Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
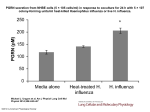
Am J Physiol Cell Physiol 298: C209 –C210, 2010; doi:10.1152/ajpcell.00528.2009. Editorial Focus From blood typing to a transport metabolon at a crossroad. Focus on “Ammonium-dependent sodium uptake in mitochondrion-rich cells of medaka (Oryzias latipes) larvae” Shigehisa Hirose1 and Tsutomu Nakada2 1 Department of Biological Sciences, Tokyo Institute of Technology, Yokohama, Japan; and 2Department of Molecular Pharmacology, Shinshu University School of Medicine, Nagano, Japan Address for reprint requests and other correspondence: S. Hirose, Dept. of Biological Sciences, Tokyo Institute of Technology, 4259-B-19 Nagatsuta-cho, Midori-ku, Yokohama 226-8501, Japan (e-mail: [email protected]). http://www.ajpcell.org proteins in pufferfish (Rhag, Rhbg, Rhcg1, and Rhcg2; lowercase refers to the proteins of nonhuman species) and determined their locations in the gill, the major site of gas exchange, ionoregulation, and ammonia elimination in adult teleost fish; it is noteworthy that gills are needed for ionoregulation before they are needed for oxygen uptake during development (16). Gill surface area is greatly increased by its lamellar structure. The ammonia transporters Rhag, Rhbg, and Rhcg2 are distributed on these lamellar cells, which is ideal for optimizing ammonia elimination. In contrast, Rhcg1 exhibits a somewhat unexpected localization in mitochondrion-rich cells (MRCs or ionocytes). MRCs are relatively minor among the population of the gill epithelial cells and are specialized to actively absorb or secrete NaCl to maintain body fluid homeostasis. MRCs are present in the skin and yolk sac during early developmental stages before functional gills are present. Since the MRCs found in the skin are easily accessible to experimental analysis, they are frequently studied. Since this report, one or more members of the Rh protein family including those mentioned above (1, 2, 7, 12, 15; for review see Refs. 24 and 27) and Rhp2 (p for primary) (14) have been identified in other fish species including zebrafish, a freshwater fish in which a knockdown experiment can be performed for detailed functional analyses of specific gene products by using antisense morpholino oligonucleotides. In zebrafish, Rhcg1 has been demonstrated to be expressed in the skin and gill MRCs that are rich in vacuolar-type H⫹-ATPase (vH-MRCs) (12, 17), and implicated as being somehow involved in Na⫹ uptake (3, 12). Apical localization of Na⫹/H⫹ exchanger 3 (NHE3) in vH-MRCs is also observed in zebrafish (29). By integrating these pieces of evidence and other information accumulated by previous molecular physiological studies, a collaborative group of researchers at the McMaster University and the University of Guelph (21, 27) has proposed a “Na⫹/NH⫹ 4 exchange complex” consisting of the transporters Rhcg, vacuolar-type H⫹-ATPase (vH-ATPase), Na⫹/H⫹ exchanger (NHE), and epithelial Na⫹ channel (ENaC) working together as a metabolon. Except for the inclusion of ENaC, which is not present in teleost genomes (22), the model is very attractive since the ammonia is not simply discarded; instead, the downhill gradient for ammonia excretion into the water is used to drive uphill Na⫹ uptake from the water into the freshwater fish. Wu et al. (28) provided a piece of supporting evidence for the model. After failing to detect significant Na⫹/NH⫹ 4 exchange in zebrafish, these investigators analyzed another model fish, Japanese medaka (Oryzias latipes). The electrophysiological technique they used warrants a comment. SIET (also known as the self-referencing ion-selective electrode 0363-6143/10 $8.00 Copyright © 2010 the American Physiological Society C209 Downloaded from http://ajpcell.physiology.org/ by 10.220.32.246 on August 11, 2017 THE RH BLOOD GROUP-RELATED PROTEINS have recently been recognized as ammonia transporters acting in erythrocyte and nonerythrocyte membranes. They play an important role in excretion of nitrogenous waste formed mainly from amino acid metabolism. Because of a restricted availability of water, most terrestrial animals detoxify ammonia into urea or uric acid before removal from the body. However, aquatic animals, living in an abundant supply of water, can directly excrete ammonia into the environment immediately as it is produced. This method of ammonia elimination is strategic in avoiding urea synthesis, an anabolic process requiring ATP. In this issue of American Journal of Physiology-Cell Physiology, Wu et al. (28) add a further twist to this adaptive strategy by providing evidence for a coupling of the ammonia excretion with a sodium uptake process, which is essential for living in freshwater conditions. The Rh antigen was first identified about 70 years ago by Landsteiner and Wiener (8), who showed that rabbits and guinea pigs, when immunized with red blood cells of Rhesus monkey, produce an antibody that also agglutinates human red blood cells (Rh for Rhesus). The Rh antigen (Rh), now recognized for its strong immunogenic nature and use for diagnosis of pregnancy at risk for hemolytic disease of the newborn, forms a complex with other subunits including Rh-associated glycoprotein (RhAG), CD47, ICAM-4, and glycophorin B (25). Rh and RhAG are the major components of the complex and belong to the same family of proteins. They were expected to carry some physiologic function based on their well-conserved nature among the species and especially because they have 12 transmembrane-spanning segments characteristic of transporters. A comparative genomic approach led to the following breakthrough discovery that RhAG acts as an ammonia transporter. A low (⬃20%) but significant sequence similarity was found by Marini et al. (10) between RhAG and the members of the Mep/Amt family previously identified as ammonia transporters in microorganisms and plants for which ammonia is a preferred source of nitrogen (Mep for methylammonium permease; Amt for ammonium transporter); for the evolution of the Mep/ Amt/Rh family, see Huang and Peng (6). RhAG and its nonerythroid homologues, RhBG and RhCG, were then demonstrated to behave indeed as ammonia transporters (9, 26). The discoveries briefly summarized above have greatly contributed to our understanding of the mechanism of ammonia secretion in fish. Nakada et al. (13) identified four Rh glyco- Editorial Focus C210 DISCLOSURES No conflicts of interest are declared by the author(s). REFERENCES 1. Braun MH, Steele SL, Ekker M, Perry SF. Nitrogen excretion in developing zebrafish (Danio rerio): a role for Rh proteins and urea transporters. Am J Physiol Renal Physiol 296: F994 –F1005, 2009. 2. Claiborne JB, Kratochvilova H, Diamanduros AW, Hall C, Phillips J, Miller E, Hirose S, Edwards S. Expression of branchial Rh glycoprotein ammonia transporters in the marine longhorn sculpin (Myoxocephalus octodecemspinosus). Bull Mt Desert Isl Biol Lab 47: 67–68, 2008. 3. Esaki M, Hoshijima K, Kobayashi S, Fukuda H, Kawakami K, Hirose S. Visualization in zebrafish larvae of Na⫹ uptake in mitochondria-rich cells whose differentiation is dependent on foxi3a. Am J Physiol Regul Integr Comp Physiol 292: R470 –R480, 2007. 4. Fredriksson S, Gullberg M, Jarvius J, Olsson C, Pietras K, Gustafsdottir SM, Ostman A, Landegren U. Protein detection using proximitydependent DNA ligation assays. Nat Biotechnol 20: 473–477, 2002. 5. Hiroi J, Yasumasu S, McCormick SD, Hwang PP, Kaneko T. Evidence for an apical Na-Cl cotransporter involved in ion uptake in a teleost fish. J Exp Biol 211: 2584 –2599, 2008. 6. Huang CH, Peng J. Evolutionary conservation and diversification of Rh family genes and proteins. Proc Natl Acad Sci USA 102: 15512–15517, 2005. 7. Hung CY, Tsui KN, Wilson JM, Nawata CM, Wood CM, Wright PA. Rhesus glycoprotein gene expression in the mangrove killifish Kryptolebias marmoratus exposed to elevated environmental ammonia levels and air. J Exp Biol 210: 2419 –2429, 2007. 8. Landsteiner K, Wiener AS. An agglutinable factor in human blood recognized by immune sera for rhesus blood. Proc Soc Exp Biol Med 43: 223–224, 1940. AJP-Cell Physiol • VOL 9. Marini AM, Matassi G, Raynal V, Andre B, Cartron JP, CherifZahar B. The human Rhesus-associated RhAG protein and a kidney homologue promote ammonium transport in yeast. Nat Genet 26: 341– 344, 2000. 10. Marini AM, Urrestarazu A, Beauwens R, Andre B. The Rh (rhesus) blood group polypeptides are related to NH⫹ 4 transporters. Trends Biochem Sci 22: 460 –461, 1997. 11. Messerli MA, Collis LP, Smith PJ. Ion trapping with fast-response ion-selective microelectrodes enhances detection of extracellular ion channel gradients. Biophys J 96: 1597–1605, 2009. 12. Nakada T, Hoshijima K, Esaki M, Nagayoshi S, Kawakami K, Hirose S. Localization of ammonia transporter Rhcg1 in mitochondrion-rich cells of yolk sac, gill, and kidney of zebrafish and its ionic strength-dependent expression. Am J Physiol Regul Integr Comp Physiol 293: R1743–R1753, 2007. 13. Nakada T, Westhoff CM, Kato A, Hirose S. Ammonia secretion from fish gill depends on a set of Rh glycoproteins. FASEB J 21: 1067–1074, 2007. 14. Nakada T, Westhoff CM, Yamaguchi Y, Hyodo S, Li X, Muro T, Kato A, Nakamura N, Hirose S. Rhesus glycoprotein p2 (Rhp2) is a novel member of the Rh family of ammonia transporters highly expressed in shark kidney. J Biol Chem. In press. 15. Nawata CM, Hung CC, Tsui TK, Wilson JM, Wright PA, Wood CM. Ammonia excretion in rainbow trout (Oncorhynchus mykiss): evidence for Rh glycoprotein and H⫹-ATPase involvement. Physiol Genomics 31: 463–474, 2007. 16. Rombough P. Gills are needed for ionoregulation before they are needed for O2 uptake in developing zebrafish, Danio rerio. J Exp Biol 205: 1787–1794, 2002. 17. Shih TH, Horng JL, Hwang PP, Lin LY. Ammonia excretion by the skin of zebrafish (Danio rerio) larvae. Am J Physiol Cell Physiol 295: C1625– C1632, 2008. 18. Smith PJ, Hammar K, Porterfield DM, Sanger RH, Trimarchi JR. Self-referencing, non-invasive, ion selective electrode for single cell detection of trans-plasma membrane calcium flux. Microsc Res Tech 46: 398 –417, 1999. 19. Smith PJ, Trimarchi J. Noninvasive measurement of hydrogen and potassium ion flux from single cells and epithelial structures. Am J Physiol Cell Physiol 280: C1–C11, 2001. 20. Soderberg O, Leuchowius KJ, Gullberg M, Jarvius M, Weibrecht I, Larsson LG, Landegren U. Characterizing proteins and their interactions in cells and tissues using the in situ proximity ligation assay. Methods 45: 227–232, 2008. 21. Tsui TK, Hung CY, Nawata CM, Wilson JM, Wright PA, Wood CM. Ammonia transport in cultured gill epithelium of freshwater rainbow trout: the importance of Rhesus glycoproteins and the presence of an apical Na⫹/NH⫹ 4 exchange complex. J Exp Biol 212: 878 –892, 2009. 22. Venkatesh B, Kirkness EF, Loh YH, Halpern AL, Lee AP, Johnson J, Dandona N, Viswanathan LD, Tay A, Venter JC, Strausberg RL, Brenner S. Survey sequencing and comparative analysis of the elephant shark (Callorhinchus milii) genome. PLoS Biol 5: e101, 2007. 23. Wang YF, Tseng YC, Yan JJ, Hiroi J, Hwang PP. Role of SLC12A10.2, a Na-Cl cotransporter-like protein, in a Cl uptake mechanism in zebrafish (Danio rerio). Am J Physiol Regul Integr Comp Physiol 296: R1650 –R1660, 2009. 24. Weihrauch D, Wilkie MP, Walsh PJ. Ammonia and urea transporters in gills of fish and aquatic crustaceans. J Exp Biol 212: 1716 –1730, 2009. 25. Westhoff CM. The structure and function of the Rh antigen complex. Semin Hematol 44: 42–50, 2007. 26. Westhoff CM, Ferreri-Jacobia M, Mak DO, Foskett JK. Identification of the erythrocyte Rh blood group glycoprotein as a mammalian ammonium transporter. J Biol Chem 277: 12499 –12502, 2002. 27. Wright PA, Wood CM. A new paradigm for ammonia excretion in aquatic animals: role of Rhesus (Rh) glycoproteins. J Exp Biol 212: 2303–2312, 2009. 28. Wu SC, Horng JL, Liu ST, Hwang PP, Wen ZH, Lin CS, Lin LY. Ammonium-dependent sodium uptake in mitochondrion-rich cells of medaka (Oryzias latipes) larvae. Am J Physiol Cell Physiol (November 25, 2009). doi:10.1152/ajpcell.00373.2009. 29. Yan JJ, Chou MY, Kaneko T, Hwang PP. Gene expression of Na⫹/H⫹ exchanger in zebrafish H⫹-ATPase-rich cells during acclimation to lowNa⫹ and acidic environments. Am J Physiol Cell Physiol 293: C1814 – C1823, 2007. 298 • FEBRUARY 2010 • www.ajpcell.org Downloaded from http://ajpcell.physiology.org/ by 10.220.32.246 on August 11, 2017 technique) is a noninvasive method for measuring minute extracellular ion gradients created within the unstirred layer on the surface of ion-transporting epithelial cells (18, 19). SIET uses a microelectrode having a tip diameter of ⬃5 m and a thin layer (⬃250 m) of an ion-selective ionophore cocktail in its tip. By a modulation technique using a vibrating electrode, which is termed self-referencing, the drift components inherent to electrodes can be removed and the electrochemical detection of signals is greatly improved. The principle is simple: A microelectrode is placed within the unstirred boundary layer of a cell; a second measurement is then made, with the same electrode attached to a vibrator, at a known distance away, and the two compared; the process is repeated with the help of a scanning device. By using fast responding electrodes, this self-referencing method is now being improved so as to characterize rapid events such as a singlechannel activity under native conditions (11). Using SIET, Wu et al. (28) first demonstrated that MRCs on the skin of medaka larvae ⫹ actually secrete NH⫹ 4 and absorb Na and then showed colocalization of Rhcg1 and NHE3 (slc9a3) in MRCs by in situ hybridization. Tight coupling of ammonia secretion and sodium uptake was also suggested by inhibition experiments and by altering compositions of bathing media. It is interesting that the rate of ammonia secretion by individual MRCs is more than 10 times higher in medaka than in zebrafish (17). Future studies should aim to provide direct evidence for the physical coupling of Rhcg1 and NHE3 (i.e., a transport metabolon). The recently developed proximity ligation assay (4, 20) may prove useful in this model. Concerning the mechanism for Na⫹ uptake by MRCs of freshwater fish, the role of the apical Na⫹-Cl⫺ cotransporter (NCC) should be considered because it has been demonstrated in another teleost fish, tilapia (5), while in zebrafish MRCs’ the NCC has been shown to be mainly involved in Cl⫺ uptake and play only an indirect or minor role in Na⫹ uptake (23).