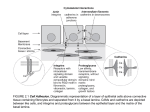
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Physiol Genomics 14: 171–197, 2003; 10.1152/physiolgenomics.00021.2002. invited review Genetic programs and responses of neural stem/progenitor cells during demyelination: potential insights into repair mechanisms in multiple sclerosis Jaime Imitola,1 Evan Y. Snyder,2 and Samia J. Khoury1 1 Center for Neurologic Diseases, Partners MS Center, Department of Neurology, Brigham and Women’s Hospital, and 2Department of Neurology, Harvard Institutes of Medicine, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston Massachusetts 02115 neural stem cells; experimental autoimmune encephalomyelitis; neuroregeneration; cytokines; neural transplantation; neurodegeneration MULTIPLE SCLEROSIS (MS) is the most commonly diagnosed neurological disease in young adults (201). This chronic debilitating disease represents a high cost to society in terms of both productivity and chronic health care resources (45). The current treatment of MS relies on immunologic manipulation. However, it is becoming clear that repair should be considered as a goal for treatment. Demyelination, axonal degeneration, neuronal dysfunction, and apoptosis (57, 157) are key features in MS pathology, and current evidence suggests that remyelination to some extent also occurs spontaneously (236, 238). As these “self-repair” mechanisms begin to be better understood, optimization and permanence of neural repair may become a target for new therapies. Here we review what is known about the role of neural stem cells (NSCs) and progenitor cells in demyelinating diseases. We will first discuss basic concepts of NSC and progenitor biology, especially oligodendrocyte progenitor cells (OPCs). We will then review the genetic programs involved in NSC and OPC proliferation, differentiation, and apoptosis, then discuss the responses of endogenous and exogenous NSCs and OPCs to the demyelinating environment Article published online before print. See web site for date of publication (http://physiolgenomics.physiology.org). Address for reprint requests and other correspondence: S. J. Khoury, 77 Ave. Louis Pasteur R710, Harvard Institutes of Medicine, Boston, MA 02115 (E-mail: [email protected]). and the emerging evidence of molecules that may be involved in such responses. Finally, we will analyze the therapeutic potential of these findings. GLOSSARY For the aid of the reader, here we briefly provide some definitions and descriptions of relevant concepts. Self-renewing capacity. This is the ability of a stem cell in vivo and in vitro to generate new stem cells with the same characteristics. Multipotency. Multipotency is the ability of a stem cell to give rise to a lineage-specific committed progenitor cell that will terminally differentiate in mature progeny. Symmetric vs. asymmetric division. Symmetric division is the generation of two clonally derived stem cells, whereas asymmetric division is the generation of a stem cell and a committed progenitor. NG2. NG2 is a proteoglycan, i.e., a marker for oligodendrocyte progenitor cells. O4. O4 is an antibody that has recognized a developmental regulated epitope, which signals the first step into terminal oligodendrocyte differentiation. Th1 cytokines. Th1 cytokines are secreted preferentially by T cells that have differentiated along a T helper 1 phenotype, including IFN-␥, lymphotoxin, and interleukin-2. Th2 cytokines. Th2 cytokines are secreted preferentially by T cells that have differentiated along a T 1094-8341/03 $5.00 Copyright © 2003 the American Physiological Society 171 Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 Imitola, Jaime, Evan Y. Snyder, and Samia J. Khoury. Genetic programs and responses of neural stem/progenitor cells during demyelination: potential insights into repair mechanisms in multiple sclerosis. Physiol Genomics 14: 171–197, 2003; 10.1152/physiolgenomics.00021.2002.—In recent years, it has become evident that the adult mammalian CNS contains a population of neural stem cells (NSCs) described as immature, undifferentiated, multipotent cells, that may be called upon for repair in neurodegenerative and demyelinating diseases. NSCs may give rise to oligodendrocyte progenitor cells (OPCs) and other myelinating cells. This article reviews recent progress in elucidating the genetic programs and dynamics of NSC and OPC proliferation, differentiation, and apoptosis, including the response to demyelination. Emerging knowledge of the molecules that may be involved in such responses may help in the design of future stem cell-based treatment of demyelinating diseases such as multiple sclerosis. 172 INVITED REVIEW BASIC CONCEPTS OF NEURAL STEM AND PROGENITORS CELLS Neural Stem and Progenitor Cells The current definition of a NSC relies upon “operational” criteria rather than on a set of molecular markers alone. NSCs are cells from the central nervous system (CNS) and peripheral nervous system (PNS) that are multipotent and self-renewing (76) (Fig. 1A). Multipotency is the ability of a single cell clone to give rise to the three major types of cells in the CNS (e.g., neurons, oligodendrocytes, and astrocytes) throughout the nervous system at all stages of life, as well as to repopulate those regions when depleted of cells. The term “self-renewal” is used to define the capacity of a clone to generate new stem cells with identical properties from generation to generation. NSCs seem to have a proliferative response to certain cytokines such as epidermal growth factor (EGF) and basic fibroblast growth factor (bFGF), although the number of such factors seems to be growing (76). “Progenitor cells” are a step further along than NSCs in the differentiation process; they have committed to a particular lineage (neuronal or glial) and have begun to express lineage-specific markers. How irreversible or invariant such commitment is remains an area of active speculation and investigation. Progenitor cells such as OPCs may, under certain circumstances in vitro, exhibit “stemness” in that they can be induced to generate neurons, astrocytes, and oligodendrocytes (18, 134, 166). Even “end”-differentiated cells have been observed to evince this property; adult ependymal cells, for example, when cultured in vitro or when monitored in vivo are reported to give rise to both neurons and glia; in vivo they yield neurons that migrate to the olfactory bulb and astrocytes that respond to injury (122), although controversy exists about the real stemness of ependymal cells. Other investigators suggest that subventricular zone (SVZ) astrocytes rather than ependymal cells are the true stem cells in that periventricular region. This finding has been clarified recently by Rietze et al. (228), who found nestinpositive pluripotent stem cells in both the subventricular and the ependymal area; interestingly, these cells were negative for glial fibrillary acidic protein (GFAP), suggesting heterogeneity of NSCs (59, 139, 228), a Physiol Genomics • VOL 14 • phenomenon that has recently been observed in human NSCs at the molecular level (259). Other investigators have found stem-like cells within a supposedly committed “glial” population extracted from the postnatal and adult cortex, adult substantia nigra, as well as the adult optic nerve; such immature “glia” give rise to neurons as well as mature oligodendrocytes and astrocytes (139, 147, 206). It remains unclear what this heterogeneity of progenitor populations implies: a true biological phenomenon or a limitation in our ability to provide cell-specific unambiguous defining markers. There are specific germinative zones in the embryonic and adult mammalian CNS (5) that can be considered as sources of NSCs. These zones include the ventricular zone, the SVZ, the external germinal layer of the cerebellum, the subgranular zone of the dentate gyrus, and the ependymal layer of the spinal cord (77). During the earliest stages of cerebrogenesis, neurons and glia are born in the ventricular zone from NSCs that become progenitors and migrate out using the radial glia as a scaffold to reach the cortex. In addition, radial glia themselves may be multipotent neural precursors with stem cell properties (86, 111, 143, 187). These same NSCs give rise to secondary germinal zones, including the SVZ, that contain a reservoir of undifferentiated, uncommitted NSCs that persist into adulthood (205). In the adult mammalian CNS, NSCs can be isolated from the SVZ, striatum, cortex, and the rostral extension of the olfactory bulb (92). Adult neurogenesis from hippocampus has been observed in humans and rodents during normal and abnormal conditions (64, 194), but the exact cells giving rise to new neurons are still under investigation. Previous studies have suggested that neurogenesis in the adult hippocampus arises from NSCs (64, 276) and that astrocytes from hippocampus are capable of regulating neurogenesis by instructing the stem cells to adopt a neuronal fate (253). These adult hippocampal NSCs are able to generate functional neurons (254). However, these findings have been challenged by recent work; Seaberg et al. (239) showed that the dentate gyrus contains multipotent NSCs during development and until birth, but these cells become restricted neuronal progenitors during adulthood. These results would suggest that what were thought to be adult NSCs from hippocampus were actually contaminating cells from the periventricular region that persist near the adult dentate gyrus and that neuronal restricted progenitors and not NSCs are the source of newly generated dentate gyrus neurons throughout adulthood. The adult SVZ (153) is a heterogeneous complex of several layers of cells adjacent to the ependyma, surrounding the lateral ventricles. So far, four different types of cells have been identified in the SVZ (Fig. 1, B and C): type A migrating neuronal precursors that migrate to the olfactory bulb; type B nestin-positive, GFAP-positive astrocytes that exhibit NSC properties; type C nestin-positive multipotent progenitors cells (61); and ependymal cells that line the ventricles and the spinal cord and that may have stem-like qualities as well (60, 122, 144). Current evidence suggests that www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 helper 2 phenotype, including interleukin-4 and interleukin-5. Lysolecithin Lysolecithin is an analog of lysophosphatidylcholine that cause destruction of oligodendrocytes and is used in a model of chemically induced demyelination. Cuprizone. Cuprizone is a toxin that targets oligodendrocytes and causes destruction of oligodendrocytes and is used in model of chemically induced demyelination. Experimental autoimmune encephalomyelitis (EAE). EAE is a model for the human disease, MS, that is achieved by immunizing a susceptible strain of rodents and primates with myelin antigens. INVITED REVIEW 173 in the adult SVZ, NSCs express GFAP (115). The developmental origin of these cells is unclear; it is suggested that these cells may evolve directly from radial glia, which are multipotent precursor cells that participate in neurogenesis as an anchor for tangential chain migration of neuroblasts (59, 60, 139, 259). Another possibility is that adult NSCs are indeed radial glia that adopt an adult developmental expression of GFAP (115). In the adult CNS, NSCs of the SVZ divide symmetrically, yielding two NSCs, one of which dies by apoptosis leaving the other to maintain the NSC pool during the life span of the individual (188). During developPhysiol Genomics • VOL 14 • ment or when external triggering occurs (such as injury), NSCs proliferate either by symmetric division, where each cell divides into two new NSCs, or by asymmetric division, where the NSC gives rise to one new NSC and a progenitor cell. When asymmetric division predominates, a great number of rapidly proliferating progenitors are generated that migrate out to their final destination. In the songbird (131), NSCs in the adult participate in the neurogenesis that is responsible for seasonal changes of the song. Although neurogenesis has been found to exist in adult mammalian brain, including humans, it is unclear what role NSCs play there, although they do seem to become www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 Fig. 1. Operational definition of a neural stem cell (NSC). A: adult stem cells can be isolated by dissection of the subventricular zone (SVZ). A single cell suspension generates multipotent cells in epidermal growth factor (EGF) and fibroblast growth factor (FGF); these cells give rise to a “primary neurosphere” that contains NSC and progenitor cells. When dissociated at the single cell level, these cells give rise to cells with identical multipotentiality (in this case, defined as a “secondary neurosphere”) to be deemed as “self-renewing.” NSCs exhibit multipotency when cultured in medium containing serum; these give rise to neuronal and glial progenitors and subsequently more mature and terminally differentiated cells. B: confocal laser microscopy of bromodeoxyuridine (BrdU) incorporation by cells in the SVZ of adult mice demonstrating proliferation of SVZ precursor cells. Bar ⫽ 100 m. CC, corpus callosum. C: representation of the types of cellular architecture in the SVZ. The ependymal cell layer (EP) lines the ventricles; these cells have been postulated to be NSCs. The SVZ is composed of a heterogeneous population of cells: neuronal precursors (“A”), astrocytes or type B cells that may also be stem-like NSCs (“B”), type C multipotent precursor cells that exhibit NSC properties in vivo and in vitro (“C”), and adult oligodendrocyte progenitor cells (OPCs, in red). 174 INVITED REVIEW integrated into neuronal circuitry of the hippocampus, possibly participating in neural plasticity (64, 274, 275) and learning. Impairing the proliferation of endogenous dentate precursors has been shown to affect behavioral learning in the mouse (245, 246). Characterization and Pluripotency of NSCs Physiol Genomics • VOL 14 • www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 NSCs will proliferate in response to EGF and/or bFGF. This proliferative response can be used to isolate and expand stem-like cells from the CNS. Any proliferative cell grown in serum-free medium without a substrate will form a floating cluster of cells. When such clusters are obtained from primary dissociated neural cultures (e.g., from the SVZ or embryonic forebrain), they may contain and help select for cells with stem-like features. Some investigators who employ this technique have termed this a “neurosphere” assay. When plated on an adherent substrate with serumcontaining medium, neurons, astrocytes, oligodendrocytes, as well as undifferentiated cells may appear. It should be emphasized that a single cluster does not equal a clone unless it has been proven to derive from a single isolated cell in an isolated well (1, 53, 55, 59, 122, 171, 188, 280). Self-renewal can be implied if a single cell can give rise to differentiated cells as well as to other undifferentiated cells that can be similarly placed into an isolated well and give rise to all differentiated and undifferentiated progeny. The above-mentioned maneuvers constitute an “operational” definition of a stem cell and remain today the only valid way of discerning a self-renewing and multipotent NSC. However, surrogate biological markers are constantly being investigated and tested for sensitivity and specificity. Nestin, an intermediate filament protein, has been useful in the study of NSCs (141, 280). Nestin, however, while sensitive, is not specific; it is expressed by a variety of cells that include reactive astrocytes, endothelial cells, SVZ neuronal progenitor cells, and ependymal cells (149, 175), as well as NSCs. It can also be expressed by non-NSCs, such as embryonic stem (ES) cells (151, 158) and pancreatic stem cells (310). Some investigators have used the low expression of peanut agglutinin (PNA)-binding and heat stable antigen (HSA) to identify a nestin-positive pluripotent NSC from the adult SVZ in mice with the phenotype nestin⫹ PNAlo HSAlo (228). Multipotent neural progenitor cells from the SVZ may also express the intermediate filament vimentin, the polysialic form of neural cell adhesion molecule (PS-NCAM), and the transcription factor mushashi (242, 304). Another proposed marker is the carbohydrated Lewis X expressed on embryonic pluripotent stem cells and found in 4% of the adult SVZ, but not in the ependymal region. These Lex-positive populations showed stem cells characteristics (41). Some studies have identified and isolated populations of NSCs based on flow cytometric characteristics in response to uptake of the DNA dye Hoechst: both red and blue wavelengths, similar to studies done in other stem cells (88, 89). The isolation is based on the differential efflux of the dye by a multidrug transporter. The population isolated in this way is called “the side population” or “SP” fraction because the cells segregate to a tiny bin that looks like a small “hook” of cells branching off from the main accumulation of cells after fluorescence-activated cell sorting (FACS). Floating clusters of cells in serum-free medium from dissociated neural cultures may segregate to this SP population (108). Furthermore, the novel hematopoietic stem cell marker CD133 has been used to define a population of human neural precursors cells that exhibit a CD133⫹ CD34⫺ CD45⫺ phenotype and fulfill all the criteria for human NSCs. However, it is not clear whether these cells are a distinct population of NSCs or simply express distinct markers (273). Other NSCs such as neural crest stem cells (NCSCs), have been prospectively isolated from fetal mouse sciatic nerve using cell surface antigens such as p75, the low-affinity neurotrophin receptor, and P0, a peripheral myelin protein: these cells exhibit phenotype p75⫹P0⫺ and are capable of self-renewal and the generation of neurons and glia (185). It is most likely that a battery of markers will be needed to define a genuine stem cell. Future work will involve defining the molecular profile or fingerprint of a NSC, much as has been derived for the hematopoietic stem cell. The term pluripotency refers to the capacity of stem cells to give rise to the full range of cells and tissues in an organism. This most extensive potential was thought to be a specific property of ES cells alone. However, several controversial observations over the past 2 yr have suggested that other stem cells, including NSCs, may have such potential under extraordinary nonphysiological circumstances. NSCs have been reported to give rise to mature nonneural cells (251). Adult murine NSCs may become hematopoietic cells after injection into an irradiated host and express such T cell markers as CD3 (22). Others have shown that, when injected into early stage mice and chick embryos, adult NSCs may differentiate into cells of the gastrointestinal tract, heart, and kidney, expressing markers typically found in these tissues in a nontumorogenic manner (55). NSCs can also produce skeletal myotubes both in vitro and in vivo (79). This behavior is also observed in other stem cells, such as bone-marrowderived stem cells that after intravenous injection in mice travel to the CNS, express neuronal markers, and generate an astroglial phenotype (27, 178). The purity of the NSC populations used in these experiments as well as the specificity of the markers used to define specific neural and nonneural cell types remain issues of ongoing investigation and inquiry in such experiments. Other groups have failed to reproduce these findings; recently, Morshead et al. (186) transplanted 128 host animals with NSCs, and they could not confirm a contribution to hematopoiesis. An alternative explanation to the Bjornson experiment (22) is that fusion rather than true transdifferentiation may explain some of the extraordinary claims of pluripotency of NSCs (264, 299). In support of this explanation is the 175 INVITED REVIEW Oligodendrocyte Progenitor Cells In the mature CNS, oligodendrocytes are terminally differentiated cells, yet new oligodendrocytes seem to originate from OPCs (probably by way of NSCs) (99) since there is production of new oligodendrocytes in the adult forebrain (293) and spinal cord (107). OPCs can be identified by several markers: the ganglioside GD3; the cell surface marker A2B5; the O4 antigen; 14F7 monoclonal antibody (301); the glycoprotein AN2 (196); the spliced form of 2⬘,3⬘cyclic nucleotide 3⬘-phosphohydrolase (CNPase) (302); the transcription factors Olig-1, Olig-2, Nkx2.2, Dlx2 (155, 222); and the integral membrane proteoglycan NG2 (142, 197). Although NG2 may be expressed by macrophages and endothelial cells, the pattern of staining and the complex morphology of OPCs helps to make the distinction (124, 215) (Fig. 2, A and B). Using these markers, researchers have detected OPCs in the adult CNS (226). Fig. 2. Oligodendrocytes developmental lineage and morphology. NG2-positive (stained red) adult OPCs from the corpus callosum can exhibit multipolar (A) and bipolar morphology (B). C: symmetrical cell division is shown in two young OPCs (red) with incorporated BrdU (green) indicative of cell division and showing typical morphology with radially oriented processes in the adult mice spinal cord. Arrows in A and B point to OPC processes and in C to BRDU-positive nucleus. Scale bar ⫽ 10 m. D: developmental stages of oligodendrocytes from a NSC. PDGF, plateletderived growth factor; Gal-C, galactocerebrosidase; PLP, proteolipid protein; CNPase, 2⬘,3⬘-cyclic nucleotide 3⬘-phosphohydrolase; MBP, myelin basic protein; MOG, myelin oligodendrocyte glycoprotein. Physiol Genomics • VOL 14 • www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 recent observation of bone-marrow-derived cells fused to human Purkinje neurons after bone marrow transplantation (284). Regardless of how confirmatory experiments for the above-cited studies turn out, the differentiation potential of stem cells, including NSCs, remains quite extraordinary. Their ability to differentiate in response to specific environmental clues, both physiological and nonphysiological, suggests a critical role for cytokines and cell-to-cell contact signals and the relative levels of transcription factors on stem cells. For example, the oct-4 gene defines pluripotency and self-renewal of ES cells (198); however, it is downregulated when ES cells change to nestin-positive stem cells (158), suggesting that different “master” transcription factors may regulate pluripotency of NSCs. Terminal differentiation likely depends upon the spatiotemporal expression and interactions of multiple cytokines with their receptors on the NSC surface that activate specific transcriptional programs (235, 271). 176 INVITED REVIEW Physiol Genomics • VOL 14 • adult OPCs in vivo; Horner et al. (107) found that less than 1% of OPCs divide and exhibit a prolonged cell cycle (Fig. 2C). It is possible that environmental cues absent in the adult brain but present in the myelinating brain or molecules enriched in the adult brain may influence this behavior; molecules like Notch and Jagged have been shown to inhibit OPC differentiation (282). Moreover, OPCs are regionally heterogeneous in their ability to differentiate; for example, cortical OPCs isolated from postnatal day 7 rats exhibit a slow differentiation behavior when compared with optic nervederived cells (216). Despite the biological difference, adult OPCs also share many of the characteristics of the perinatal OPCs, such as response to growth factors, the triiodothyronine (T3)-dependent timing for differentiation, and maintenance of their self-renewing capacity (Table 1). Adult OPCs, which can yield mature oligodendrocytes in the adult brain, express NG2, the PDGF␣ receptor, O4, and the FGF receptor (145). They also express c-kit, a marker for hematopoietic stem cells that is lost when they differentiate into postmitotic oligodendrocyte (112). Adult OPCs are maintained in a quiescent state by a mechanism that involves ␣-amino-3-hydroxy-5methyl-4-isoxazole propionate (AMPA) receptor (208), the cell cycle inhibitor p27Kip1 (44), and Notch. Activation of the Notch receptor on OPCs blocks their oligodendrocyte differentiation in the developing optic nerve (282). Because of their abundance in the CNS and the fact that their processes contact the nodes of Ranvier (19), it has been suggested that OPCs subserve synaptic remodeling and plasticity in the adult CNS (199, 291). Recent data, however, point to a broader differentiation potential and replication capacity of OPC; depending on the cytokine milieu, they may exhibit actual neural stem-like characteristics (134, 206, 261). OPCs express several functional cytokine receptors such as FGF-2R (176) and the PDGF␣ receptor (225); in fact, as noted above, such receptors have been used as markers for OPCs (218). OPCs have been isolated from brains of healthy humans and of MS patients, and these express the p75NTR (neurotrophin receptor) that is implicated in oligodendrocyte survival and apoptosis (300). In vitro oligodendroglial development can be recapitulated from NSCs; Zhang et al. (306) demon- Table 1. Characteristics of perinatal vs. adult OPCs Characteristics Perinatal Adult Proliferation Differentiation Levels of cyclin E Levels of CDK2 Levels of p27Kip1 Response to growth factors T3 dependence for differentiation Self-renewing capacity NG2 expression high high high high low yes low low low low high yes yes yes yes yes yes yes OPC, oligodendrocyte progenitor cell. www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 Around embryonic day E15, oligodendrocytes arise from a subpopulation of precursor cells within the ventral ventricular zone of the spinal cord and the SVZ (143, 200). These populations are characterized by the coexpression of the transcription factors Olig-2 and Nkx2.2, both of which are involved in OPCs differentiation and myelination (155). Reynolds and colleagues (99, 227) characterized the time course of oligodendrocyte marker expression and differentiation using the anti-ganglioside GD3 antibody; these cells arise from subependymal layers, migrate out, and subsequently express galactocerebrosidase-C (Gal-C) and the myelin-associated protein CNPase. Myelin basic protein (MBP) is expressed 2–3 days later, prior to myelin formation (99, 227). During oligodendrocyte development, OPCs undergo several maturational stages determined by specific antigenic expression and developmental potential (Fig. 2D). The OPCs isolated from the neonatal optic nerve were initially termed “O2A progenitor cells” (221) because they were determined to give rise in vitro to oligodendrocytes (“O”) as well as type 2 (fibrous) astrocytes (“2A”) depending on the concentration of serum and platelet-derived growth factor (PDGF) in the growth medium. In serum-free medium they differentiate into oligodendrocytes, whereas in serum-containing medium they become astrocytes (65). These proliferative, self-renewing, bipotent glial precursors were immunoreactive to antibodies against GD3, vimentin, A2B5, and NG2. Adult optic nerve also contains a similar type of cell that conserves some properties of its perinatal counterpart (290). In the initial stages of development, OPCs exhibit the NG2⫹ PDGFR-␣⫹ O4⫺ phenotype. These cells give rise to more advanced progenitors expressing O4, which, in turn, generate premyelinating oligodendrocytes that lose the expression of NG2 and gain the expression of proteolipid protein (PLP), and, finally, to mature oligodendrocytes expressing MBP and CNPase (65). Expression of O4 is a first step into terminal oligodendrocyte differentiation (83) and indicates a more committed stage with less proliferative potential (283). Gal-C expression is associated with terminal differentiation and is followed by MBP, PLP, and myelin/oligodendrocyte glycoprotein (MOG) expression (237). Some of the above-described precursors persist as adult OPCs. These OPCs differ from their perinatal counterparts in several respects: they divide, migrate, and differentiate 3–4 times more slowly than perinatal OPCs. These differences are likely to be cell intrinsic, but the exact mechanism of the progressive change of potential from perinatal OPCs to adult OPCs is unknown. The cell cycle inhibitor p27Kip1 accumulates as these cells divide and has been proposed as one of the mechanisms leading to the change in proliferation potential (240). Additionally, the levels of cdk2 and cyclin are diminished in adult OPCs compared with perinatal cells; this may account for the proliferative state of adult OPCs (16). However, cell intrinsic mechanisms are not sufficient to explain the dormancy of 177 INVITED REVIEW MOLECULAR PROGRAMS THAT CONTROL NSC AND OPC FUNCTION Genes Involved in NSC Proliferation The signals responsible for controlling proliferation and differentiation of precursor cells in the CNS are starting to be elucidated. Among these molecules, the cytokines EGF and bFGF (also called “FGF-2”) and their receptors are critical for the proliferation of NSCs (93, 132, 280). But many other molecules are involved (Table 2). EGF and FGF-2 signal through ERK kinases and induce the proliferation of NSCs and progenitors (242). NSC expresses both receptors at varying levels in individual cells. Furthermore, there are specific FGF or EGF-responsive NSCs. FGF-responsive NSCs are present early, and EGF-responsive NSCs emerge later in development. Both FGF-responsive and EGF-responsive NSCs retain their self-renewal and multipotency (168, 269). The activity of FGF is mediated by FGFR1, which allows the cells to undergo self-renewal mediated through a calcium-dependent mechanism (163). The acquisition of a primitive NSC identity from ES cells is mediated by leukemia inhibitory factor (LIF) (268) and is enhanced by FGF-2. LIF is involved in the maintenance of NSCs of the adult SVZ. Several cytokines [LIF, ciliary neurotrophic factor (CNTF), interleukin-6 (IL-6)] act on the common receptor gp130, and LIFR⫺/⫺ mice exhibit a decrease in number of stem cells, whereas administration of CNTF intraventricularly enhances self-renewal of NSCs in vivo (243). Transforming growth factor-␣ (TGF-␣) is also relevant to SVZ precursor cell fate. The tgf-␣⫺/⫺ mice exhibit decreased proliferation of SVZ cells, resulting in a decreased number of progenitors in the SVZ and a reduced number of neuronal progenitors migrating to the olfactory bulb. However, their stem cell number Table 2. Summary of genes related to neural stem and progenitor function Functional Categories Gene Name(s) Factors involved in NSCs proliferation Factors involved in self-renewing capacity Factors involved in NSCs neurogenic differentiation Factors involved in NSCs astrocytic differentiation Factors involved in OPCs proliferation Factors involved in OPCs differentiation Factors involved in OPCs apoptosis and survival Factors involved in OPCs migration EGF, FGF-2, LIF, IL-6, PTEN, -catenin, neuregulin, EPO, TGF-␣, IL-6, CNTF, ephrin-B2, Sonic hedgehog, Emx-2, reelin P21, N-COR PS-1, Hes-1, cystatin-C, ephrins, BDNF, IL-11, IL-5, IL-7, IL-9, FGF2, neurogenin-1, SOCS-2 BMP2-4, gp130, CNTF, STAT3 and 5, Notch, PDFG-␣, neuregulin, N-COR neuregulin, PDGF-␣, IGF-I, IL-1, GGF-2 gp130, CNTF, Notch, neuregulin, PDGF-␣, Olig-1, Olig-2, Sox-10, FGF3, histone deacetylases, thyroid hormone, Id4 Fas, TNF-␣, IL-10, PPAR, glutamate, Jagged CXCR2, Sema-3A, netrin-1, semaphorin F NSC, neural stem cell. Physiol Genomics • VOL 14 • www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 does not change (267). Conversely, intraventricular infusion of TGF-␣ increases cell proliferation in the SVZ (67). In vivo, EGF and FGF exhibit different effects on NSC. FGF-2 increases the rate of newborn cells, especially neurons, whereas EGF increases the numbers of glia in the hippocampus (136). Additionally, EGF has the capacity to convert type C progenitors (see Neural Stem and Progenitor Cells, above, under BASIC CONCEPTS. . .) cells form the SVZ into multipotent stem cells (61). FGF is also mitogenic for oligodendrocytes and OPCs (225). The importance of FGF-2 is revealed by the observation that introduction of a dominant-negative version of FGFR1 into OPCs results in cells that are not responsive to FGF-2 and that fail to migrate in vivo (203). Some members of the ephrin (Eph) family of tyrosine kinase receptors are expressed by cells of the SVZ and have been shown to be involved in the migration of neuronal progenitors in the adult SVZ. The infusion of a truncated version of ephrin-B2 into the lateral ventricle increases proliferation of NSCs and disrupts migration of neuronal progenitors (56). Other developmental molecules involved are the Sonic hedgehog, working through its receptor patched, which regulates proliferation of NSCs from the hippocampus (138); the homeobox transcription factor Emx2, which participates in symmetric division of adult SVZ stem cells (80); and reelin, which facilitates the proliferation of hippocampal NSCs (130). Hypoxia-induced cytokines such as erythropoietin (EPO) regulate the numbers of NSCs in the SVZ, promoting the production of neuronal progenitors and new olfactory bulb interneurons (244). Other molecules can act as negative regulators of NSC proliferation; pten⫺/⫺ neurospheres exhibit a greater proliferation, decreased cell death, and enlarged cell and brain size, which may suggest a role for pten in the control of neural progenitors proliferation (95). -Catenin transgenic mice show an increase in the proliferation of neural precursors and increase cerebral cortical and brain size, suggesting that this molecule plays a role in the decision of precursors to proliferate or differentiate during mammalian neuronal development (52). strated that human oligodendrocytes appear from neurospheres and express PDGF␣ receptor. The proliferation of these human OPCs was augmented by coculture with neurons and astrocytes (306). Furthermore, human neurospheres increase their yield of oligodendrocyte production when cultured with T3 (190). Finally, control of the timing of OPC differentiation requires extracellular signals such as PDGF␣ and T3 (63). 178 INVITED REVIEW CNS development but also for their vulnerability during neurodegenerative processes. Genes That Control Neuronal Differentiation Of NSCs Physiol Genomics • VOL 14 • Genes That Regulate Glial Differentiation There are several specific molecules that mediate glial differentiation; these include cytokines and transcription factors (Table 2). For example, OPCs express receptors for bone morphogenetic protein (BMP). BMP actually inhibits oligodendrocyte differentiation by OPCs and promotes their differentiation into astrocytes (94, 160). Transgenic overexpression of BMP-4 leads to increased numbers of astrocytes and reduced numbers of oligodendrocytes (87). Insulin-like growth factor I (IGF-I), neurotrophin-3 (NT-3), and gp130 receptor subunit-related ligands induce the proliferation of OPCs and postmitotic oligodendrocytes (14). In addition, different gp130-associated neuropoietic cytokines [CNTF, LIF, oncostatin-M, and hematopoietic cytokines IL-6, IL-11, IL-12, and granulocyte colonystimulating factor (GCSF)] have differential trophic effects on oligodendroglial lineage (172). Furthermore, the terminal maturation of OPCs and glial differentiation are dependent on factors that activate gp130/ LIF- receptors, such as CNTF (165). CNTF enhances the proliferation of OPC as well as the generation of oligodendrocytes in vivo (12). These effects appear to be mediated by phosphorylation of the nuclear corepressor N-Cor with its redistribution out of the nucleus into the cytoplasm. N-Cor⫺/⫺ NSCs exhibit lack self-renewing capacity and spontaneous astrocytic differentiation, establishing N-Cor as a downstream target of CNTF (102). The downstream signaling pathways responsible for differentiation after gp130 engagement involve the STAT family of transcription factors (182). IL-6 and LIF (both of which signal through gp130) modulate the growth of NSCs and progenitors in vitro via a STAT3-dependent mechanism that requires the transcriptional coactivator p300 (193). STAT5 also plays a role in the proliferation of progenitor cells after cytokine receptor stimulation (46). Additionally, IL-1 promotes mitotic arrest and oligodendrocyte differentiation of OPCs (278). The Notch family of proteins is critical for NSC and progenitor biology. Notch1 has been used to isolate ependymal stem cells (122). NSCs are depleted in both Notch⫺/⫺ and preselinin-1⫺/⫺ mice (a key regulator of Notch signaling). These mice exhibit alteration of both neuronal and glial differentiation (278). This work contrasts with many reports suggesting an involvement of Notch in the inhibition of neuronal fate while promoting glial fate or favoring astrocytic differentiation at the expense of neurogenesis (90). Notch inhibits NCSC differentiation into neurons; even a transient exposure to Notch ligands causes an irreversible change from neurogenesis to gliogenesis (184). Chambers et al. (48) studied the effects of Notch in vivo and found that an excess of activated Notch markedly inhibits the generation of neurons from precursors and increases the generation of astroglial progeny. Notch can also inhibit OPC differentiation during development (282) and inwww.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 FGF regulates the differentiation of NSCs. At low concentrations, FGF-2 induces neurogenic differentiation, whereas it induces oligodendroglial differentiation at high doses (220). FGF-2 requires a novel autocrine cofactor, a glycosylated form of cystatin C that synergizes with FGF-2, to stimulate neuronal differentiation of NSCs in vitro (263). Presenilin-1 (PS1) is involved in neuronal progenitors fate. PS1 cleaves Notch and induces an activated form that maintains the neuronal progenitors pool. PS1⫺/⫺ mice exhibit premature neuronal differentiation associated with aberrant neuronal migration and disorganization of architecture of the cerebral hemisphere, due to alteration of the notch pathway (98). Blocking of HES-1, another member of the Notch pathway, results in upregulation of p21, suppression of NSCs proliferation, and generation of GABAergic neurons (125). Other cytokines are involved in the lineage commitment of NSCs. Brain-derived neurotrophic factor (BDNF) increases the morphological and antigenic differentiation of neuronal progenitors derived from NSCs (2). Other molecules, such as basic helix loop helix (bHLH) transcription factors, are related to the differentiation of NSCs to glia and to neurons. Neurogenin promotes neurogenesis by functioning as a transcriptional activator and inhibits the differentiation of NSCs into astrocytes by inhibiting the STAT signal transduction associated with astrocytic differentiation (258). Suppressor of cytokine signaling-2 (SOCS-2), an intracellular regulator of cytokine signaling, regulates neurogenesis. SOCS-2⫺/⫺ mice generated fewer neurons and more astrocytes, and the lack of neurons was related to a decrease of the bHLH neurogenin-1. Overexpression of SOCS-2 increased neurogenesis by blocking growth hormone-mediated downregulation of neurogenin-1 (272). Hematopoietic cytokines also regulate NSC differentiation. Using immortalized hippocampal stem cells, Mehler et al. (173) showed that cytokines that participate in T cell maturation, immunoglobulin synthesis, and hematopoiesis induce progressive neuronal differentiation. The cytokine IL-11 induces the differentiation of NSCs to immature neurons. However, IL-5, IL-7, and IL-9 induce differentiation of NSCs into more mature neurons when combined with TGF-␣ after pretreatment with FGF-2. These findings may be of relevance in the context of the potential interaction between immune cell-derived cytokines and precursor cells during immune-mediated demyelination and injury (Table 2). Thus several immune-related cytokines play a role in the proliferation of NSC and progenitor cells. These molecules may be involved during CNS inflammatory pathological states such as brain and spinal cord injury and MS (15). Therefore, these cytokines and chemokines and their receptors may not only be critical for INVITED REVIEW Physiol Genomics • VOL 14 • Genome-wide Screening of Neural Stem and Progenitor Cells The functional genetic programs of NSC and OPC generation, proliferation, and differentiation are being defined. The introduction of global genomic analysis and gene profiling is helping to define novel genes. Geschwind et al. (85), using microarrays derived from subtractive hybridization between undifferentiated and differentiated progenitors, found novel and known genes differentially expressed in the germinative zone and the NSC clone C17-2 (249). Some of the known genes with differential expression in the germinative zone could provide potential markers for NSCs such as PGAM1 (phosphoglycerate mutase); CAL1H (calpactin 1 heavy chain); Hcyclin D2; cyclin D1; MELK (maternal embryonic leucine zipper kinase) (32). There are genes that appear to be also enriched in hematopoietic stem cells, suggesting overlapping genetic programs between neural and hematopoietic stem cells. Among such genes is Cyt28, a novel transmembrane protein member of the secretin subfamily of seven transmembrane receptors (7TM), found in the SVZ and also expressed in fetal liver stem cells. Other genes such as Ku80, and cyclin D1 are part of a common genetic program of hematopoietic and NSCs (265). Although this approach may yield data about novel genes critical for stem cell function, it is possible that known genes that are important for other stem cell populations, such as cytokine and chemokine receptors, may also be functionally expressed in these cells. Recently, two groups have independently defined the transcriptional profile of several stem cells trying to define the “stemness” by enrichment of genes expressed on several stem cells compared with their mature progeny using Affymetrix microarrays. Compared with differentiated cell types, stem cells expressed a significantly higher number of genes (represented by expressed sequence tags) whose functions are unknown. Murine and human hematopoietic stem cells were found to share a number of expressed gene products, which define key conserved regulatory pathways in this developmental system. This overlapping set of gene products represents a molecular signature of stem cells. The molecular attributes include 1) active JAK/STAT (Janus kinase/signal transducers and activators of transcription), TGF-, Yes (Yamaguchi sarcoma) kinase, and Notch signaling; 2) capacity to sense growth hormone and thrombin; 3) interaction with the extracellular matrix via integrin 6/1, Adam 9, and bystin; 4) engagement in the cell cycle, either arrested in G1 or cycling; 5) high resistance to stress, with upregulated DNA repair, protein folding, ubiquitin system, and detoxifier systems; 6) a remodeled chromatin, acted upon by DNA helicases, DNA methylases, and histone deacetylases; and 7) translation regulated by RNA helicases (120, 223). In embryonic neuroepithelial stem cells (NEP) and progenitors microarrays a few genes appeared to be present exclusively in NEP and not in progenitor cells, e.g., Frizzled 9, Sox-2. The following genes appeared to be enriched in neuronal (e.g., CDK5, neurowww.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 duces the generation of radial glia that become periventricular astrocytes in the adult (78). Notch receptor activation inhibits OPC maturation and myelination in the CNS (282) and may underlie the lack of neurogenic differentiation of progenitors after injury (296). Neuregulins are cytokines present in the ventral ventricular zone where they exert their influence on early OPCs. The neuregulin, glial growth factor-2 (GGF2), is mitogenic for OPCs and is able to maintain OPC plasticity by reverting their phenotype to nestinexpressing precursor cells (39, 40). Neuregulins, as well as their specific receptors ErbB2 and ErbB4, are expressed in multipotent neural precursors from the striatum. They can control the growth of neurospheres. Blocking neuregulin activity in these precursors was associated with decreased mitosis and increased apoptosis. During differentiation of these precursors, neuregulin was required for oligodendrocyte development (32). Vartanian et al. (277) demonstrated that neuregulin is necessary to obtain mature oligodendrocytes from OPCs in the spinal cord. Finally, Sonic hedgehog induces oligodendrocyte development and OPC proliferation from subventricular precursor cells (189). PDGF␣ is involved in generating (225) and is the principal mitogen for OPCs (262). It can also induce chemotaxis of NSCs (73) and inhibit their further terminal differentiation into oligodendrocytes (262). PDGF␣-induced chemotaxis and oligodendrocyte precursor migration of OPCs can be blocked through the chemokine CXCL1 and its CXCR2 receptor (270). Other molecules that affect migration of OPCs are semaphorins and netrins; sema 3A exerts a repulsive effect, whereas netrin-1 has migratory effect, and sema-F appears to have a dual effect (256). During development, bHLH proteins regulate formation of neurons from multipotent progenitor cells. The bHLH protein Id4 stimulates cell proliferation and blocks OPC differentiation, whereas downregulation of this molecule is required in order for OPCs to exit the cell cycle and differentiate (133). The transcription factor genes olig-1 and olig-2 that encode for bHLH proteins are associated with the development of OPCs, and olig-2 is required for oligodendrocyte and motor neuron differentiation in the spinal cord (155) (308). Furthermore, the onset of terminal differentiation of OPC into myelin-forming cells requires the expression of transcription factor Sox10 (257). FGFR3 has also been implicated in this process; in FGFR3⫺/⫺ mice OPCs proliferation is preserved, but terminal differentiation and myelination of OPCs is impaired (202). Hormones and histone deacetylase are important in oligodendroglial development: treatment of rodent and human neurospheres with T3 increases the number of oligodendrocytes observed (190). In contrast, treatment with the histone deacetylase inhibitor trichostatin-A decreases the differentiation of oligodendrocytes resulting in lack of progression of progenitors into mature oligodendrocytes (164)(Table 2). 179 180 INVITED REVIEW pilin, EphB2, FGF11) or glial (e.g., CXCR4, RhoC, CD44, tenascin C) precursors (159). These results open a number of potential discoveries in neural stem biology; the potential of these findings is illustrated by the recent observations that the protein tyrosine phosphatase receptor type Z (PTPRZ), an enriched gene in adult NSCs data set (223), is critical for MS and EAE pathology. Harroch et al. (101), using PTPRZ⫺/⫺ mice, demonstrated that these animals show impaired recovery from EAE and increased apoptosis of mature oligodendrocytes. More importantly in MS, this gene is expressed in remyelinating oligodendrocytes, suggesting a link with remyelinating progenitors cells (101). Endogenous Progenitor Responses During Experimental Injury There is increasing evidence for the existence of regions in the CNS (including in the adult) that can be considered niches for NSCs and pools for different progenitors (206). Recently, it has been shown that, during cerebrogenesis, even exogenous NSCs (used as “reporter” NSCs) segregate into two distinct pools: one that migrates and participates in cerebrogenesis and one that establishes a “dormant” pool of immature cells in secondary germinal zones both surrounding the ventricles and even within the cortical parenchyma (205). These pools may be engaged in repair mechanisms during CNS disease states, for instance. It was demonstrated some time ago that exogenous NSCs in the adult neocortex will give rise solely to glia (the developmentally appropriate response), unless there is a site of neuronal apoptotic death, in which case they will then switch their differentiation fate to yield the missing neurons (252), a program usually played out only during fetal corticogenesis. Furthermore, an incremental response in the form of proliferation and migration of SVZ cells toward lesions was reported (121, 260, 295). Nestin immunoreactivity in the SVZ is increased after injury, and these nestinpositive cells differentiate into astrocytes (105) and possibly into neurons in an experimental model of dopaminergic neurodegeneration (127). Ependymal stem cells migrate out of the central canal and differentiate into astrocytes in response to longitudinal fasciculus injury (75); whereas experimental cortical lesions increase the proliferation and induce expression of PS-NCAM in the SVZ (260). Finally, Magavi et al. (161) found that experimental targeted neuronal apoptosis induced a small degree of de novo cortical neurogenesis, possibly from SVZ precursors, in adult mice CNS. Others found that after injury in the hippocampus, there is increased generation of functional neurons from precursor cells (194) that participate in behavioral learning response (245). Hence, the microenvironment after injury may control the fate of adult stem cells by reinvoking developmental programs and generating new cells that may actively participate in neural circuitry (254). Physiol Genomics • VOL 14 • Several models of experimental demyelination have been used: X-irradiation, chemical induced (lysolecithin, ethidium bromide, cuprizone), viral-induced demyelination, mutant myelin strains and immune mediate demyelination (74, 104). In the study of repair mechanism, the use of a reductionist focused model has the advantage of separating the components of the disease; however, to study progenitors and repair during MS will require the use of a model that closely resemble several aspects of the disease in humans. EAE, an animal model of MS, is characterized by an infiltration in the CNS of autoreactive T cells and macrophages, accompanied by the production of proinflammatory Th1 cytokines (118) and oxidative injury mediators (106). Similar to MS, EAE is an inflammatory, demyelinating disorder in which several pathological and clinical aspects are reproduced, namely, axonal damage, neuronal dysfunction and apoptosis, disruption of the blood-brain barrier (BBB), astrocytosis, prominent inflammatory infiltrate, demyelinating lesions, and remitting relapsing course of the disease (135, 294). These pathological aspects should be investigated in the context of endogenous and exogenous repair and cannot be replicated with more restricted models, such as chemically induced demyelination or with X-irradiation in which there is a limited inflammatory response. In addition, EAE, due to the close correlation of the clinical course of the disease with the subjacent pathology, has served as a screening tool for potential therapies that are now clinically useful (309). In fact NSCs injections have been shown to have an impact in the clinical score of EAE (213). Another beneficial aspect of EAE as a model for studying repair is the existence of several types of induction protocols that have different clinical and pathological severity but yet preserve the inflammatory demyelinating nature of the disease. During the course of EAE, there is increased proliferation of cells in the SVZ, suggesting that stem or progenitor cells in this area may proliferate in response to inflammation/demyelination (34). Picard-Riera et al. (211) extended these observations to a chronic model of EAE, where they demonstrated proliferation of cells from the SVZ, migration to the corpus callosum, and differentiation to astrocytes and a small number of oligodendrocytes. Furthermore, using a model of EAE combined with either injection of MOG antibodies to induce inflammatory demyelination or nonspecific mouse immunoglobulins to induce an inflammatory response without demyelination, Di Bello et al. (58) suggested that remyelination by OPCs appears to be associated more closely with demyelination than with inflammation. However, the greater increase in OPC in inflammatory demyelinating lesions compared with those induced by anti-Gal-C antibodies suggests a mitogenic role for cytokines and growth factors released during inflammation (128). It is possible that the inflammatory environment is an important component of www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 RESPONSES OF PROGENITORS CELLS DURING DEMYELINATION Endogenous Progenitor Responses During Demyelination INVITED REVIEW Exogenous Progenitor Responses To Demyelination The prospect of using transplants of NSCs and progenitors during demyelination requires an understanding of the behavior of these cells in experimental models. This would provide information about the interaction between the damaged host and the NSCs (205, 252). Several groups have used this approach to show that NSCs and OPCs can differentiate into mature oligodendrocytes. For instance, injection of murine NSC in the neonatal SVZ of dysmyelinated shiverer (shi) mice results in widespread oligodendrocyte differentiation with myelination of 40% of the host neuronal process in regions of engraftment with evident clinical improvement (297). EGF-responsive NSCs also have shown efficacy and oligodendrocyte differentiation when transplanted into the thoracolumbar region of md rats (96) and the shaking (Sh) pup canine myelin mutant (96, 179). Nestin-positive human neural precursor cells removed from surgical specimens have been used to induce repair in rat spinal cord with extensive remyelination (4). Even ES cells differentiate into oligodendrocytes and myelinate in culture and after spinal cord transplantation in shi mice (151) and md rat (28). Transplanted OPCs were able to repair focal demyelinated areas in the neonatal and adult canine mutant (8). In the md mutant rat, carrying a mutation in PLP, injection of OPCs results in myelin formation in a wide distribution in the host parenchyma (66), and these cells can be isolated from the rat SVZ as well and produce robust myelin after transplantation (305). Some progenitors isolated from the SVZ require in vitro pre-induction to an oligodendroglial lineage to achieve myelination in vivo (247). Other cells such as olfactory ensheathing cells (OECs) and Schwann cells have been proposed as useful cells for myelin repair. Purified populations of human OECs have extremely Physiol Genomics • VOL 14 • high viability in tissue culture and are capable of remyelinating persistently demyelinated CNS axons and of induction of axonal regeneration (113), following transplantation into experimentally induced demyelinating lesions in the rat spinal cord (11, 126). In addition, Schwann cells have been used for transplantation into demyelinated areas; Schwann cells derived from human sural nerve were transplanted into the X-irradiation/ethidium bromide-lesioned dorsal columns of rats showing extensive remyelination, engraftment, and improved conduction velocity by electrophysiology analysis several weeks postransplantation (11, 113). These results indicate that several types of progenitors and even Schwann cell may be used for therapy and that they receive environmental cues that drive their migration and differentiation toward oligodendrocyte lineage (74). However, the selection of a particular cell type will require extensive experimentation and careful consideration of other factors such as source of isolation, in vitro manipulation, and ethical issues. Finally, the use of immature multipotent progenitors may be more beneficial considering that NSCs cannot only replace neurons and oligodendrocytes, but serve as modulators of the microenvironment by their ability to release growth factors even without transgenic manipulation (213). These results suggest that demyelinating microenvironment may be able to engage exogenous and endogenous progenitor cells to become effectively myelinating cells. However, more information on the behavior the exogenous NSCs and OPCs in a deleterious environment like EAE is needed. MOLECULAR MEDIATORS INVOLVED IN NEURAL PROGENITOR RESPONSES DURING DEMYELINATION Previous studies suggest that, during demyelinating insults induced by gliotoxic agents and in EAE and MS, there is hypertrophy and proliferation of OPCs (50, 58) (Fig. 3, A and B). There is also proliferation of the SVZ where putative NSCs reside, reaching a maximum number at the peak of remyelination, suggesting a response to the local environment (34, 191). Direct evidence of the formation of (remyelinating) oligodendrocytes from OPCs in EAE is lacking (58, 224). However, studies of demyelination induced by the injection of anti-galactocerebroside antibodies and serum complement into the rat spinal cord suggest that remyelination is carried out by OPCs rather than by surviving oligodendrocytes; repeated insults may lead to chronic depletion of the OPC pool in the CNS (26). Therefore the fate and dynamics of NSCs and neural progenitors may be different from those observed in models with a monophasic disease. The chronic nature of human disease (e.g., MS) is likely to influence the dynamics of the pool in a different way: that is, repeated episodes of myelin destruction may eventually result in the formation of chronic demyelinated lesions that fail to remyelinate as observed in MS (50, 289). There are several hypotheses linked to a “nonpermissive” molecular milieu to explain the potential fate of neural precursors www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 the engagement of the endogenous and exogenous progenitors (34, 58). Several models of experimental demyelination have shown that spontaneous remyelination by precursor cells occurs (24). In a lysolecithin-induced demyelination model, progenitor cells of the adult mouse SVZ proliferate, migrate, and differentiate into oligodendrocytes (191, 204). Other groups have confirmed these results with retrovirus labeling of proliferating cells, demonstrating that endogenous progenitors in the subcortical white matter migrate and engage in repair of the lesion (42, 43, 84). Recently, Calza et al. (33), demonstrated that the SVZ cells can be redirected toward oligodendroglial differentiation with the administration of thyroid hormone (T4). T4 administration reduces cell proliferation and nestin expression in the SVZ and upregulates the expression of markers for oligodendrocyte progenitors O4, A2B5, and mature oligodendrocytes in the spinal cord, olfactory bulb, and SVZ. These findings provide a potential molecular target to influence the differentiation of stem cells and oligodendrocyte progenitors into myelinating cells. 181 182 INVITED REVIEW during sustained demyelinating insults: 1) the destruction of OPCs and exhaustion of the OPC or NSC pools by apoptosis; 2) the inability of adult OPCs to fully differentiate into myelinating oligodendrocytes due to an ongoing humoral and cellular immune reaction; 3) The inability of adult OPCs to fully differentiate into myelinating oligodendrocytes due to the lack of appropriate molecular signals or the existence of negative molecular signals for OPC differentiation because of destroyed axons and neurons; and 4) the inhibitory release of molecules during astrogliosis. The following paragraphs outline the experimental evidence and molecular mechanism that may support these hypotheses. Evidence for Precursor Cell Death by Immune Mediators of Demyelination There is evidence that precursor cells might be susceptible to cytokine-induced apoptosis (35, 181) (Fig. 3). For instance, pro-inflammatory cytokines can induce cell death of precursors cells via Fas-FasL interPhysiol Genomics • VOL 14 • actions (62, 233). Oligodendrocytes as well as OPCs are vulnerable to apoptosis by a Fas-mediated mechanism (6, 214). TNF-␣ is upregulated during EAE and MS (219) and can mediate OPC apoptosis in vitro (35). TNF-␣ also potentiates the apoptotic effects of IFN-␥ on OPC cells, and this effect is partially reversible by caspase inhibitors (6). TNF-␣ reduces the expression of transcriptional regulators of survival and differentiation, namely peroxisome proliferator activator receptors (PPARs), especially PPAR␦ (54). The effects of TNF-␣ in vivo may be more complex; mice that lack TNF-␣ or the TNF-␣ receptor-2 (TNFR2) exhibit less remyelination than mice lacking TNF-␣ receptor 1 (TNFR1) in a model of cuprizone-induced demyelination, suggesting a long-term requirement of TNF-␣ in the proliferation of OPCs (9). Thus there may be a dual role for TNF-␣, where an increase in TNF-␣ during the acute phase may be deleterious as observed in experimental brain injury (234). IFN-␥ and lipopolysaccharide induce apoptosis of mature oligodendrocytes and www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 Fig. 3. Changes in OPCs during demyelination and postulated mediators of apoptosis of OPCs. A: a typical NG2 multipolar OPCs is shown in normal spinal cord. B: during the acute phase of experimental demyelination there is an increase in reactivity of NG2 with changes in morphology. The arrows point to OPC processes. C: cytokines and other inflammatory mediators induce cell death of OPCs and other precursor cells by multiple pathways that involve Th1 cytokines, oxidative stress, neurotransmitters, and antibodies (anti-Alu, anti-AN2) that react with specific molecules expressed on OPCs. Surviving precursors can arrest their differentiation through Nmethyl-D-aspartate (NMDA)-mediated signaling. TNF-␣, tumor necrosis factor-␣; IFN-␥, interferon-␥; MHC-I, class I major histocompatibility complex molecule. INVITED REVIEW Inability of Adult OPCs To Fully Differentiate Into Myelinating Oligodendrocytes Data from MS pathology support a role for NSCs and progenitor cells in the pathogenesis of MS (217) (Fig. 4). The analysis of demyelinating lesions in MS during the chronic phase shows that a limited amount of remyelination occurs (229) and appears to be provided by migrating OPCs or NSCs that reside near the lesions. Some authors have shown that MS lesions contain a significant number of quiescent OPCs (286, 287). Using PDGF␣ receptor as a marker, Scolding et al. (236) have identified OPCs in the CNS of normal adults and of MS patients. Others have identified a relatively Physiol Genomics • VOL 14 • quiescent population of OPCs expressing NG2. These OPCs have been observed in lesions of patients with all the clinical subtypes of MS (50). In the chronic stages of the disease, most immature oligodendrocytes did not appear to be engaged in myelination (50), suggesting that oligodendrocyte differentiation of precursor cells is impaired in chronic MS, which is consistent with the general failure of myelin repair observed during the later stages of this disease (289). The lack of remyelination in MS may be due to factors that induce OPC cell death before they can fully mature into myelinating oligodendrocytes, leading to the exhaustion of the progenitor pool. Archelos et al. (7) demonstrated that serum and cerebrospinal fluid (CSF) from 44% of patients with relapsing remitting MS react with protein bearing Alu peptides sequences only found on OPCs, suggesting that there is an ongoing immune response to the OPC in MS. Similarly, CSF from patients with relapsing-remitting active MS, but not from stable MS, contains antibodies against the AN2 molecule, a glycoprotein critical for myelination and expressed only by OPC (195). In vitro experiments confirmed the importance of AN2 in myelination (196). In myelinating cultures, anti-AN2 antibody suppressed the synthesis of myelin protein, supporting the hypothesis that an immunologic attack on AN2-positive OPCs may be responsible for the impaired remyelination capacity (195). Rat OPCs coexpress MOG and NG2 in normal adult spinal cord, raising the possibility that they may be targets of MOGspecific antibodies in this model (146). Lack of Appropriate Molecular Signals from Destroyed Axons and Neurons Normal axons and neurons produce signals that induce oligodendroglial differentiation (31, 156). Neurons, oligodendrocytes and astrocytes not only form an anatomical unit that maintains the nerve transmission, they also maintain a reciprocal paracrine loop between axons, myelin, and oligodendrocyte (174). However, immunologic attacks severely affect the integrity of this unit (174) (Fig. 4). OPCs are influenced by the axonal and neuronal environment through both diffusible and contact-mediated signals such as the neuregulins, PDGF␣ and PDGF, and the FGFs (69, 100, 303). In active and chronic active MS lesions, astrocyte-derived neuregulin is dramatically reduced (279). During the course of MS, there is evidence for neuronal and axonal damage (23, 135, 177, 248), in addition to white matter abnormalities, that can jeopardize the ability for neurons to maintain oligodendrocytes homeostasis; this phenomenon may contribute to the absence of OPC full differentiation (40, 57, 135). Recent work by Trapp and colleagues (49) suggest that the lack of a physical substrate, a normal axon, is an obstacle for terminal differentiation of PLP-positive premyelinating oligodendrocytes. Thus some OPCs may survive the initial nonpermissive milieu and progress to more differentiated cells; however, the lack of a complete normal axon could be a sufficient obstacle for further differentiation. www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 OPC via endogenous nitric oxide production (181), an effect that can be also obtained by cocultures of OPCs with activated microglial cells (207), probably through Toll-like receptor-4 (TLR4) (140). In vivo microglial activation is deleterious to oligodendrocyte progenitors (307), and microgliosis was observed in association with irradiation-induced neurogenesis dysfunction in the hippocampus (183), pointing to the importance of the environment for the fate of neural precursor cells. Infiltrating immune cells are a source of glutamate and can induce damage by excitotoxicity (209, 210). Glutamate receptor AMPA-mediated excitotoxicity is an independent and additional source of oligodendrocyte and axonal damage in EAE (212) and has been shown to induce apoptosis of mature differentiating oligodendrocytes (170, 212). Excitotoxicity may cause damage to other progenitors and NSCs as well, since both multipotent uncommitted and committed progenitors express functional AMPA receptors (81). Furthermore, glutamate agonists induce growth arrest and inhibit oligodendrocyte progenitor differentiation (82). Oligodendrocytes and adult OPCs of adult rats are specifically susceptible to lytic effects of complement (292). However, there are other potential damaging mechanisms for NSC and progenitors during demyelinating injury, such as the expression of molecules that may render these cells immunogenic to the attack of T or B cells; IFN-␥ induces the expression of class I major histocompatibility complex molecule (MHC-I) on neural progenitors, thus making these cells potential targets during an immune response (10, 171). Cytokinestimulated human immature neuroepithelial cell precursors express intercellular adhesion molecule 1 (ICAM-1) and interact with neutrophils by ICAM-1LFA ligation (20). We have demonstrated that murine NSCs in vitro express MHC-I and functional CD80 and CD86 costimulatory molecules, which are upregulated after IFN-␥ stimulation and during stress-induced apoptosis (114). In summary, there is evidence to suggest that OPCs and NSCs may be vulnerable to inflammation, since they express receptors for various mediators of cell death. In vivo studies suggested that in some cases these cells may disappear during acute attacks of MS (50, 288). However, a systematic study of the dynamics of cell proliferation vs. cell death and its role in progression of the disease is still lacking. 183 184 INVITED REVIEW Myelin itself can inhibit the maturation of OPCs in a concentration-dependent manner, suggesting an effect from membrane-associated proteins. Furthermore, axonal regeneration may be inhibited by molecules from injured oligodendrocytes and myelin exerting negative signals. Thus the potential neuronal self-repair and axonal regenerative mechanisms maybe contravened by negative signals released during myelin destruction. A molecule like NOGO-A with known inhibitory effects on axonal regeneration is released after traumatic myelin and oligodendrocytes injury (51, 91, 110). Physiol Genomics • VOL 14 • However, it is not known whether these molecules are released during MS flares nor whether these are responsible for the failure of remyelination in MS. It is likely that several complementary mechanisms contribute to the lack of remyelination in MS (49). Inhibitory Effects of Molecules Released by Reactive Astrocytes The lack of remyelination in MS may result in part from the suppressive effects of gliotic astrocytes on www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 Fig. 4. Effect of axon-myelin unit disturbance and astrogliosis on OPCs. A: scheme depicting a demyelinating lesion. The plaque is composed for two concentric regions the core (blue oval) and the periphery (white oval) with a gradient of pathological alterations; oligodendrocyte loss is represented as black cells with “membrane blebbing,” myelin sheath with different degrees of injury represented by loss of the black color, axonal injury represented by dashed lines, neuronal loss represented by cells with membrane blebbing, and astrocytosis and infiltrating immune cells. This nonpermissive environment perturbs the differentiation of OPCs and blocks axonal regeneration. B: astrogliosis during experimental autoimmune encephalomyelitis (EAE). Astrocytes exhibit thick processes and increase were negative for glial fibrillary acidic protein (GFAP) immunoreactivity (green) compared with control (C). Scale bar ⫽ 20 m. INVITED REVIEW Evidence of Neuroprotective and Trophic Signals for Neural Precursors There is evidence that Th2 cytokines and neurotrophins, in contrast to Th1 cytokines, may be protective Physiol Genomics • VOL 14 • in experimental models of MS and are able to induce the differentiation of OPCs and NSCs in vitro. Certain models of EAE mimic the relapsing-remitting form of MS and are characterized by remission periods that correlate with a shift toward Th2 cytokine production in the CNS, such as TGF- and IL-10 (118). Experimental treatments used in EAE may induce a change toward Th2. For example, CD28-B7 costimulatory blockade results in disease amelioration and inhibits Th1 but spares Th2 cytokines in the CNS (129). In addition to producing cytokines, T cells are able to produce neurotrophins such as NT-3, nerve growth factor (NGF), and BDNF (97, 180). In EAE, there is an increase in NGF immunoreactivity in the SVZ (34). NGF protects the common marmoset against EAE by inducing a Th2 cytokine switch within the CNS by IL-10 upregulation on astrocytes (281). More interestingly, neurotrophins such as NGF are mitogenic for NSCs (47). NT-3 promotes survival, clonal expansion, and proliferation of OPCs (13), and Th2 cytokines favor oligodendroglial development from NSCs (172). The cytokine GGF2 promotes the proliferation, survival, and expansion of oligodendrocytes and enhances remyelination in animals with EAE (162); these animals then evince reduced relapses and increased remyelination, IL-10 expression in the CNS, and upregulation of exon-2 of MBP, indicative of remyelination (36). Furthermore, OPCs have been found to express receptors for GGF2 (38). Additional evidence supporting the idea that Th2 cytokines can interact beneficially with OPCs includes the observation that IL-10 and IL-4 receptors are expressed on OPCs and that these cytokines can protect OPCs from oxidative stress and apoptosis (181). These results suggest a protective role of Th2 and neuropoietic cytokines in the survival and differentiation of OPCs in the CNS during demyelinating injury such as in EAE and MS and suggest that the combination of neurotrophin and Th2 cytokine secretion may improve disease not only by downregulation of autoreactive T cells, but also by improving NSC, OPC, and oligodendroglial survival, leading to replenishment of the oligodendrocyte population. A Theoretical Model of the Role of Neural Precursor Cells in Demyelinating Diseases The heterogenous pathology of MS suggests diversity in the underlying mechanisms causing and/or promoting disease (157). The failure of oligodendrocytes to repair demyelinated axons contributes to the cumulative neurological disability in MS (154). However, it is clear from the experimental and clinical evidence that in the diverse pathological types, OPCs and NSCs are likely to be implicated. A model consistent with alterations in the endogenous potential and survival of these cells is offered in Fig. 5. In normal brain there is a paucity of new oligodendrocytes; OPCs divide infrequently and require signals form neuron and axons for homeostasis (Fig. 5A). The acute lesion of MS may be initiated by an attack from autoreactive T cells, macrophages, and the production www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 myelin formation. EAE and MS lesions are characterized by an increase in astrocyte reactivity demonstrated by hypertrophy and increase in the expression of GFAP but not astrocyte proliferation, despite the fact that MBP peptides are mitogenic in vitro for astrocytes (255). In vivo and in vitro experiments have shown that an astrocytic environment is not permissive for migration of OPCs or for axonal regrowth (68). Reactive astrocytes limit the remyelination capacity of Schwann cells injected into the CNS (25, 241). Furthermore, in MS patients, Schwann cell remyelination is seen in spinal cord lesions in areas that are devoid of astroglial scar (119) (Fig. 4, B and C). In vitro data support these in vivo observations: astrocyte explants were shown to inhibit axonal myelination (117). Interestingly, astrocytes can activate Notch1 receptor on oligodendrocyte precursors via the ligand Jagged-1, which inhibits maturation of these cells. This activation can be mediated by TGF-1, a cytokine that is upregulated in MS. In vivo, Jagged-1 expression was high in hypertrophic astrocytes, and Notch1 was increased on immature oligodendrocytes, suggesting that astrocytes release molecules may have a important regulatory activity on precursor differentiation (123). Although astrocytes may induce a nonpermissive environment by diffusible factors, data from the GFAP⫺/⫺ mice suggest that this intermediate filament gene product (GFAP) expressed by astrocytes is required for long-term maintenance of CNS myelination. When EAE is induced in GFAP⫺/⫺ mice, they exhibit a worsened disease despite remyelination, indicating a distinct functional role of GFAP in astrocytes in control of the disease (148). Astrocytes can modify the migratory properties of OPCs. Fok-Seang et al. (71) studied the ability of cytokine-stimulated astrocytes on OPC migration and neurite outgrowth and found that IFN-␥ exerts an inhibitory effect on OPC migration, whereas IL-1 and bFGF greatly increased axon outgrowth and that this effect could be blocked by TGF-1 and IFN-␥. Interestingly, others have found permissive signals from astrocytes as well (72). Hippocampal astrocytes are uniquely equipped to modulate neurogenesis of NSCs, by instructing the NSCs to adopt a neurogenic fate (253). Taken together these data suggest that cytokine activation of astrocytes may influence both the degree of axonal growth and OPC migration and differentiation; in the case of immune-mediated demyelination, it is likely that astrocyte-derived cytokine production results in a blockade of axonal regeneration and arrest of OPCs differentiation. These cytokines together with other molecular mediators released during demyelinating injury (e.g., tenascin, brevican, neurocan, N-cadherin) (68, 285) may result in a nonpermissive environment for remyelination and axonal regeneration. 185 186 INVITED REVIEW of Th1 cytokines that induce demyelination. OPCs and NSCs may undergo apoptosis through excitotoxicity, oxidative stress, pro-apoptotic cytokines, and antibodymediated cytotoxicity. Some surviving OPCs may increase their numbers, accounting for the initial inPhysiol Genomics • VOL 14 • crease in NG2-positive cells (Fig. 5B). Inactive lesions during the remission phase may show an increase in OPCs with some remyelination (Fig. 5C). However, as the disease progresses during the chronic phase, the fate and the viability of endogenous progenitors may be www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 Fig. 5. Stages of OPC behavior during demyelination. The inflammatory environment may influence the kinetics of endogenous precursor cells during autoimmune-demyelination by altering the balance between apoptosis and proliferation/differentiation of OPCs. A: in normal brain there is a paucity of new oligodendrocytes; OPCs divide infrequently and require signals from neuron and axons for homeostasis. B: an acute multiple sclerosis (MS) lesion may exhibit the same rate of both apoptotic cell death and cellular proliferation with preservation or minimal deterioration of axonal-neuronal signals. C: inactive lesion during the remission phase or a change toward a Th2 environment by immunologic treatment (e.g., with glatiramer acetate) may help to increase proliferation and differentiation of the endogenous pool, this would be protective for the axon as well. D: chronic active lesions and relapses over time may induce four pathological and complementary effects: 1) a loss of the self-renewing capacity of precursor cells as a result of age of the lesion, or the deleterious environment; 2) apoptosis as a direct result of immune factors; 3) lack of trophic support or survival factors form axons and neurons that are failing; 4) direct blockade of differentiation from molecules such as Notch ligands or because of the lack of axons (physical substrate). INVITED REVIEW PROSPECTS FOR THERAPY It remains unclear whether the best strategy for repair will entail promoting remyelination by endogenous progenitors or transplanting in new exogenous progenitors. Much depends on a better understanding not only of the fundamental biological properties of stem/progenitor cells but of the fundamental pathophysiological processes underlying demyelinating/dysmyelinating diseases. If the etiology for disease progression resides in the host genome (e.g., abnormal lifespan, excessive vulnerability to stress, diminished self-renewal capacity), then endogenous progenitors will also be flawed and will not represent a good source for repair (76, 84, 230, 286). If the process is not cell-autonomous, then implanting exogenous cells may be problematic unless they have been engineered to be resistant to the environment that caused the injury. In MS it is likely that both scenarios are implicated, and a combined strategy may be needed. Either way, to exploit the biological properties of precursors and to promote their differentiation in desired directions (e.g., to remyelinating oligodendrocytes), we require a better understanding of the molecular pathways governing Physiol Genomics • VOL 14 • the NSC and OPC pools during development and disease. Cellular therapies may need to be augmented or optimized by adjunctive molecular therapies. For example, it has been demonstrated that the intraventricular injection of TGF-␣ in an animal model of Parkinson’s disease induces a massive proliferation of forebrain cells and their migration to the striatum; this may be accompanied by clinical improvement (67). Cannella et al. (36) showed that the neuregulin GGF2 induced clinical improvement and remyelination in EAE. One of the potential mechanisms of recovery could be mediated by the effects of GGF2 on OPC (36, 38), as OPCs survive and proliferate when exposed to this neuregulin (39, 40, 69, 213). IGF-I is a mitogen for OPCs, and several reports have suggested involvement of IGF-I and PDGF␣ during remyelination in lysolecithin (103) and cuprizone-mediated demyelination models (169). IGF-I is also important for OPC proliferation, and several reports have shown positive effects in gliotoxic models of demyelination and EAE (298). In EAE, clinical amelioration has been reported after IGF-I treatment by some groups (152, 298); others have reported less encouraging results (37). Others investigators showed LIF to have a beneficial effect on oligodendrocyte survival in EAE; whereas in CNTF⫺/⫺ mice EAE severity was increased and OPCs proliferation decreased by 60% and oligodendrocyte apoptosis increased by 50% (150). The continuous local infusion of exogenous cytokines may have positive effects on recruiting endogenous progenitors. In vivo infusion of EGF and FGF results in proliferation and differentiation of progenitors cell (167). In a model of hippocampal injury the infusion of EGF restored the architecture and neurogenesis by recruitment of endogenous progenitors (194). However, it is not clear how long such manipulations are required in the clinical setting to induce an effect; on the other hand a continuous infusion may induce SVZ hyperplasia, raising questions about the tumorogenic potential of such manipulations (136). It is important to remember that trophic factors have pleiotropic effects on the brain: although some may be desirable, others may be undesirable and may, in fact, work at cross-purposes to the other. Indeed, administering the trophic factors via cellular vehicles (such as via neural stem/progenitor cells that have genetically engineered ex vivo) may actually be an optimal delivery system because the factors are provided in a site-specific and in a regulated manner. In addition, because many factors are intrinsically made by such neural-derived cells, potential downregulation of a foreign transgene may be compensated for. Indeed, exogenous NSCs have been successfully isolated (109, 192), genetically manipulated, and transplanted to replace cells and introduce genes in models of neurodegenerative disease in mice (70, 137, 231, 232, 250). Several authors have shown that NSCs survive and differentiate in diverse disease models. NSCs are able to differentiate into the three major cell types in the mammalian CNS; thus they offer promiswww.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 determined by the balance of neuroprotective vs. neurotoxic cytokines, the alteration of neuron-OPC signals, and the changes in self-renewing and differentiation capacity of progenitors. The initial increase in OPC numbers may be lost during the chronic phase. A relapse may induce neurotoxic cytokines in an already altered milieu causing OPC differentiation arrest and apoptosis. Moreover, the gradual alterations in axons and neurons result in a lack of positive signals or the induction of negative signals that arrest the differentiation of surviving OPCs; additionally, the lack of a viable substrate (the axon itself) would be the last event in an irreversible pathway. Repeated attacks over prolonged periods of time may exhaust the pool of cells that have repair ability resulting in complete remyelination failure (Fig. 5D). Therefore, the fate of stem and progenitor cells pools in MS and EAE may depend on the delicate balance between apoptosis-inducing cytokines and proliferation and differentiation-inducing factors (e.g., T cell and neuropoietic cytokines) that are dynamically regulated during inflammatory neurodegeneration or encouraged by immunologic treatments (15, 97, 118). It is likely that the lack of remyelination is mediated by more than one mechanism and that it is a dynamic process that cannot be fully understood with cross-sectional pathology studies. It is possible that the known heterogeneity of NSCs and progenitors extends to heterogeneity in their susceptibility to the milieu; for example, it has been shown that some acute MS lesions do not contain OPCs, suggesting that susceptible cells die during the initial attacks. However, the reappearance of OPCs in the lesion suggests that a subpopulation of cells proliferates, migrates, and perhaps engages in remyelination. 187 188 INVITED REVIEW Physiol Genomics • VOL 14 • NSCs may be a better source of myelinating transplants than OPCs. (230) They seem to possess a greater ability than the more differentiated OPCs for widespread migration toward sites of pathology (1, 297). NSCs may also be easier to use because they can be obtained in great numbers theoretically without the need to predifferentiate them in vitro; allowing environmental cues present at the transplantation site to drive their differentiation (252). Indeed, Yandava et al. (297) found that an environment deficient in functioning oligodendrocytes forced multipotent NSCs to shift their differentiation fate and yield an even greater proportion of that cell type. Whether priming NSCs toward an oligodendrocyte lineage ex vivo would optimize their efficacy remains an important area of investigation. Finally, NSCs may inherently, or after ex vivo manipulation, provide additional neurotrophic factors to repair axonal and neuronal abnormalities present in MS and EAE (3). SUMMARY The goals for MS therapy are as follows: 1) the induction and maintenance of immunologic tolerance toward self-antigens, 2) the promotion of remyelination, 3) the promotion of axonal regeneration, as well as 4) the prevention of further axonal degeneration and neuronal dysfunction as soon as the diagnosis is made. Current therapies deal substantially with the inflammatory and immunologic aspects. This strategy is critical to modulate the “nonpermissive environment” to ensure success when contemplating repair mechanisms. There is a need, however, for a more comprehensive and interdisciplinary analysis of MS molecular pathology. The complex dynamic between the cellular mechanisms responsible for symptoms and for neurodegenerative damage pose important questions that need to be addressed. Regardless of the initial mechanism, disability is cumulative. That is why considering MS as a chronic inflammatory neurodegenerative disease is likely to reflect its true nature and should prompt us to develop a more comprehensive therapy. Although there are now therapies that can prevent exacerbations, we still lack knowledge about how to promote effective remyelination. Adult brain contains adult NSCs and progenitors scattered throughout the CNS in regional pools, and these cells may be able to engage in some degree of endogenous repair. However, the size of the lesion may overcome the capacity for endogenous precursor cells to repair the damage (64, 76). On the other hand, precursor cells could be the targets of the pathological process with adverse repercussions on the innate ability of the CNS to self-repair. Thus any effort in this direction will have to be accompanied by strategies to improve precursors long-term survival (288). Boosting the endogenous pool may be achieved by manipulation of specific molecular signals to induce a preferential differentiation of endogenous NSCs and their derived progenitors toward cells with remyelinating capability. On the other hand, these endogenous www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 ing strategies for transplant as well as gene therapy protocols (137). However, it remains to be determined whether a particular CNS disease will benefit from NSC transplantation (21). The chances for success seem best in those diseases where clinical efficacy is determined ostensibly by a single biological mechanism, e.g., a single gene defect in lysosomal storage diseases. Parkinson’s and Huntington’s disease might fit into that category, i.e., requiring the replacement of a single neuronal cell type in a circumscribed region. MS has always represented a major therapeutic challenge because its pathology is so widespread, affecting multiple sites in the CNS, impairing neuronal as well as glial function. Although targeting localized lesions that cause great disability, such as in the spinal cord, can be envisioned, addressing multiple lesions that extend throughout the CNS remains a daunting prospect, even with extensively migratory stem cells. Careful strategic planning and extensive animal testing will be required before clinical studies can be entertained. Several practical aspects deserve consideration. For instance, the development of methods for noninvasive monitoring of neural stem transplant is required. Magnetic resonance imaging (MRI) tracking of neural progenitors has been developed using the transferrin receptor as an efficient intracellular delivery device for magnetic nanoparticles. Transplanted tagged OPCs injected into the spinal cord of myelin-deficient rats showed a close correlation between the areas of contrast enhancement and the achieved extent of myelination (29, 30). The survival of transplanted NSCs in a potentially toxic environment remains another important consideration. There is some encouraging data on migration and proliferation of transplanted OPCs (266) and multipotent neural progenitors in the EAE model (17). Recently, injection of adult neurospheres forming cells (presumably neural stem/progenitors cells), either intravenously or intracerebroventricularly, resulted in significant numbers of donor cells entering into demyelinating areas of the CNS and differentiating into mature brain cells. Within these areas, oligodendrocyte progenitors markedly increased, with many of them being of donor origin and actively remyelinating axons (213). Replication of these results and more research in relevant models of immune demyelinating disease are needed, especially to investigate the relationship of exogenous repair with the clinicopathological features of MS such as relapses and remissions, axonal damage, sustained oxidative injury, and immune cell infiltrates. For instance, suppression of microglia activation was shown to be beneficial for the survival of transplanted progenitors (307). Furthermore, the implantation procedure itself induces a local injury and opening of the BBB for several weeks (116). It is unknown whether the amount of injury inflicted may trigger a worsening of the local inflammation in an already diseased brain, which could lead to infiltration of immune cells in the region of injection and decreased survival of exogenous progenitors cells (307). INVITED REVIEW We thank Drs. Byron Waksman and Khadir Radassi for critical reading of the manuscript. DISCLOSURES This work has been funded by grants from the National Multiple Sclerosis Society (to S. J. Khoury) and the National Institutes of Health (to S. J. Khoury and E. Y. Snyder). REFERENCES 1. Aboody KS, Brown A, Rainov NG, Bower KA, Liu S, Yang W, Small JE, Herrlinger U, Ourednik V, Black PM, Breakefield XO, and Snyder EY. From the cover: neural stem cells display extensive tropism for pathology in adult brain: evidence from intracranial gliomas. Proc Natl Acad Sci USA 97: 12846–12851, 2000. 2. Ahmed S, Reynolds BA, and Weiss S. BDNF enhances the differentiation but not the survival of CNS stem cell-derived neuronal precursors. J Neurosci 15: 5765–5778, 1995. 3. Akerud P, Canals JM, Snyder EY, and Arenas E. Neuroprotection through delivery of glial cell line-derived neurotrophic factor by neural stem cells in a mouse model of Parkinson’s disease. J Neurosci 21: 8108–8118, 2001. 4. Akiyama Y, Honmou O, Kato T, Uede T, Hashi K, and Kocsis JD. Transplantation of clonal neural precursor cells derived from adult human brain establishes functional peripheral myelin in the rat spinal cord. Exp Neurol 167: 27–39, 2001. 5. Alvarez-Buylla A and Temple S. Stem cells in the developing and adult nervous system. J Neurobiol 36: 105–110, 1998. 6. Andrews T, Zhang P, and Bhat NR. TNF␣ potentiates IFN␥induced cell death in oligodendrocyte progenitors. J Neurosci Res 54: 574–583, 1998. 7. Archelos JJ, Trotter J, Previtali S, Weissbrich B, Toyka KV, and Hartung HP. Isolation and characterization of an oligodendrocyte precursor-derived B-cell epitope in multiple sclerosis. Ann Neurol 43: 15–24, 1998. 8. Archer DR, Cuddon PA, Lipsitz D, and Duncan LD. Myelination of the canine central nervous system by glial cell transplantation: a model for repair of human myelin disease. Nat Med 3: 54–59, 1997. 9. Arnett HA, Mason J, Marino M, Suzuki K, Matsushima GK, and Ting JP. TNF␣ promotes proliferation of oligodendrocyte progenitors and remyelination. Nat Neurosci 4: 1116– 1122, 2001. 10. Bailey KA, Drago J, and Bartlett PF. Neuronal progenitors identified by their inability to express class I histocompatibility antigens in response to interferon-␥. J Neurosci Res 39: 166– 177, 1994. 11. Barnett SC, Alexander CL, Iwashita Y, Gilson JM, Crowther J, Clark L, Dunn LT, Papanastassiou V, Kennedy PG, and Franklin RJ. Identification of a human olfactory ensheathing cell that can effect transplant-mediated remyelination of demyelinated CNS axons. Brain 123: 1581– 1588, 2000. Physiol Genomics • VOL 14 • 12. Barres BA, Burne JF, Holtmann B, Thoenen H, Sendtner M, and Raff MC. Ciliary neurotrophic factor enhances the rate of oligodendrocyte generation. Mol Cell Neurosci 8: 146–156, 1996. 13. Barres BA, Raff MC, Gaese F, Bartke I, Dechant G, and Barde YA. A crucial role for neurotrophin-3 in oligodendrocyte development. Nature 367: 371–375, 1994. 14. Barres BA, Schmid R, Sendnter M, and Raff MC. Multiple extracellular signals are required for long-term oligodendrocyte survival. Development 118: 283–295, 1993. 15. Begolka WS, Vanderlugt CL, Rahbe SM, and Miller SD. Differential expression of inflammatory cytokines parallels progression of central nervous system pathology in two clinically distinct models of multiple sclerosis. J Immunol 161: 4437– 4446, 1998. 16. Belachew S, Aguirre AA, Wang H, Vautier F, Yuan X, Anderson S, Kirby M, and Gallo V. Cyclin-dependent kinase-2 controls oligodendrocyte progenitor cell cycle progression and is downregulated in adult oligodendrocyte progenitors. J Neurosci 22: 8553–8562, 2002. 17. Ben-Hur T, Einstein O, Mizrachi-Kol R, Ben-Menachem O, Reinhartz E, Karussis D, and Abramsky O. Transplanted multipotential neural precursor cells migrate into the inflamed white matter in response to experimental autoimmune encephalomyelitis. Glia 41: 73–80, 2003. 18. Ben-Hur T, Rogister B, Murray K, Rougon G, and DuboisDalcq M. Growth and fate of PSA-NCAM⫹ precursors of the postnatal brain. J Neurosci 18: 5777–5788, 1998. 19. Bergles DE, Roberts JD, Somogyi P, and Jahr CE. Glutamatergic synapses on oligodendrocyte precursor cells in the hippocampus. Nature 405: 187–191, 2000. 20. Birdsall HH. Induction of ICAM-1 on human neural cells and mechanisms of neutrophil-mediated injury. Am J Pathol 139: 1341–1350, 1991. 21. Bjorklund A and Lindvall O. Cell replacement therapies for central nervous system disorders. Nat Neurosci 3: 537–544, 2000. 22. Bjornson CR, Rietze RL, Reynolds BA, Magli MC, and Vescovi AL. Turning brain into blood: a hematopoietic fate adopted by adult neural stem cells in vivo. Science 283: 534– 537, 1999. 23. Black JA, Dib-Hajj S, Baker D, Newcombe J, Cuzner ML, and Waxman SG. Sensory neuron-specific sodium channel SNS is abnormally expressed in the brains of mice with experimental allergic encephalomyelitis and humans with multiple sclerosis. Proc Natl Acad Sci USA 97: 11598–11602, 2000. 24. Blakemore WF. Remyelination of the superior cerebellar peduncle in the mouse following demyelination induced by feeding cuprizone. J Neurol Sci 20: 73–83, 1973. 25. Blakemore WF, Crang AJ, Franklin RJ, Tang K, and Ryder S. Glial cell transplants that are subsequently rejected can be used to influence regeneration of glial cell environments in the CNS. Glia 13: 79–91, 1995. 26. Blakemore WF and Keirstead HS. The origin of remyelinating cells in the central nervous system. J Neuroimmunol 98: 69–76, 1999. 27. Brazelton TR, Rossi FM, Keshet GI, and Blau HM. From marrow to brain: expression of neuronal phenotypes in adult mice. Science 290: 1775–1779, 2000. 28. Brustle O, Jones KN, Learish RD, Karram K, Choudhary K, Wiestler OD, Duncan ID, and McKay RD. Embryonic stem cell-derived glial precursors: a source of myelinating transplants. Science 285: 754–756, 1999. 29. Bulte JW, Douglas T, Witwer B, Zhang SC, Strable E, Lewis BK, Zywicke H, Miller B, van Gelderen P, Moskowitz BM, Duncan ID, and Frank JA. Magnetodendrimers allow endosomal magnetic labeling and in vivo tracking of stem cells. Nat Biotechnol 19: 1141–1147, 2001. 30. Bulte JW, Zhang S, van Gelderen P, Herynek V, Jordan EK, Duncan ID, and Frank JA. Neurotransplantation of magnetically labeled oligodendrocyte progenitors: magnetic resonance tracking of cell migration and myelination. Proc Natl Acad Sci USA 96: 15256–15261, 1999. www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 NSCs may harbor genetic predisposition to malfunction or die or may be excessively vulnerable to the MS milieu. Hence, transplanted exogenous NSCs from a nondiseased source may be required. Such cells may be additionally genetically modified ex vivo prior to transplantation to optimize remyelination and support axonal repair. Regardless of the strategy, more research is needed to understand the signaling pathways for obtaining highly specific molecular targets without inducing aberrant neurogenesis or tumorogenic proliferation. We also must determine exactly how much remyelination is required to redress symptomatic pathology. Clearly, future clinical applications of stem cell biology (including via transplantation) for MS requires a great deal of additional experimental research. 189 190 INVITED REVIEW Physiol Genomics • VOL 14 • 50. Chang A, Nishiyama A, Peterson J, Prineas J, and Trapp BD. NG2-positive oligodendrocyte progenitor cells in adult human brain and multiple sclerosis lesions. J Neurosci 20: 6404– 6412, 2000. 51. Chen MS, Huber AB, van der Haar ME, Frank M, Schnell L, Spillmann AA, Christ F, and Schwab ME. Nogo-A is a myelin-associated neurite outgrowth inhibitor and an antigen for monoclonal antibody IN-1. Nature 403: 434–439, 2000. 52. Chenn A and Walsh CA. Regulation of cerebral cortical size by control of cell cycle exit in neural precursors. Science 297: 365–369, 2002. 53. Chiasson BJ, Tropepe V, Morshead CM, and van der Kooy D. Adult mammalian forebrain ependymal and subependymal cells demonstrate proliferative potential, but only subependymal cells have neural stem cell characteristics. J Neurosci 19: 4462–4471, 1999. 54. Cimini A, Bernardo A, Cifone MG, Di Marzio L, Di Loreto S, Cifone G, and Di Muzio L. TNF␣ downregulates PPAR␦ expression in oligodendrocyte progenitor cells: implications for demyelinating diseases. Glia 41: 3–14, 2003. 55. Clarke DL, Johansson CB, Wilbertz J, Veress B, Nilsson E, Karlstrom H, Lendahl U, and Frisen J. Generalized potential of adult neural stem cells. Science 288: 1660–1663, 2000. 56. Conover JC, Doetsch F, Garcia-Verdugo JM, Gale NW, Yancopoulos GD, and Alvarez-Buylla A. Disruption of Eph/ ephrin signaling affects migration and proliferation in the adult subventricular zone. Nat Neurosci 3: 1091–1097, 2000. 57. De Stefano N, Narayanan S, Francis GS, Arnaoutelis R, Tartaglia MC, Antel JP, Matthews PM, and Arnold DL. Evidence of axonal damage in the early stages of multiple sclerosis and its relevance to disability. Arch Neurol 58: 65–70, 2001. 58. Di Bello IC, Dawson MR, Levine JM, and Reynolds R. Generation of oligodendroglial progenitors in acute inflammatory demyelinating lesions of the rat brain stem is associated with demyelination rather than inflammation. J Neurocytol 28: 365–381, 1999. 59. Doetsch F, Caille I, Lim DA, Garcia-Verdugo JM, and Alvarez-Buylla A. Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell 97: 703–716, 1999. 60. Doetsch F, Garcia-Verdugo JM, and Alvarez-Buylla A. Cellular composition and three-dimensional organization of the subventricular germinal zone in the adult mammalian brain. J Neurosci 17: 5046–5061, 1997. 61. Doetsch F, Petreanu L, Caille I, Garcia-Verdugo JM, and Alvarez-Buylla A. EGF converts transit-amplifying neurogenic precursors in the adult brain into multipotent stem cells. Neuron 36: 1021–1034, 2002. 62. D’Souza SD, Bonetti B, Balasingam V, Cashman NR, Barker PA, Troutt AB, Raine CS, and Antel JP. Multiple sclerosis: Fas signaling in oligodendrocyte cell death. J Exp Med 184: 2361–2370, 1996. 63. Durand B and Raff M. A cell-intrinsic timer that operates during oligodendrocyte development. Bioessays 22: 64–71, 2000. 64. Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, Peterson DA, and Gage FH. Neurogenesis in the adult human hippocampus. Nat Med 4: 1313–1317, 1998. 65. Espinosa de los Monteros A, Zhang M, and De Vellis J. O2A progenitor cells transplanted into the neonatal rat brain develop into oligodendrocytes but not astrocytes. Proc Natl Acad Sci USA 90: 50–54, 1993. 66. Espinosa de los Monteros A, Zhao P, Huang C, Pan T, Chang R, Nazarian R, Espejo D, and de Vellis J. Transplantation of CG4 oligodendrocyte progenitor cells in the myelin-deficient rat brain results in myelination of axons and enhanced oligodendroglial markers. J Neurosci Res 50: 872–887, 1997. 67. Fallon J, Reid S, Kinyamu R, Opole I, Opole R, Baratta J, Korc M, Endo TL, Duong A, Nguyen G, Karkehabadhi M, Twardzik D, and Loughlin S. In vivo induction of massive proliferation, directed migration, and differentiation of neural www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 31. Butt AM and Berry M. Oligodendrocytes and the control of myelination in vivo: new insights from the rat anterior medullary velum. J Neurosci Res 59: 477–488, 2000. 32. Calaora V, Rogister B, Bismuth K, Murray K, Brandt H, Leprince P, Marchionni M, and Dubois-Dalcq M. Neuregulin signaling regulates neural precursor growth and the generation of oligodendrocytes in vitro. J Neurosci 21: 4740– 4751, 2001. 33. Calza L, Fernandez M, Giuliani A, Aloe L, and Giardino L. Thyroid hormone activates oligodendrocyte precursors and increases a myelin-forming protein and NGF content in the spinal cord during experimental allergic encephalomyelitis. Proc Natl Acad Sci USA 99: 3258–3263, 2002. 34. Calza L, Giardino L, Pozza M, Bettelli C, Micera A, and Aloe L. Proliferation and phenotype regulation in the subventricular zone during experimental allergic encephalomyelitis: in vivo evidence of a role for nerve growth factor. Proc Natl Acad Sci USA 95: 3209–3214, 1998. 35. Cammer W. Effects of TNF␣ on immature and mature oligodendrocytes and their progenitors in vitro. Brain Res 864: 213–219, 2000. 36. Cannella B, Hoban CJ, Gao YL, Garcia-Arenas R, Lawson D, Marchionni M, Gwynne D, and Raine CS. The neuregulin, glial growth factor 2, diminishes autoimmune demyelination and enhances remyelination in a chronic relapsing model for multiple sclerosis. Proc Natl Acad Sci USA 95: 10100– 10105, 1998. 37. Cannella B, Pitt D, Capello E, and Raine CS. Insulin-like growth factor-1 fails to enhance central nervous system myelin repair during autoimmune demyelination. Am J Pathol 157: 933–943, 2000. 38. Cannella B, Pitt D, Marchionni M, and Raine CS. Neuregulin and erbB receptor expression in normal and diseased human white matter. J Neuroimmunol 100: 233–242, 1999. 39. Canoll PD, Kraemer R, Teng KK, Marchionni MA, and Salzer JL. GGF/neuregulin induces a phenotypic reversion of oligodendrocytes. Mol Cell Neurosci 13: 79–94, 1999. 40. Canoll PD, Musacchio JM, Hardy R, Reynolds R, Marchionni MA, and Salzer JL. GGF/neuregulin is a neuronal signal that promotes the proliferation and survival and inhibits the differentiation of oligodendrocyte progenitors. Neuron 17: 229–243, 1996. 41. Capela A and Temple S. LeX/ssea-1 is expressed by adult mouse CNS stem cells, identifying them as nonependymal. Neuron 35: 865–875, 2002. 42. Carroll WM and Jennings AR. Early recruitment of oligodendrocyte precursors in CNS demyelination. Brain 117: 563– 578, 1994. 43. Carroll WM, Jennings AR, and Ironside LJ. Identification of the adult resting progenitor cell by autoradiographic tracking of oligodendrocyte precursors in experimental CNS demyelination. Brain 121: 293–302, 1998. 44. Casaccia-Bonnefil P, Hardy RJ, Teng KK, Levine JM, Koff A, and Chao MV. Loss of p27Kip1 function results in increased proliferative capacity of oligodendrocyte progenitors but unaltered timing of differentiation. Development 126: 4027– 4037, 1999. 45. Catanzaro M and Weinert C. Economic status of families living with multiple sclerosis. Int J Rehabil Res 15: 209–218, 1992. 46. Cattaneo E, De Fraja C, Conti L, Reinach B, Bolis L, Govoni S, and Liboi E. Activation of the JAK/STAT pathway leads to proliferation of ST14A central nervous system progenitor cells. J Biol Chem 271: 23374–23379, 1996. 47. Cattaneo E and McKay R. Proliferation and differentiation of neuronal stem cells regulated by nerve growth factor. Nature 347: 762–765, 1990. 48. Chambers CB, Peng Y, Nguyen H, Gaiano N, Fishell G, and Nye JS. Spatiotemporal selectivity of response to Notch1 signals in mammalian forebrain precursors. Development 128: 689–702, 2001. 49. Chang A, Tourtellotte WW, Rudick R, and Trapp BD. Premyelinating oligodendrocytes in chronic lesions of multiple sclerosis. N Engl J Med 346: 165–173, 2002. INVITED REVIEW 68. 69. 70. 71. 73. 74. 75. 76. 77. 78. 79. 80. 81. 82. 83. 84. 85. 86. 87. Physiol Genomics • VOL 14 • 88. Goodell MA, Brose K, Paradis G, Conner AS, and Mulligan RC. Isolation and functional properties of murine hematopoietic stem cells that are replicating in vivo. J Exp Med 183: 1797–1806, 1996. 89. Goodell MA, Rosenzweig M, Kim H, Marks DF, DeMaria M, Paradis G, Grupp SA, Sieff CA, Mulligan RC, and Johnson RP. Dye efflux studies suggest that hematopoietic stem cells expressing low or undetectable levels of CD34 antigen exist in multiple species. Nat Med 3: 1337–1345, 1997. 90. Grandbarbe L, Bouissac J, Rand M, Hrabe De Angelis M, Artavanis-Tsakonas S, and Mohier E. Delta-Notch signaling controls the generation of neurons/glia from neural stem cells in a stepwise process. Development 130: 1391–1402, 2003. 91. GrandPre T, Nakamura F, Vartanian T, and Strittmatter SM. Identification of the Nogo inhibitor of axon regeneration as a Reticulon protein. Nature 403: 439–444, 2000. 92. Gritti A, Bonfanti L, Doetsch F, Caille I, Alvarez-Buylla A, Lim DA, Galli R, Verdugo JM, Herrera DG, and Vescovi AL. Multipotent neural stem cells reside into the rostral extension and olfactory bulb of adult rodents. J Neurosci 22: 437–445, 2002. 93. Gritti A, Frolichsthal-Schoeller P, Galli R, Parati EA, Cova L, Pagano SF, Bjornson CR, and Vescovi AL. Epidermal and fibroblast growth factors behave as mitogenic regulators for a single multipotent stem cell-like population from the subventricular region of the adult mouse forebrain. J Neurosci 19: 3287–3297, 1999. 94. Gross RE, Mehler MF, Mabie PC, Zang Z, Santschi L, and Kessler JA. Bone morphogenetic proteins promote astroglial lineage commitment by mammalian subventricular zone progenitor cells. Neuron 17: 595–606, 1996. 95. Groszer M, Erickson R, Scripture-Adams DD, Lesche R, Trumpp A, Zack JA, Kornblum HI, Liu X, and Wu H. Negative regulation of neural stem/progenitor cell proliferation by the Pten tumor suppressor gene in vivo. Science 294: 2186– 2189, 2001. 96. Hammang JP, Archer DR, and Duncan ID. Myelination following transplantation of EGF-responsive neural stem cells into a myelin-deficient environment. Exp Neurol 147: 84–95, 1997. 97. Hammarberg H, Lidman O, Lundberg C, Eltayeb SY, Gielen AW, Muhallab S, Svenningsson A, Linda H, van Der Meide PH, Cullheim S, Olsson T, and Piehl F. Neuroprotection by encephalomyelitis: rescue of mechanically injured neurons and neurotrophin production by CNS-infiltrating T and natural killer cells. J Neurosci 20: 5283–5291, 2000. 98. Handler M, Yang X, and Shen J. Presenilin-1 regulates neuronal differentiation during neurogenesis. Development 127: 2593–2606, 2000. 99. Hardy R and Reynolds R. Proliferation and differentiation potential of rat forebrain oligodendroglial progenitors both in vitro and in vivo. Development 111: 1061–1080, 1991. 100. Hardy R and Reynolds R. Rat cerebral cortical neurons in primary culture release a mitogen specific for early (GD3⫹/ O4⫺) oligodendroglial progenitors. J Neurosci Res 34: 589–600, 1993. 101. Harroch S, Furtado GC, Brueck W, Rosenbluth J, Lafaille J, Chao M, Buxbaum JD, and Schlessinger J. A critical role for the protein tyrosine phosphatase receptor type Z in functional recovery from demyelinating lesions. Nat Genet 32: 411–414, 2002. 102. Hermanson O, Jepsen K, and Rosenfeld MG. N-CoR controls differentiation of neural stem cells into astrocytes. Nature 419: 934–939, 2002. 103. Hinks GL and Franklin RJ. Delayed changes in growth factor gene expression during slow remyelination in the CNS of aged rats. Mol Cell Neurosci 16: 542–556, 2000. 104. Hiremath MM, Saito Y, Knapp GW, Ting JP, Suzuki K, and Matsushima GK. Microglial/macrophage accumulation during cuprizone-induced demyelination in C57BL/6 mice. J Neuroimmunol 92: 38–49, 1998. 105. Holmin S, Almqvist P, Lendahl U, and Mathiesen T. Adult nestin-expressing subependymal cells differentiate to astrowww.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 72. cells in the adult mammalian brain. Proc Natl Acad Sci USA 97: 14686–14691, 2000. Fawcett JW and Asher RA. The glial scar and central nervous system repair. Brain Res Bull 49: 377–391, 1999. Fernandez PA, Tang DG, Cheng L, Prochiantz A, Mudge AW, and Raff MC. Evidence that axon-derived neuregulin promotes oligodendrocyte survival in the developing rat optic nerve. Neuron 28: 81–90, 2000. Flax JD, Aurora S, Yang C, Simonin C, Wills AM, Billinghurst LL, Jendoubi M, Sidman RL, Wolfe JH, Kim SU, and Snyder EY. Engraftable human neural stem cells respond to developmental cues, replace neurons, and express foreign genes. Nat Biotechnol 16: 1033–1039, 1998. Fok-Seang J, DiProspero NA, Meiners S, Muir E, and Fawcett JW. Cytokine-induced changes in the ability of astrocytes to support migration of oligodendrocyte precursors and axon growth. Eur J Neurosci 10: 2400–2415, 1998. Fok-Seang J, Smith-Thomas LC, Meiners S, Muir E, Du JS, Housden E, Johnson AR, Faissner A, Geller HM, Keynes RJ, and et al. An analysis of astrocytic cell lines with different abilities to promote axon growth. Brain Res 689: 207–223, 1995. Forsberg-Nilsson K, Behar TN, Afrakhte M, Barker JL, and McKay RD. Platelet-derived growth factor induces chemotaxis of neuroepithelial stem cells. J Neurosci Res 53: 521– 530, 1998. Franklin RJ and Blakemore WF. Transplanting oligodendrocyte progenitors into the adult CNS. J Anat 190: 23–33, 1997. Frisen J, Johansson CB, Torok C, Risling M, and Lendahl U. Rapid, widespread, and longlasting induction of nestin contributes to the generation of glial scar tissue after CNS injury. J Cell Biol 131: 453–464, 1995. Gage FH. Mammalian neural stem cells. Science 287: 1433– 1438, 2000. Gage FH, Kempermann G, Palmer TD, Peterson DA, and Ray J. Multipotent progenitor cells in the adult dentate gyrus. J Neurobiol 36: 249–266, 1998. Gaiano N, Nye JS, and Fishell G. Radial glial identity is promoted by Notch1 signaling in the murine forebrain. Neuron 26: 395–404, 2000. Galli R, Borello U, Gritti A, Minasi MG, Bjornson C, Coletta M, Mora M, De Angelis MG, Fiocco R, Cossu G, and Vescovi AL. Skeletal myogenic potential of human and mouse neural stem cells. Nat Neurosci 3: 986–991, 2000. Galli R, Fiocco R, De Filippis L, Muzio L, Gritti A, Mercurio S, Broccoli V, Pellegrini M, Mallamaci A, and Vescovi AL. Emx2 regulates the proliferation of stem cells of the adult mammalian central nervous system. Development 129: 1633–1644, 2002. Gallo V, Pende M, Scherer S, Molne M, and Wright P. Expression and regulation of kainate and AMPA receptors in uncommitted and committed neural progenitors. Neurochem Res 20: 549–560, 1995. Gallo V, Zhou JM, McBain CJ, Wright P, Knutson PL, and Armstrong RC. Oligodendrocyte progenitor cell proliferation and lineage progression are regulated by glutamate receptormediated K⫹ channel block. J Neurosci 16: 2659–2670, 1996. Gard AL and Pfeiffer SE. Two proliferative stages of the oligodendrocyte lineage (A2B5⫹O4⫺ and O4⫹GalC⫺) under different mitogenic control. Neuron 5: 615–625, 1990. Gensert JM and Goldman JE. Endogenous progenitors remyelinate demyelinated axons in the adult CNS. Neuron 19: 197–203, 1997. Geschwind DH, Ou J, Easterday MC, Dougherty JD, Jackson RL, Chen Z, Antoine H, Terskikh A, Weissman IL, Nelson SF, and Kornblum HI. A genetic analysis of neural progenitor differentiation. Neuron 29: 325–339, 2001. Goldman JE, Zerlin M, Newman S, Zhang L, and Gensert J. Fate determination and migration of progenitors in the postnatal mammalian CNS. Dev Neurosci 19: 42–48, 1997. Gomes WA, Mehler MF, and Kessler JA. Transgenic overexpression of BMP4 increases astroglial and decreases oligodendroglial lineage commitment. Dev Biol 255: 164–177, 2003. 191 192 106. 107. 108. 109. 111. 112. 113. 114. 115. 116. 117. 118. 119. 120. 121. 122. 123. cytes in response to brain injury. Eur J Neurosci 9: 65–75, 1997. Hooper DC, Bagasra O, Marini JC, Zborek A, Ohnishi ST, Kean R, Champion JM, Sarker AB, Bobroski L, Farber JL, Akaike T, Maeda H, and Koprowski H. Prevention of experimental allergic encephalomyelitis by targeting nitric oxide and peroxynitrite: implications for the treatment of multiple sclerosis. Proc Natl Acad Sci USA 94: 2528–2533, 1997. Horner PJ, Power AE, Kempermann G, Kuhn HG, Palmer TD, Winkler J, Thal LJ, and Gage FH. Proliferation and differentiation of progenitor cells throughout the intact adult rat spinal cord. J Neurosci 20: 2218–2228, 2000. Hulspas R and Quesenberry PJ. Characterization of neurosphere cell phenotypes by flow cytometry. Cytometry 40: 245– 250, 2000. Hulspas R, Tiarks C, Reilly J, Hsieh CC, Recht L, and Quesenberry PJ. In vitro cell density-dependent clonal growth of EGF-responsive murine neural progenitor cells under serum-free conditions. Exp Neurol 148: 147–156, 1997. Hunter SF, Leavitt JA, and Rodriguez M. Direct observation of myelination in vivo in the mature human central nervous system. A model for the behaviour of oligodendrocyte progenitors and their progeny. Brain 120: 2071–2082, 1997. Hunter-Schaedle KE. Radial glial cell development and transformation are disturbed in reeler forebrain. J Neurobiol 33: 459–472, 1997. Ida JA Jr, Dubois-Dalcq M, and McKinnon RD. Expression of the receptor tyrosine kinase c-kit in oligodendrocyte progenitor cells. J Neurosci Res 36: 596–606, 1993. Imaizumi T, Lankford KL, Burton WV, Fodor WL, and Kocsis JD. Xenotransplantation of transgenic pig olfactory ensheathing cells promotes axonal regeneration in rat spinal cord. Nat Biotechnol 18: 949–953, 2000. Imitola J, Snyder EY, Sayegh MH, and Khoury SJ. Differential upregulation of B7–1 costimulatory molecule during stress induced-apoptosis in neural stem cells (Abstract). Neurology 52: 336, 1999. Imura T, Kornblum HI, and Sofroniew MV. The predominant neural stem cell isolated from postnatal and adult forebrain but not early embryonic forebrain expresses GFAP. J Neurosci 23: 2824–2832, 2003. Isenmann S, Brandner S, Kuhne G, Boner J, and Aguzzi A. Comparative in vivo and pathological analysis of the bloodbrain barrier in mouse telencephalic transplants. Neuropathol Appl Neurobiol 22: 118–128, 1996. Ishikawa M, Tsukamoto T, and Yamamoto T. Long-term cultured astrocytes inhibit myelin formation, but not axonal growth in the co-cultured nerve tissue. Mult Scler 2: 91–95, 1996. Issazadeh S, Navikas V, Schaub M, Sayegh M, and Khoury S. Kinetics of expression of costimulatory molecules and their ligands in murine relapsing experimental autoimmune encephalomyelitis in vivo. J Immunol 161: 1104–1112, 1998. Itoyama Y, Ohnishi A, Tateishi J, Kuroiwa Y, and Webster HD. Spinal cord multiple sclerosis lesions in Japanese patients: Schwann cell remyelination occurs in areas that lack glial fibrillary acidic protein (GFAP). Acta Neuropathol (Berl) 65: 217–223, 1985. Ivanova NB, Dimos JT, Schaniel C, Hackney JA, Moore KA, and Lemischka IR. A stem cell molecular signature. Science 298: 601–604, 2002. Jin K, Minami M, Lan JQ, Mao XO, Batteur S, Simon RP, and Greenberg DA. Neurogenesis in dentate subgranular zone and rostral subventricular zone after focal cerebral ischemia in the rat. Proc Natl Acad Sci USA 98: 4710–4715, 2001. Johansson CB, Momma S, Clarke DL, Risling M, Lendahl U, and Frisen J. Identification of a neural stem cell in the adult mammalian central nervous system. Cell 96: 25–34, 1999. John GR, Shankar SL, Shafit-Zagardo B, Massimi A, Lee SC, Raine CS, and Brosnan CF. Multiple sclerosis: re-expression of a developmental pathway that restricts oligodendrocyte maturation. Nat Med 8: 1115–1121, 2002. Physiol Genomics • VOL 14 • 124. Jones LL, Yamaguchi Y, Stallcup WB, and Tuszynski MH. NG2 is a major chondroitin sulfate proteoglycan produced after spinal cord injury and is expressed by macrophages and oligodendrocyte progenitors. J Neurosci 22: 2792–2803, 2002. 125. Kabos P, Kabosova A, and Neuman T. Blocking HES1 expression initiates GABAergic differentiation and induces the expression of p21 CIP1/WAF1 in human neural stem cells. J Biol Chem 24: 24, 2002. 126. Kato T, Honmou O, Uede T, Hashi K, and Kocsis JD. Transplantation of human olfactory ensheathing cells elicits remyelination of demyelinated rat spinal cord. Glia 30: 209– 218, 2000. 127. Kay JN and Blum M. Differential response of ventral midbrain and striatal progenitor cells to lesions of the nigrostriatal dopaminergic projection. Dev Neurosci 22: 56–67, 2000. 128. Keirstead HS, Levine JM, and Blakemore WF. Response of the oligodendrocyte progenitor cell population (defined by NG2 labelling) to demyelination of the adult spinal cord. Glia 22: 161–170, 1998. 129. Khoury SJ, Akalin E, Chandraker A, Turka LA, Linsley PS, Sayegh MH, and Hancock WW. CD28-B7 costimulatory blockade by CTLA4Ig prevents actively induced experimental autoimmune encephalomyelitis and inhibits Th1 but spares Th2 cytokines in the central nervous system. J Immunol 155: 4521–4524, 1995. 130. Kim HM, Qu T, Kriho V, Lacor P, Smalheiser N, Pappas GD, Guidotti A, Costa E, and Sugaya K. Reelin function in neural stem cell biology. Proc Natl Acad Sci USA 99: 4020– 4025, 2002. 131. Kirn JR, Fishman Y, Sasportas K, Alvarez-Buylla A, and Nottebohm F. Fate of new neurons in adult canary high vocal center during the first 30 days after their formation. J Comp Neurol 411: 487–494, 1999. 132. Kitchens DL, Snyder EY, and Gottlieb DI. FGF and EGF are mitogens for immortalized neural progenitors. J Neurobiol 25: 797–807, 1994. 133. Kondo T and Raff M. The Id4 HLH protein and the timing of oligodendrocyte differentiation. EMBO J 19: 1998–2007, 2000. 134. Kondo T and Raff M. Oligodendrocyte precursor cells reprogrammed to become multipotential CNS stem cells. Science 289: 1754–1757, 2000. 135. Kornek B, Storch MK, Weissert R, Wallstroem E, Stefferl A, Olsson T, Linington C, Schmidbauer M, and Lassmann H. Multiple sclerosis and chronic autoimmune encephalomyelitis: a comparative quantitative study of axonal injury in active, inactive, and remyelinated lesions. Am J Pathol 157: 267–276, 2000. 136. Kuhn HG, Winkler J, Kempermann G, Thal LJ, and Gage FH. Epidermal growth factor and fibroblast growth factor-2 have different effects on neural progenitors in the adult rat brain. J Neurosci 17: 5820–5829, 1997. 137. Lacorazza HD, Flax JD, Snyder EY, and Jendoubi M. Expression of human -hexosaminidase ␣-subunit gene (the gene defect of Tay-Sachs disease) in mouse brains upon engraftment of transduced progenitor cells. Nat Med 2: 424–429, 1996. 138. Lai K, Kaspar BK, Gage FH, and Schaffer DV. Sonic hedgehog regulates adult neural progenitor proliferation in vitro and in vivo. Nat Neurosci 6: 21–27, 2003. 139. Laywell ED, Rakic P, Kukekov VG, Holland EC, and Steindler DA. Identification of a multipotent astrocytic stem cell in the immature and adult mouse brain. Proc Natl Acad Sci USA 97: 13883–13888, 2000. 140. Lehnardt S, Lachance C, Patrizi S, Lefebvre S, Follett PL, Jensen FE, Rosenberg PA, Volpe JJ, and Vartanian T. The toll-like receptor TLR4 is necessary for lipopolysaccharide-induced oligodendrocyte injury in the CNS. J Neurosci 22: 2478–2486, 2002. 141. Lendahl U, Zimmerman LB, and McKay RD. CNS stem cells express a new class of intermediate filament protein. Cell 60: 585–595, 1990. 142. Levine JM, Stincone F, and Lee YS. Development and differentiation of glial precursor cells in the rat cerebellum. Glia 7: 307–321, 1993. www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 110. INVITED REVIEW INVITED REVIEW Physiol Genomics • VOL 14 • 163. 164. 165. 166. 167. 168. 169. 170. 171. 172. 173. 174. 175. 176. 177. 178. 179. Gwynne D, and Raine CS. Neuregulin in neuron/glial interactions in the central nervous system. GGF2 diminishes autoimmune demyelination, promotes oligodendrocyte progenitor expansion, and enhances remyelination. Adv Exp Med Biol 468: 283–295, 1999. Maric D, Maric I, Chang YH, and Barker JL. Prospective cell sorting of embryonic rat neural stem cells and neuronal and glial progenitors reveals selective effects of basic fibroblast growth factor and epidermal growth factor on self-renewal and differentiation. J Neurosci 23: 240–251, 2003. Marin-Husstege M, Muggironi M, Liu A, and CasacciaBonnefil P. Histone deacetylase activity is necessary for oligodendrocyte lineage progression. J Neurosci 22: 10333–10345, 2002. Marmur R, Kessler JA, Zhu G, Gokhan S, and Mehler MF. Differentiation of oligodendroglial progenitors derived from cortical multipotent cells requires extrinsic signals including activation of gp130/LIF receptors. J Neurosci 18: 9800–9811, 1998. Marmur R, Mabie PC, Gokhan S, Song Q, Kessler JA, and Mehler MF. Isolation and developmental characterization of cerebral cortical multipotent progenitors. Dev Biol 204: 577– 591, 1998. Martens DJ, Seaberg RM, and van der Kooy D. In vivo infusions of exogenous growth factors into the fourth ventricle of the adult mouse brain increase the proliferation of neural progenitors around the fourth ventricle and the central canal of the spinal cord. Eur J Neurosci 16: 1045–1057, 2002. Martens DJ, Tropepe V, and van Der Kooy D. Separate proliferation kinetics of fibroblast growth factor-responsive and epidermal growth factor-responsive neural stem cells within the embryonic forebrain germinal zone. J Neurosci 20: 1085– 1095, 2000. Mason JL, Jones JJ, Taniike M, Morell P, Suzuki K, and Matsushima GK. Mature oligodendrocyte apoptosis precedes IGF-1 production and oligodendrocyte progenitor accumulation and differentiation during demyelination/remyelination. J Neurosci Res 61: 251–262, 2000. McDonald JW, Althomsons SP, Hyrc KL, Choi DW, and Goldberg MP. Oligodendrocytes from forebrain are highly vulnerable to AMPA/kainate receptor-mediated excitotoxicity. Nat Med 4: 291–297, 1998. McLaren FH, Svendsen CN, Van der Meide P, and Joly E. Analysis of neural stem cells by flow cytometry: cellular differentiation modifies patterns of MHC expression. J Neuroimmunol 112: 35–46, 2001. Mehler MF, Marmur R, Gross R, Mabie PC, Zang Z, Papavasiliou A, and Kessler JA. Cytokines regulate the cellular phenotype of developing neural lineage species. Int J Dev Neurosci 13: 213–240, 1995. Mehler MF, Rozental R, Dougherty M, Spray DC, and Kessler JA. Cytokine regulation of neuronal differentiation of hippocampal progenitor cells. Nature 362: 62–65, 1993. Merrill JE and Scolding NJ. Mechanisms of damage to myelin and oligodendrocytes and their relevance to disease. Neuropathol Appl Neurobiol 25: 435–458, 1999. Messam CA, Hou J, and Major EO. Coexpression of nestin in neural and glial cells in the developing human CNS defined by a human-specific anti-nestin antibody. Exp Neurol 161: 585– 596, 2000. Messersmith DJ, Murtie JC, Le TQ, Frost EE, and Armstrong RC. Fibroblast growth factor 2 (FGF2) and FGF receptor expression in an experimental demyelinating disease with extensive remyelination. J Neurosci Res 62: 241–256, 2000. Meyer R, Weissert R, Diem R, Storch MK, de Graaf KL, Kramer B, and Bahr M. Acute neuronal apoptosis in a rat model of multiple sclerosis. J Neurosci 21: 6214–6220, 2001. Mezey E, Chandross KJ, Harta G, Maki RA, and McKercher SR. Turning blood into brain: cells bearing neuronal antigens generated in vivo from bone marrow. Science 290: 1779–1782, 2000. Milward EA, Lundberg CG, Ge B, Lipsitz D, Zhao M, and Duncan ID. Isolation and transplantation of multipotential populations of epidermal growth factor-responsive, neural pro- www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 143. Levison SW and Goldman JE. Both oligodendrocytes and astrocytes develop from progenitors in the subventricular zone of postnatal rat forebrain. Neuron 10: 201–212, 1993. 144. Levison SW and Goldman JE. Multipotential and lineage restricted precursors coexist in the mammalian perinatal subventricular zone. J Neurosci Res 48: 83–94, 1997. 145. Levison SW, Young GM, and Goldman JE. Cycling cells in the adult rat neocortex preferentially generate oligodendroglia. J Neurosci Res 57: 435–446, 1999. 146. Li G, Crang AJ, Rundle JL, and Blakemore WF. Oligodendrocyte progenitor cells in the adult rat CNS express myelin oligodendrocyte glycoprotein (MOG). Brain Pathol 12: 463–471, 2002. 147. Lie DC, Dziewczapolski G, Willhoite AR, Kaspar BK, Shults CW, and Gage FH. The adult substantia nigra contains progenitor cells with neurogenic potential. J Neurosci 22: 6639–6649, 2002. 148. Liedtke W, Edelmann W, Chiu FC, Kucherlapati R, and Raine CS. Experimental autoimmune encephalomyelitis in mice lacking glial fibrillary acidic protein is characterized by a more severe clinical course and an infiltrative central nervous system lesion. Am J Pathol 152: 251–259, 1998. 149. Lin RCS, Matesic DF, Marvin M, McKay RD, and Brustle O. Re-expression of the intermediate filament nestin in reactive astrocytes. Neurobiol Dis 2: 79–85, 1995. 150. Linker RA, Maurer M, Gaupp S, Martini R, Holtmann B, Giess R, Rieckmann P, Lassmann H, Toyka KV, Sendtner M, and Gold R. CNTF is a major protective factor in demyelinating CNS disease: a neurotrophic cytokine as modulator in neuroinflammation. Nat Med 8: 620–624, 2002. 151. Liu S, Qu Y, Stewart TJ, Howard MJ, Chakrabortty S, Holekamp TF, and McDonald JW. Embryonic stem cells differentiate into oligodendrocytes and myelinate in culture and after spinal cord transplantation. Proc Natl Acad Sci USA 97: 6126–6131, 2000. 152. Liu X, Yao DL, and Webster H. Insulin-like growth factor I treatment reduces clinical deficits and lesion severity in acute demyelinating experimental autoimmune encephalomyelitis. Mult Scler 1: 2–9, 1995. 153. Lois C and Alvarez-Buylla A. Proliferating subventricular zone cells in the adult mammalian forebrain can differentiate into neurons and glia. Proc Natl Acad Sci USA 90: 2074–2077, 1993. 154. Lovas G, Szilagyi N, Majtenyi K, Palkovits M, and Komoly S. Axonal changes in chronic demyelinated cervical spinal cord plaques. Brain 123: 308–317, 2000. 155. Lu QR, Yuk D, Alberta JA, Zhu Z, Pawlitzky I, Chan J, McMahon AP, Stiles CD, and Rowitch DH. Sonic hedgehogregulated oligodendrocyte lineage genes encoding bHLH proteins in the mammalian central nervous system. Neuron 25: 317–329, 2000. 156. Lubetzki C and Stankoff B. Role of axonal signals in myelination of the central nervous system. . . [in French, English summary]. Pathol Biol (Paris) 48: 63–69, 2000. 157. Lucchinetti C, Bruck W, Parisi J, Scheithauer B, Rodriguez M, and Lassmann H. Heterogeneity of multiple sclerosis lesions: implications for the pathogenesis of demyelination. Ann Neurol 47: 707–717, 2000. 158. Lumelsky N, Blondel O, Laeng P, Velasco I, Ravin R, and McKay R. Differentiation of embryonic stem cells to insulinsecreting structures similar to pancreatic islets. Science 292: 1389–1394, 2001. 159. Luo Y, Cai J, Liu Y, Xue H, Chrest FJ, Wersto RP, and Rao M. Microarray analysis of selected genes in neural stem and progenitor cells. J Neurochem 83: 1481–1497, 2002. 160. Mabie PC, Mehler MF, Marmur R, Papavasiliou A, Song Q, and Kessler JA. Bone morphogenetic proteins induce astroglial differentiation of oligodendroglial-astroglial progenitor cells. J Neurosci 17: 4112–4120, 1997. 161. Magavi SS, Leavitt BR, and Macklis JD. Induction of neurogenesis in the neocortex of adult mice. Nature 405: 951–955, 2000. 162. Marchionni MA, Cannella B, Hoban C, Gao YL, GarciaArenas R, Lawson D, Happel E, Noel F, Tofilon P, 193 194 180. 181. 182. 183. 185. 186. 187. 188. 189. 190. 191. 192. 193. 194. 195. 196. genitor cells from the canine brain. J Neurosci Res 50: 862–871, 1997. Moalem G, Gdalyahu A, Shani Y, Otten U, Lazarovici P, Cohen IR, and Schwartz M. Production of neurotrophins by activated T cells: implications for neuroprotective autoimmunity. J Autoimmun 15: 331–345, 2000. Molina-Holgado E, Vela JM, Arevalo-Martin A, and Guaza C. LPS/IFN-␥ cytotoxicity in oligodendroglial cells: role of nitric oxide and protection by the anti-inflammatory cytokine IL-10. Eur J Neurosci 13: 493–502, 2001. Molne M, Studer L, Tabar V, Ting YT, Eiden MV, and McKay RD. Early cortical precursors do not undergo LIFmediated astrocytic differentiation. J Neurosci Res 59: 301– 311, 2000. Monje ML, Mizumatsu S, Fike JR, and Palmer TD. Irradiation induces neural precursor-cell dysfunction. Nat Med 8: 955–962, 2002. Morrison SJ, Perez SE, Qiao Z, Verdi JM, Hicks C, Weinmaster G, and Anderson DJ. Transient Notch activation initiates an irreversible switch from neurogenesis to gliogenesis by neural crest stem cells. Cell 101: 499–510, 2000. Morrison SJ, White PM, Zock C, and Anderson DJ. Prospective identification, isolation by flow cytometry, and in vivo self-renewal of multipotent mammalian neural crest stem cells. Cell 96: 737–749, 1999. Morshead CM, Benveniste P, Iscove NN, and van der Kooy D. Hematopoietic competence is a rare property of neural stem cells that may depend on genetic and epigenetic alterations. Nat Med 8: 268–273, 2002. Morshead CM, Craig CG, and van der Kooy D. In vivo clonal analyses reveal the properties of endogenous neural stem cell proliferation in the adult mammalian forebrain. Development 125: 2251–2261, 1998. Morshead CM, Reynolds BA, Craig CG, McBurney MW, Staines WA, Morassutti D, Weiss S, and van der Kooy D. Neural stem cells in the adult mammalian forebrain: a relatively quiescent subpopulation of subependymal cells. Neuron 13: 1071–1082, 1994. Murray K, Calaora V, Rottkamp C, Guicherit O, and Dubois-Dalcq M. Sonic hedgehog is a potent inducer of rat oligodendrocyte development from cortical precursors in vitro. Mol Cell Neurosci 19: 320–332, 2002. Murray K and Dubois-Dalcq M. Emergence of oligodendrocytes from human neural spheres. J Neurosci Res 50: 146–156, 1997. Nait-Oumesmar B, Decker L, Lachapelle F, AvellanaAdalid V, Bachelin C, and Van Evercooren AB. Progenitor cells of the adult mouse subventricular zone proliferate, migrate and differentiate into oligodendrocytes after demyelination. Eur J Neurosci 11: 4357–4366, 1999. Nakafuku M and Nakamura S. Establishment and characterization of a multipotential neural cell line that can conditionally generate neurons, astrocytes, and oligodendrocytes in vitro. J Neurosci Res 41: 153–168, 1995. Nakashima K, Yanagisawa M, Arakawa H, Kimura N, Hisatsune T, Kawabata M, Miyazono K, and Taga T. Synergistic signaling in fetal brain by STAT3-Smad1 complex bridged by p300. Science 284: 479–482, 1999. Nakatomi H, Kuriu T, Okabe S, Yamamoto S, Hatano O, Kawahara N, Tamura A, Kirino T, and Nakafuku M. Regeneration of hippocampal pyramidal neurons after ischemic brain injury by recruitment of endogenous neural progenitors. Cell 110: 429–441, 2002. Niehaus A, Shi J, Grzenkowski M, Diers-Fenger M, Hartung HP, Toyka K, Bruck W, and Trotter J. Patients with active relapsing-remitting multiple sclerosis synthesize antibodies recognizing oligodendrocyte progenitor cell surface protein: implications for remyelination. Ann Neurol 48: 362–371, 2000. Niehaus A, Stegmuller J, Diers-Fenger M, and Trotter J. Cell-surface glycoprotein of oligodendrocyte progenitors involved in migration. J Neurosci 19: 4948–4961, 1999. Physiol Genomics • VOL 14 • 197. Nishiyama A, Chang A, and Trapp BD. NG2⫹ glial cells: a novel glial cell population in the adult brain. J Neuropathol Exp Neurol 58: 1113–1124, 1999. 198. Niwa H, Miyazaki J, and Smith AG. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or selfrenewal of ES cells. Nat Genet 24: 372–376, 2000. 199. Noble M, Wren D, and Wolswijk G. The O-2A(adult) progenitor cell: a glial stem cell of the adult central nervous system. Semin Cell Biol 3: 413–422, 1992. 200. Noll E and Miller RH. Oligodendrocyte precursors originate at the ventral ventricular zone dorsal to the ventral midline region in the embryonic rat spinal cord. Development 118: 563–573, 1993. 201. Noseworthy JH, Lucchinetti C, Rodriguez M, and Weinshenker BG. Multiple sclerosis. N Engl J Med 343: 938–952, 2000. 202. Oh LY, Denninger A, Colvin JS, Vyas A, Tole S, Ornitz DM, and Bansal R. Fibroblast growth factor receptor 3 signaling regulates the onset of oligodendrocyte terminal differentiation. J Neurosci 23: 883–894, 2003. 203. Osterhout DJ, Ebner S, Xu J, Ornitz DM, Zazanis GA, and McKinnon RD. Transplanted oligodendrocyte progenitor cells expressing a dominant-negative FGF receptor transgene fail to migrate in vivo. J Neurosci 17: 9122–9132, 1997. 204. Oumesmar BN, Vignais L, Duhamel-Clerin E, AvellanaAdalid V, Rougon G, and Baron-Van Evercooren A. Expression of the highly polysialylated neural cell adhesion molecule during postnatal myelination and following chemically induced demyelination of the adult mouse spinal cord. Eur J Neurosci 7: 480–491, 1995. 205. Ourednik V, Ourednik J, Flax JD, Zawada WM, Hutt C, Yang C, Park KI, Kim SU, Sidman RL, Freed CR, and Snyder EY. Segregation of human neural stem cells in the developing primate forebrain. Science 293: 1820–1824, 2001. 206. Palmer TD, Markakis EA, Willhoite AR, Safar F, and Gage FH. Fibroblast growth factor-2 activates a latent neurogenic program in neural stem cells from diverse regions of the adult CNS. J Neurosci 19: 8487–8497, 1999. 207. Pang Y, Cai Z, and Rhodes PG. Effects of lipopolysaccharide on oligodendrocyte progenitor cells are mediated by astrocytes and microglia. J Neurosci Res 62: 510–520, 2000. 208. Patneau DK, Wright PW, Winters C, Mayer ML, and Gallo V. Glial cells of the oligodendrocyte lineage express both kainate- and AMPA-preferring subtypes of glutamate receptor. Neuron 12: 357–371, 1994. 209. Piani D, Frei K, Do KQ, Cuenod M, and Fontana A. Murine brain macrophages induced NMDA receptor mediated neurotoxicity in vitro by secreting glutamate. Neurosci Lett 133: 159–162, 1991. 210. Piani D, Spranger M, Frei K, Schaffner A, and Fontana A. Macrophage-induced cytotoxicity of N-methyl-D-aspartate receptor positive neurons involves excitatory amino acids rather than reactive oxygen intermediates and cytokines. Eur J Immunol 22: 2429–2436, 1992. 211. Picard-Riera N, Decker L, Delarasse C, Goude K, NaitOumesmar B, Liblau R, Pham-Dinh D, and Evercooren AB. Experimental autoimmune encephalomyelitis mobilizes neural progenitors from the subventricular zone to undergo oligodendrogenesis in adult mice. Proc Natl Acad Sci USA 99: 13211–13216, 2002. 212. Pitt D, Werner P, and Raine CS. Glutamate excitotoxicity in a model of multiple sclerosis. Nat Med 6: 67–70, 2000. 213. Pluchino S, Quattrini A, Brambilla E, Gritti A, Salani G, Dina G, Galli R, Del Carro U, Amadio S, Bergami A, Furlan R, Comi G, Vescovi AL, and Martino G. Injection of adult neurospheres induces recovery in a chronic model of multiple sclerosis. Nature 422: 688–694, 2003. 214. Pouly S, Becher B, Blain M, and Antel JP. Interferon-␥ modulates human oligodendrocyte susceptibility to Fas-mediated apoptosis. J Neuropathol Exp Neurol 59: 280–286, 2000. 215. Pouly S, Prat A, Blain M, Olivier A, and Antel J. NG2 immunoreactivity on human brain endothelial cells. Acta Neuropathol (Berl) 102: 313–320, 2001. www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 184. INVITED REVIEW INVITED REVIEW Physiol Genomics • VOL 14 • 236. Scolding N, Franklin R, Stevens S, Heldin CH, Compston A, and Newcombe J. Oligodendrocyte progenitors are present in the normal adult human CNS and in the lesions of multiple sclerosis. Brain 121: 2221–2228, 1998. 237. Scolding NJ, Frith S, Linington C, Morgan BP, Campbell AK, and Compston DA. Myelin-oligodendrocyte glycoprotein (MOG) is a surface marker of oligodendrocyte maturation. J Neuroimmunol 22: 169–176, 1989. 238. Scolding NJ, Rayner PJ, and Compston DA. Identification of A2B5-positive putative oligodendrocyte progenitor cells and A2B5-positive astrocytes in adult human white matter. Neuroscience 89: 1–4, 1999. 239. Seaberg RM and van der Kooy D. Adult rodent neurogenic regions: the ventricular subependyma contains neural stem cells, but the dentate gyrus contains restricted progenitors. J Neurosci 22: 1784–1793, 2002. 240. Shi J, Marinovich A, and Barres BA. Purification and characterization of adult oligodendrocyte precursor cells from the rat optic nerve. J Neurosci 18: 4627–4636, 1998. 241. Shields SA, Blakemore WF, and Franklin RJ. Schwann cell remyelination is restricted to astrocyte-deficient areas after transplantation into demyelinated adult rat brain. J Neurosci Res 60: 571–578, 2000. 242. Shihabuddin LS, Ray J, and Gage FH. FGF-2 is sufficient to isolate progenitors found in the adult mammalian spinal cord. Exp Neurol 148: 577–586, 1997. 243. Shimazaki T, Shingo T, and Weiss S. The ciliary neurotrophic factor/leukemia inhibitory factor/gp130 receptor complex operates in the maintenance of mammalian forebrain neural stem cells. J Neurosci 21: 7642–7653, 2001. 244. Shingo T, Sorokan ST, Shimazaki T, and Weiss S. Erythropoietin regulates the in vitro and in vivo production of neuronal progenitors by mammalian forebrain neural stem cells. J Neurosci 21: 9733–9743, 2001. 245. Shors TJ, Miesegaes G, Beylin A, Zhao M, Rydel T, and Gould E. Neurogenesis in the adult is involved in the formation of trace memories. Nature 410: 372–376, 2001. 246. Shors TJ, Townsend DA, Zhao M, Kozorovitskiy Y, and Gould E. Neurogenesis may relate to some but not all types of hippocampal-dependent learning. Hippocampus 12: 578–584, 2002. 247. Smith PM and Blakemore WF. Porcine neural progenitors require commitment to the oligodendrocyte lineage prior to transplantation in order to achieve significant remyelination of demyelinated lesions in the adult CNS. Eur J Neurosci 12: 2414–2424, 2000. 248. Smith T, Groom A, Zhu B, and Turski L. Autoimmune encephalomyelitis ameliorated by AMPA antagonists. Nat Med 6: 62–66, 2000. 249. Snyder EY, Deitcher DL, Walsh C, Arnold-Aldea S, Hartwieg EA, and Cepko CL. Multipotent neural cell lines can engraft and participate in development of mouse cerebellum. Cell 68: 33–51, 1992. 250. Snyder EY, Taylor RM, and Wolfe JH. Neural progenitor cell engraftment corrects lysosomal storage throughout the MPS VII mouse brain. Nature 374: 367–370, 1995. 251. Snyder EY and Vescovi AL. The possibilities/perplexities of stem cells. Nat Biotechnol 18: 827–828, 2000. 252. Snyder EY, Yoon C, Flax JD, and Macklis JD. Multipotent neural precursors can differentiate toward replacement of neurons undergoing targeted apoptotic degeneration in adult mouse neocortex. Proc Natl Acad Sci USA 94: 11663–11668, 1997. 253. Song H, Stevens CF, and Gage FH. Astroglia induce neurogenesis from adult neural stem cells. Nature 417: 39–44, 2002. 254. Song HJ, Stevens CF, and Gage FH. Neural stem cells from adult hippocampus develop essential properties of functional CNS neurons. Nat Neurosci 5: 438–445, 2002. 255. South SA, Deibler GE, Tzeng SF, Badache A, Kirchner MG, Muja N, and De Vries GH. Myelin basic protein (MBP) and MBP peptides are mitogens for cultured astrocytes. Glia 29: 81–90, 2000. 256. Spassky N, de Castro F, Le Bras B, Heydon K, QueraudLeSaux F, Bloch-Gallego E, Chedotal A, Zalc B, and www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 216. Power J, Mayer-Proschel M, Smith J, and Noble M. Oligodendrocyte precursor cells from different brain regions express divergent properties consistent with the differing time courses of myelination in these regions. Dev Biol 245: 362–375, 2002. 217. Prineas JW, Barnard RO, Kwon EE, Sharer LR, and Cho ES. Multiple sclerosis: remyelination of nascent lesions. Ann Neurol 33: 137–151, 1993. 218. Pringle NP and Richardson WD. A singularity of PDGF ␣-receptor expression in the dorsoventral axis of the neural tube may define the origin of the oligodendrocyte lineage. Development 117: 525–533, 1993. 219. Probert L, Eugster HP, Akassoglou K, Bauer J, Frei K, Lassmann H, and Fontana A. TNFR1 signalling is critical for the development of demyelination and the limitation of T-cell responses during immune-mediated CNS disease. Brain 123: 2005–2019, 2000. 220. Qian X, Davis AA, Goderie SK, and Temple S. FGF2 concentration regulates the generation of neurons and glia from multipotent cortical stem cells. Neuron 18: 81–93, 1997. 221. Raff MC, Miller RH, and Noble M. A glial progenitor cell that develops in vitro into an astrocyte or an oligodendrocyte depending on culture medium. Nature 303: 390–396, 1983. 222. Rakic SZN. Early oligodendrocyte progenitor cells in the human fetal telencephalon. Glia 41: 117–127, 2003. 223. Ramalho-Santos M, Yoon S, Matsuzaki Y, Mulligan RC, and Melton DA. “Stemness”: transcriptional profiling of embryonic and adult stem cells. Science 298: 597–600, 2002. 224. Redwine JM and Armstrong RC. In vivo proliferation of oligodendrocyte progenitors expressing PDGF␣R during early remyelination. J Neurobiol 37: 413–428, 1998. 225. Redwine JM, Blinder KL, and Armstrong RC. In situ expression of fibroblast growth factor receptors by oligodendrocyte progenitors and oligodendrocytes in adult mouse central nervous system. J Neurosci Res 50: 229–237, 1997. 226. Reynolds R and Hardy R. Oligodendroglial progenitors labeled with the O4 antibody persist in the adult rat cerebral cortex in vivo. J Neurosci Res 47: 455–470, 1997. 227. Reynolds R and Wilkin GP. Development of macroglial cells in rat cerebellum. II. An in situ immunohistochemical study of oligodendroglial lineage from precursor to mature myelinating cell. Development 102: 409–425, 1988. 228. Rietze RL, Valcanis H, Brooker GF, Thomas T, Voss AK, and Bartlett PF. Purification of a pluripotent neural stem cell from the adult mouse brain. Nature 412: 736–739, 2001. 229. Rodriguez M and Scheithauer B. Ultrastructure of multiple sclerosis. Ultrastruct Pathol 18: 3–13, 1994. 230. Rogister B, Ben-Hur T, and Dubois-Dalcq M. From neural stem cells to myelinating oligodendrocytes. Mol Cell Neurosci 14: 287–300, 1999. 231. Rosario CM, Yandava BD, Kosaras B, Zurakowski D, Sidman RL, and Snyder EY. Differentiation of engrafted multipotent neural progenitors towards replacement of missing granule neurons in meander tail cerebellum may help determine the locus of mutant gene action. Development 124: 4213– 4224, 1997. 232. Ryder EF, Snyder EY, and Cepko CL. Establishment and characterization of multipotent neural cell lines using retrovirus vector-mediated oncogene transfer. J Neurobiol 21: 356– 375, 1990. 233. Sabelko-Downes KA, Russell JH, and Cross AH. Role of Fas-FasL interactions in the pathogenesis and regulation of autoimmune demyelinating disease. J Neuroimmunol 100: 42– 52, 1999. 234. Scherbel U, Raghupathi R, Nakamura M, Saatman KE, Trojanowski JQ, Neugebauer E, Marino MW, and McIntosh TK. Differential acute and chronic responses of tumor necrosis factor-deficient mice to experimental brain injury. Proc Natl Acad Sci USA 96: 8721–8726, 1999. 235. Schuldiner M, Yanuka O, Itskovitz-Eldor J, Melton DA, and Benvenisty N. From the cover: effects of eight growth factors on the differentiation of cells derived from human embryonic stem cells. Proc Natl Acad Sci USA 97: 11307–11312, 2000. 195 196 257. 258. 259. 260. 262. 263. 264. 265. 266. 267. 268. 269. 270. 271. 272. 273. 274. Thomas JL. Directional guidance of oligodendroglial migration by class 3 semaphorins and netrin-1. J Neurosci 22: 5992– 6004, 2002. Stolt CC, Rehberg S, Ader M, Lommes P, Riethmacher D, Schachner M, Bartsch U, and Wegner M. Terminal differentiation of myelin-forming oligodendrocytes depends on the transcription factor Sox10. Genes Dev 16: 165–170, 2002. Sun Y, Nadal-Vicens M, Misono S, Lin MZ, Zubiaga A, Hua X, Fan G, and Greenberg ME. Neurogenin promotes neurogenesis and inhibits glial differentiation by independent mechanisms. Cell 104: 365–376, 2001. Suslov ON, Kukekov VG, Ignatova TN, and Steindler DA. Neural stem cell heterogeneity demonstrated by molecular phenotyping of clonal neurospheres. Proc Natl Acad Sci USA 99: 14506–14511, 2002. Szele FG and Chesselet MF. Cortical lesions induce an increase in cell number and PSA-NCAM expression in the subventricular zone of adult rats. J Comp Neurol 368: 439–454, 1996. Tang DG, Tokumoto YM, Apperly JA, Lloyd AC, and Raff MC. Lack of replicative senescence in cultured rat oligodendrocyte precursor cells. Science 291: 868–871, 2001. Tang DG, Tokumoto YM, and Raff MC. Long-term culture of purified postnatal oligodendrocyte precursor cells. Evidence for an intrinsic maturation program that plays out over months. J Cell Biol 148: 971–984, 2000. Taupin P, Ray J, Fischer WH, Suhr ST, Hakansson K, Grubb A, and Gage FH. FGF-2-Responsive neural stem cell proliferation requires CCg, a novel autocrine/paracrine cofactor. Neuron 28: 385–397, 2000. Terada N, Hamazaki T, Oka M, Hoki M, Mastalerz DM, Nakano Y, Meyer EM, Morel L, Petersen BE, and Scott EW. Bone marrow cells adopt the phenotype of other cells by spontaneous cell fusion. Nature 416: 542–545, 2002. Terskikh AV, Easterday MC, Li L, Hood L, Kornblum HI, Geschwind DH, and Weissman IL. From hematopoiesis to neuropoiesis: evidence of overlapping genetic programs. Proc Natl Acad Sci USA 98: 7934–7939, 2001. Tourbah A, Linnington C, Bachelin C, Avellana-Adalid V, Wekerle H, and Baron-Van Evercooren A. Inflammation promotes survival and migration of the CG4 oligodendrocyte progenitors transplanted in the spinal cord of both inflammatory and demyelinated EAE rats. J Neurosci Res 50: 853–861, 1997. Tropepe V, Craig CG, Morshead CM, and van der Kooy D. Transforming growth factor-␣ null and senescent mice show decreased neural progenitor cell proliferation in the forebrain subependyma. J Neurosci 17: 7850–7859, 1997. Tropepe V, Hitoshi S, Sirard C, Mak TW, Rossant J, and van der Kooy D. Direct neural fate specification from embryonic stem cells. A primitive mammalian neural stem cell stage acquired through a default mechanism. Neuron 30: 65–78, 2001. Tropepe V, Sibilia M, Ciruna BG, Rossant J, Wagner EF, and van der Kooy D. Distinct neural stem cells proliferate in response to EGF and FGF in the developing mouse telencephalon. Dev Biol 208: 166–188, 1999. Tsai H, Frost E, To V, Robinson S, Ffrench-Constant C, Geertman R, Ransohoff R, and Miller R. The chemokine receptor CXCR2 controls positioning of oligodendrocyte precursors in developing spinal cord by arresting their migration. Cell 110: 373, 2002. Tsai RY and McKay RD. Cell contact regulates fate choice by cortical stem cells. J Neurosci 20: 3725–3735, 2000. Turnley AM, Faux CH, Rietze RL, Coonan JR, and Bartlett PF. Suppressor of cytokine signaling 2 regulates neuronal differentiation by inhibiting growth hormone signaling. Nat Neurosci 5: 1155–1162, 2002. Uchida N, Buck DW, He D, Reitsma MJ, Masek M, Phan TV, Tsukamoto AS, Gage FH, and Weissman IL. Direct isolation of human central nervous system stem cells. Proc Natl Acad Sci USA 97: 14720–14725, 2000. van Praag H, Christie BR, Sejnowski TJ, and Gage FH. Running enhances neurogenesis, learning, and long-term poPhysiol Genomics • VOL 14 • 275. 276. 277. 278. 279. 280. 281. 282. 283. 284. 285. 286. 287. 288. 289. 290. 291. 292. 293. 294. 295. tentiation in mice. Proc Natl Acad Sci USA 96: 13427–13431, 1999. van Praag H, Kempermann G, and Gage FH. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat Neurosci 2: 266–270, 1999. van Praag H, Schinder AF, Christie BR, Toni N, Palmer TD, and Gage FH. Functional neurogenesis in the adult hippocampus. Nature 415: 1030–1034, 2002. Vartanian T, Fischbach G, and Miller R. Failure of spinal cord oligodendrocyte development in mice lacking neuregulin. Proc Natl Acad Sci USA 96: 731–735, 1999. Vela JM, Molina-Holgado E, Arevalo-Martin A, Almazan G, and Guaza C. Interleukin-1 regulates proliferation and differentiation of oligodendrocyte progenitor cells. Mol Cell Neurosci 20: 489–502, 2002. Viehover A, Miller RH, Park SK, Fischbach G, and Vartanian T. Neuregulin: an oligodendrocyte growth factor absent in active multiple sclerosis lesions. Dev Neurosci 23: 377–386, 2001. Villa A, Snyder EY, Vescovi A, and Martinez-Serrano A. Establishment and properties of a growth factor-dependent, perpetual neural stem cell line from the human CNS. Exp Neurol 161: 67–84, 2000. Villoslada P, Hauser SL, Bartke I, Unger J, Heald N, Rosenberg D, Cheung SW, Mobley WC, Fisher S, and Genain CP. Human nerve growth factor protects common marmosets against autoimmune encephalomyelitis by switching the balance of T helper cell type 1 and 2 cytokines within the central nervous system. J Exp Med 191: 1799–1806, 2000. Wang S, Sdrulla AD, diSibio G, Bush G, Nofziger D, Hicks C, Weinmaster G, and Barres BA. Notch receptor activation inhibits oligodendrocyte differentiation. Neuron 21: 63–75, 1998. Warrington AE, Barbarese E, and Pfeiffer SE. Differential myelinogenic capacity of specific developmental stages of the oligodendrocyte lineage upon transplantation into hypomyelinating hosts. J Neurosci Res 34: 1–13, 1993. Weimann JM, Charlton CA, Brazelton TR, Hackman RC, and Blau HM. Contribution of transplanted bone marrow cells to Purkinje neurons in human adult brains. Proc Natl Acad Sci USA 100: 2088–2093, 2003. Wilby MJ, Muir EM, Fok-Seang J, Gour BJ, Blaschuk OW, and Fawcett JW. N-Cadherin inhibits Schwann cell migration on astrocytes. Mol Cell Neurosci 14: 66–84, 1999. Wolswijk G. Chronic stage multiple sclerosis lesions contain a relatively quiescent population of oligodendrocyte precursor cells. J Neurosci 18: 601–609, 1998. Wolswijk G. Oligodendrocyte precursor cells in chronic multiple sclerosis lesions. Mult Scler 3: 168–169, 1997. Wolswijk G. Oligodendrocyte precursor cells in the demyelinated multiple sclerosis spinal cord. Brain 125: 338–349, 2002. Wolswijk G. Oligodendrocyte survival, loss and birth in lesions of chronic-stage multiple sclerosis. Brain 123: 105–115, 2000. Wolswijk G, Riddle PN, and Noble M. Coexistence of perinatal and adult forms of a glial progenitor cell during development of the rat optic nerve. Development 109: 691–698, 1990. Wren D, Wolswijk G, and Noble M. In vitro analysis of the origin and maintenance of O-2A adult progenitor cells. J Cell Biol 116: 167–176, 1992. Wren DR and Noble M. Oligodendrocytes and oligodendrocyte/type-2 astrocyte progenitor cells of adult rats are specifically susceptible to the lytic effects of complement in absence of antibody. Proc Natl Acad Sci USA 86: 9025–9029, 1989. Wu HY, Dawson MR, Reynolds R, and Hardy RJ. Expression of QKI proteins and MAP1B identifies actively myelinating oligodendrocytes in adult rat brain. Mol Cell Neurosci 17: 292– 302, 2001. Wujek JR, Bjartmar C, Richer E, Ransohoff RM, Yu M, Tuohy VK, and Trapp BD. Axon loss in the spinal cord determines permanent neurological disability in an animal model of multiple sclerosis. J Neuropathol Exp Neurol 61: 23–32, 2002. Yagita Y, Kitagawa K, Ohtsuki T, Takasawa K, Miyata T, Okano H, Hori M, and Matsumoto M. Neurogenesis by www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 261. INVITED REVIEW INVITED REVIEW 296. 297. 298. 300. 301. 302. Physiol Genomics • VOL 14 • 303. 304. 305. 306. 307. 308. 309. 310. source of oligodendrocyte precursors in the ventricular zone of the neural tube. Neuron 12: 1353–1362, 1994. Zajicek J and Compston A. Myelination in vitro of rodent dorsal root ganglia by glial progenitor cells. Brain 117: 1333– 1350, 1994. Zerlin M, Levison SW, and Goldman JE. Early patterns of migration, morphogenesis, and intermediate filament expression of subventricular zone cells in the postnatal rat forebrain. J Neurosci 15: 7238–7249, 1995. Zhang SC, Ge B, and Duncan ID. Adult brain retains the potential to generate oligodendroglial progenitors with extensive myelination capacity. Proc Natl Acad Sci USA 96: 4089– 4094, 1999. Zhang SC, Ge B, and Duncan ID. Tracing human oligodendroglial development in vitro. J Neurosci Res 59: 421–429, 2000. Zhang SC, Goetz BD, and Duncan ID. Suppression of activated microglia promotes survival and function of transplanted oligodendroglial progenitors. Glia 41: 191–198, 2003. Zhou Q and Anderson DJ. The bHLH transcription factors OLIG2 and OLIG1 couple neuronal and glial subtype specification. Cell 109: 61–73, 2002. Ziemssen T, Neuhaus O, Farina C, Hartung HP, and Hohlfeld R. Treatment of multiple sclerosis with glatiramer acetate. Current aspects of mechanisms of action, pharmacokinetics, adverse effect profile and clinical studies. . .[in German, English summary]. Nervenarzt 73: 321–331, 2002. Zulewski H, Abraham EJ, Gerlach MJ, Daniel PB, Moritz W, Muller B, Vallejo M, Thomas MK, and Habener JF. Multipotential nestin-positive stem cells isolated from adult pancreatic islets differentiate ex vivo into pancreatic endocrine, exocrine, and hepatic phenotypes. Diabetes 50: 521–533, 2001. www.physiolgenomics.org Downloaded from http://physiolgenomics.physiology.org/ by 10.220.33.5 on August 3, 2017 299. progenitor cells in the ischemic adult rat hippocampus. Stroke 32: 1890–1896, 2001. Yamamoto S, Nagao M, Sugimori M, Kosako H, Nakatomi H, Yamamoto N, Takebayashi H, Nabeshima Y, Kitamura T, Weinmaster G, Nakamura K, and Nakafuku M. Transcription factor expression and Notch-dependent regulation of neural progenitors in the adult rat spinal cord. J Neurosci 21: 9814–9823, 2001. Yandava BD, Billinghurst LL, and Snyder EY. “Global” cell replacement is feasible via neural stem cell transplantation: evidence from the dysmyelinated shiverer mouse brain. Proc Natl Acad Sci USA 96: 7029–7034, 1999. Yao DL, Liu X, Hudson LD, and Webster HD. Insulin-like growth factor I treatment reduces demyelination and up-regulates gene expression of myelin-related proteins in experimental autoimmune encephalomyelitis. Proc Natl Acad Sci USA 92: 6190–6194, 1995. Ying QL, Nichols J, Evans EP, and Smith AG. Changing potency by spontaneous fusion. Nature 416: 545–548, 2002. Yoon SO, Casaccia-Bonnefil P, Carter B, and Chao MV. Competitive signaling between TrkA and p75 nerve growth factor receptors determines cell survival. J Neurosci 18: 3273– 3281, 1998. Yoshimura K, Sakurai Y, Nishimura D, Tsuruo Y, Nomura M, Kawato S, Seiwa C, Iguchi T, Itoh K, and Asou H. Monoclonal antibody 14F7, which recognizes a stage-specific immature oligodendrocyte surface molecule, inhibits oligodendrocyte differentiation mediated in co-culture with astrocytes. J Neurosci Res 54: 79–96, 1998. Yu WP, Collarini EJ, Pringle NP, and Richardson WD. Embryonic expression of myelin genes: evidence for a focal 197