those derived from fertilized mouse blastocysts, Stem Cells 24 (2006) 2023–2033. [12] K. Inoue, et al., Faithful expression of imprinted genes in cloned mice, Science 295 (2002) 297. [13] M.R. Mann, et al., Disruption of imprinted gene methylation and expression in cloned preimplantation stage mouse embryos, Biol. Reprod. 69 (2003) 902–9114. [14] M.S. Bartolomei, S.M. Tilghman, Genomic imprinting in mammals, Annu. Rev. Genet. 31 (1997) 493–525. [15] W. Reik, J. Walter, Genomic imprinting: parental influence on the genome, Nat. Rev. Genet. 2 (2001) 21–32. [16] A.P. Bird, CpG-rich islands and the function of DNA methylation, Nature 321 (1986) 209–213. [17] T.L. Davis, G.J. Yang, J.R. McCarrey, M.S. Bartolomei, The H19 methylation imprint is erased and re-established differentially on the parental alleles during male germ cell development, Hum. Mol. Genet. 9 (2000) 2885–2894. [18] R. Weksberg, A.C. Smith, J. Squire, P. Sadowski, Beckwith–Wiedemann syndrome demonstrates a role for epigenetic control of normal development, Hum. Mol. Genet. 12 (2003) 61–68. [19] E. Li, T.H. Bestor, R. Jaenisch, Targeted mutation of the DNA methyltransferase gene results in embryonic lethality, Cell 69 (1992) 915–926. [20] E. Li, C. Beard, R. Jaenisch, Role for DNA methylation in genomic imprinting, Nature 366 (1993) 362–365. [21] M. Okano, D.W. Bell, D.A. Haber, E. Li, DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development, Cell 99 (1999) 247–257. [22] J. McGrath, D. Solter, Completion of mouse embryogenesis requires both the maternal and paternal genomes, Cell 37 (1984) 179–183. [23] M.A. Surani, S.C. Barton, M.L. Norris, Development of reconstituted mouse eggs suggests imprinting of the genome during gametogenesis, Nature 308 (1984) 548–550. [24] T. Kono, et al., Birth of parthenogenetic mice that can develop to adulthood, Nature 428 (2004) 860–864. [25] M. Kawahara, et al., High-frequency generation of viable mice from engineered bimaternal embryos, Nat. Biotechnol. 25 (2007) 1045–1050. [26] S. Kobayashi, et al., Human PEG1/MEST, an imprinted gene on chromosome 7, Hum. Mol. Genet. 6 (1997) 781–786. [27] D. Lucifero, C. Mertineit, H.J. Clarke, T.H. Bestor, J.M. Trasler, Methylation dynamics of imprinted genes in mouse germ cells, Genomics 79 (2002) 530–538. [28] K.D. Tremblay, K.L. Duran, M.S. Bartolomei, A 5′2-kilobase-pair region of the imprinted mouse H19 gene exhibits exclusive paternal methylation throughout development, Mol. Cell. Biol. 17 (1997) 4322–4329. [29] J.H. Liu, et al., Diploid parthenogenetic embryos adopt a maternal-type methylation pattern on both sets of maternal chromosomes, Genomics 91 (2008) 121–128. [30] D. Humpherys, et al., Epigenetic instability in ES cells and cloned mice, Science 293 (2001) 95–97. [31] F.A. Eggerding, et al., Uniparental isodisomy for paternal 7p and maternal 7q in a child with growth retardation, Am. J. Hum. Genet. 55 (1994) 253–265. [32] M.R. Mann, et al., Selective loss of imprinting in the placenta following preimplantation development in culture, Development 131 (2004) 3727–3735. [33] A. Fujimoto, S.M. Mitalipov, H.C. Kuo, D.P. Wolf, Aberrant genomic imprinting in rhesus monkey embryonic stem cells, Stem Cells 24 (2006) 595–603. [34] T. Hikichi, T. Kohda, S. Wakayama, F. Ishino, T. Wakayama, Nuclear transfer alters the DNA methylation status of specific genes in fertilized and parthenogenetically activated mouse embryonic stem cells, Stem Cells 26 (2008) 783–788. [35] D. Biniszkiewicz, et al., Dnmt1 overexpression causes genomic hypermethylation, loss of imprinting, and embryonic lethality, Mol. Cell. Biol. 22 (2002) 2124–2135. [36] K. Takahashi, S. Yamanaka, Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors, Cell 126 (2006) 663–676. [37] K. Okita, T. Ichisaka, S. Yamanaka, Generation of germline-competent induced pluripotent stem cells, Nature 448 (2007) 313–317. [38] M. Wernig, et al., In vitro reprogramming of fibroblasts into a pluripotent ES-celllike state, Nature 448 (2007) 318–324. [39] K. Takahashi, et al., Induction of pluripotent stem cells from adult human fibroblasts by defined factors, Cell 131 (2007) 861–872. [40] I.H. Park, et al., Reprogramming of human somatic cells to pluripotency with defined factors, Nature 451 (2008) 141–146. [41] W.E. Lowry, et al., Generation of human induced pluripotent stem cells from dermal fibroblasts, Proc. Natl. Acad. Sci. U. S. A. 105 (2008) 2883–2888. [42] T. Wakayama, A.C. Perry, M. Zuccotti, K.R. Johnson, R. Yanagimachi, Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei, Nature 394 (1998) 369–374. [43] S. Gao, M. McGarry, K.E. Latham, I. Wilmut, Cloning of mice by nuclear transfer, Cloning Stem Cells 5 (2003) 287–294. [44] W.T. Poueymirou, et al., F0 generation mice fully derived from gene-targeted embryonic stem cells allowing immediate phenotypic analyses, Nat. Biotechnol. 25 (2007) 91–99. [45] A. Nagy, et al., Embryonic stem cells alone are able to support fetal development in the mouse, Development 110 (1990) 815–821. Please cite this article as: G. Chang, et al., Differential methylation status of imprinted genes in nuclear transfer derived ES (NT-ES) cells, Genomics (2008), doi:10.1016/j.ygeno.2008.09.011