Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
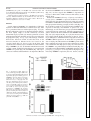
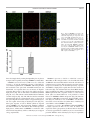
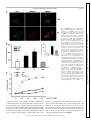
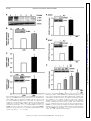
Am J Physiol Endocrinol Metab 306: E1274–E1283, 2014. First published April 15, 2014; doi:10.1152/ajpendo.00007.2014. Endothelial cells respond to hyperglycemia by increasing the LPL transporter GPIHBP1 Amy Pei-Ling Chiu,1 Fulong Wang,1 Nathaniel Lal,1 Ying Wang,1 Dahai Zhang,1 Bahira Hussein,1 Andrea Wan,1 Israel Vlodavsky,2 and Brian Rodrigues1 1 Faculty of Pharmaceutical Sciences, University of British Columbia, Vancouver, British Columbia, Canada; and 2Cancer and Vascular Biology Research Center, Rappaport Faculty of Medicine, Technion, Haifa, Israel Submitted 3 January 2014; accepted in final form 9 April 2014 heparanase; lipoprotein lipase; platelet-derived growth factor; glycosylphosphatidylinositol-anchored high-density lipoprotein binding protein 1 WITH UNINTERRUPTED CONTRACTION being a unique feature of the heart, cardiac muscle has a high demand for energy. This organ demonstrates substrate promiscuity, enabling it to utilize multiple sources of energy such as fatty acids (FA), carbohydrates, amino acids, and ketones (2). Among these, carbohydrate and FA are the major participants from which the heart derives most of its energy. Accordingly, in a basal setting, glucose and lactate account for ⬃30% of energy, whereas 70% of ATP generation is derived from FA oxidation (25). Concerning FA, the heart has a limited capacity to synthesize this substrate and thus relies on 1) release of FA from adipose and transport to the Address for reprint requests and other correspondence: B. Rodrigues, Faculty of Pharmaceutical Sciences, Univ. of British Columbia, 2405 Wesbrook Mall, Vancouver, BC, Canada V6T 1Z3 (e-mail: rodrigue@mail. ubc.ca). E1274 heart after complexing with albumin (24), 2) breakdown of endogenous cardiac triglyceride (TG) (14), and 3) lipolysis of circulating TG-rich lipoproteins to FA by lipoprotein lipase (LPL)positioned at the endothelial cell (EC) surface of the coronary lumen (26). In diabetes, because glucose uptake and oxidation are impaired, the heart is coerced to use FA almost exclusively for ATP generation (33). Multiple adaptive mechanisms, either whole body or intrinsic to the heart, operate to make this achievable. These include augmented adipose tissue lipolysis, where breakdown of stored TG increases circulating FA that are transported to the heart. If delivered to the liver, these FA can raise circulating lipoprotein concentrations, as hepatic FA availability is a rheostat for VLDL synthesis. In so doing, VLDL-TG is an additional and major resource to increase FA delivery to the heart for oxidation (31). Innate to the cardiac muscle, the uptake of albumin-bound FA is driven by plasma membrane FA transporters (for example, CD36), which increase following diabetes (21). Diabetes also enhances adipose TG lipase to mobilize the storage pool of TG within cardiomyocytes (43). Finally, the utilization of VLDL-TG as a FA source by the diabetic heart is influenced not only by elevated plasma VLDL concentrations but also the vascular content of LPL, a rate-limiting enzyme in circulating TG clearance (2). We were the first to report higher luminal LPL activity following diabetes (32). The increase in cardiac LPL with diabetes was immediate and unrelated to gene expression and involved exaggerated processing to dimeric, catalytically active enzymes, an obligatory step for ensuing secretion (39). Transfer to the coronary lumen requires movement of LPL to the cardiomyocyte plasma membrane by AMP-activated protein kinase, protein kinase D, and p38 MAPK (18). Activation of these kinases following diabetes facilitated LPL vesicle formation in addition to promoting cytoskeleton rearrangement for secretion onto surface heparan sulfate proteoglycans (HSPG) (10). For its onward movement across the interstitial space to the basolateral side of vascular ECs, detachment of LPL from the myocyte surface is a prerequisite and is likely mediated by high-glucose-induced secretion of EC heparanase (38). This endoglycosidase, exceptional in its ability to degrade heparan sulfate (HS), instigates LPL release from cardiomyocyte HSPG (38). From here, LPL traverses the interstitial space, and glycosylphosphatidylinositolanchored high-density lipoprotein-binding protein 1 (GPIHBP1) is suggested to deliver it across ECs onto the luminal binding sites of these cells, where the enzyme is functional (42). Thus, GPIHBP1, a protein expressed abundantly in the heart but only on capillary ECs, operates as a transporter, collecting LPL 0193-1849/14 Copyright © 2014 the American Physiological Society http://www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.1 on August 3, 2017 Chiu AP, Wang F, Lal N, Wang Y, Zhang D, Hussein B, Wan A, Vlodavsky I, Rodrigues B. Endothelial cells respond to hyperglycemia by increasing the LPL transporter GPIHBP1. Am J Physiol Endocrinol Metab 306: E1274 –E1283, 2014. First published April 15, 2014; doi:10.1152/ajpendo.00007.2014.—In diabetes, when glucose uptake and oxidation are impaired, the heart is compelled to use fatty acid (FA) almost exclusively for ATP. The vascular content of lipoprotein lipase (LPL), the rate-limiting enzyme that determines circulating triglyceride clearance, is largely responsible for this FA delivery and increases following diabetes. Glycosylphosphatidylinositol-anchored high-density lipoprotein-binding protein [GPIHBP1; a protein expressed abundantly in the heart in endothelial cells (EC)] collects LPL from the interstitial space and transfers it across ECs onto the luminal binding sites of these cells, where the enzyme is functional. We tested whether ECs respond to hyperglycemia by increasing GPIHBP1. Streptozotocin diabetes increased cardiac LPL activity and GPIHBP1 gene and protein expression. The increased LPL and GPIHBP1 were located at the capillary lumen. In vitro, passaging EC caused a loss of GPIHBP1, which could be induced on exposure to increasing concentrations of glucose. The high-glucoseinduced GPIHBP1 increased LPL shuttling across EC monolayers. GPIHBP1 expression was linked to the EC content of heparanase. Moreover, active heparanase increased GPIHBP1 gene and protein expression. Both ECs and myocyte heparan sulfate proteoglycanbound platelet-derived growth factor (PDGF) released by heparanase caused augmentation of GPIHBP1. Overall, our data suggest that this protein “ensemble” (heparanase-PDGF-GPIHBP1) cooperates in the diabetic heart to regulate FA delivery and utilization by the cardiomyocytes. Interrupting this axis may be a novel therapeutic strategy to restore metabolic equilibrium, curb lipotoxicity, and help prevent or delay heart dysfunction that is characteristic of diabetes. HIGH-GLUCOSE REGULATION OF GPIHBP1 from the interstitial space and transferring it across ECs (7, 9, 15). Given that GPIHBP1 mRNA levels change rapidly (22), we tested whether this protein has a part to play in the transfer of LPL to the vascular lumen after diabetes. Our data suggest a novel mechanism in which high-glucose-induced secretion of heparanase not only liberates LPL from the myocyte cell surface but can also induce GPIHBP1 expression, thus promoting LPL shuttling to the apical side of ECs. MATERIALS AND METHODS nase, protein was concentrated with an Amicon centrifuge filter (Millipore), followed by the procedures listed above. RNA isolation and real-time PCR. Total RNA was isolated from rat hearts and RAOECs using Trizol, followed by chloroform and isopropanol extraction, washing by ethanol, and dissolving in DEPCH2O. RNA was reverse transcribed into cDNA with a mixture of random primers, oligo-dT, and SuperScript III. cDNA were amplified by TaqMan probes (GPIHBP1 and -actin) in triplicate, using a 7900HT Fast Real-Time PCR system. Gene expression was calculated with the comparative cycle threshold method. EC culture. RAOECs (Cell Applications) and bovine coronary artery ECs (BCAECs; Clonetics) were cultured at 37°C in a 5% CO2 humidified incubator to 80 –90% confluence (38). Cells were detached following trypsin digestion, and aliquots were used for seeding of a new culture. Cells from passages 5–10 were used for detection of GPIHBP1 and heparanase. EC and cardiomyocyte coculture. Only RAOECs and BCAECs from passages 8 –10 were used for this experiment. ECs were seeded on Transwell inserts (24-mm diameter, 1.0-m pore size; Falcon). On reaching 80 –90% confluence, these inserts were placed in a six-well plate that had cardiomyocytes attached to the bottom. Cardiomyocytes were plated at a density of 200,000 cells/well (40). LPL transport assay. To study basolateral to apical transport of LPL, RAOECs were seeded on Transwell inserts until the cells formed a tight monolayer. The inserts were then placed in six-well plates. Subsequently, LPL (5,000 U/10 g/ml) was added to the bottom chamber. After 1 h, LPL that translocated to the apical surface of ECs was collected by heparin displacement and LPL activity determined. In vitro lipolysis of VLDL. Isolation of VLDL (without chylomicrons) by density gradient ultracentrifugation was achieved using serum from 16-h-fasted rats (6 PM to 10 AM). Fasting of rats for this duration led to the production of more VLDL and minimized the contribution from chylomicrons. In vitro lipolysis of VLDL-TG was carried out on the basis of previously described methods (36). Briefly, various concentrations (0 – 0.8 mM) of VLDL-TG were incubated with heparin-releasable LPL at 37°C for 30 min. At the end of incubation the reaction was stopped by the addition of precooled 0.3 M Na2PO4 (pH 6.9), and the tubes were immediately immersed in ice. From this reaction mixture, 50 l was pipetted in triplicate to measure free fatty acids released by hydrolysis of VLDL-TG with a NEFA C kit (Wako Chemicals). Cytokine array. Media were collected from RAOECs (EC culture medium) incubated with DMEM containing either 5.5 or 25 mM glucose for 12 h. Myocytes (myocyte culture medium) incubated with DMEM or DMEM containing 500 ng/ml purified heparanase (HPA) were also collected. Media were centrifuged at 2,000 rpm for 5 min. Array membranes were incubated with blocking buffer at room temperature (RT), followed by a 1-h incubation with EC culture medium or myocyte culture medium at RT for 2 h. Membranes were washed (3 times for 5 min) and incubated with working primary antibody and anti-horseradish peroxidase, respectively, at RT for 2 h. Array membrane was detected with enhanced chemiluminesence (ECL) and Hyperfilm ECL Film (GE Healthcare). Reagents and antibodies. Heparin (Hepalean, 1,000 U/ml) was from Organon. [3H]triolein was purchased from Amersham Canada. Anti-HPA antibody monoclonal antibody 130 was from InSight (Rehovot, Israel). Anti-GPIHBP1 antibody was from Novus Biologicals. Anti-PECAM (CD31) was from Chemicon (Millipore). Anti--actin (C4), goat anti-mouse antibody, and goat anti-rabbit antibody were from Santa Cruz Biotechnology. To measure free fatty acid released from VLDL-TG breakdown, a NEFA C assay kit was purchased from Wako. Purified active and latent heparanase were a kind gift from Dr. Israel Vlodavsky, Hadassah University Hospital, Jerusalem, Israel. Phosphatidylinositol-specific phospholipase C (PI-PLC), Alexa 555-labeled anti-mouse IgG, Alexa 488-labeled anti-rabbit IgG, Trizol, SuperScript III RT, GPIHBP1 (Rn01503971_g1) and actin AJP-Endocrinol Metab • doi:10.1152/ajpendo.00007.2014 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.1 on August 3, 2017 Experimental animals. This investigation adhered to the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health and was approved by the Animal Care Committee of the University of British Columbia (certificate no. A13-0098). Adult male Wistar rats (250 –320 g) were injected with streptozotocin (STZ), a -cell-specific toxin. STZ (55 mg/kg) was administered intravenously as a single dose to generate a model of poorly controlled type 1 diabetes (19, 20). Hyperglycemia was confirmed in blood samples from the tail vein using a glucometer (AccuSoft) and glucose test strips (Accu-Chek Advantage; Roche). STZ animals were kept for 4 days before hearts were removed for both LPL and GPIHBP1 measurement. Heart perfusion and isolation of cardiomyocytes. Rats were anesthetized with 65 mg/kg ip pentobarbital sodium, the thoracic cavities were opened, and hearts were carefully excised. After cannulation of the aorta, the heart was secured by tying below the innominate artery and perfused retrogradely with Krebs-Henseleit buffer (37). Ventricular calcium-tolerant myocytes were prepared by way of a previously described procedure (34). Briefly, myocytes were made calcium tolerant by successive exposure to increasing concentrations of calcium. Cardiomyocytes were plated at a density of 200,000 cells/well on laminin-coated six-well culture plates. Cells were maintained using Medium-199 and incubated at 37°C in a 5% CO2 humidified incubator. LPL activity. To measure coronary LPL, Krebs buffer containing heparin (8 U/ml) was used, perfusion effluent was collected over 5 min, and LPL activity was determined. To release EC surface-bound LPL, cells were incubated with heparin (8 U/ml) for 3 min. LPL activity was determined by measuring in vitro hydrolysis of [3H]triolein substrate (18). Radiolabeled FA products were extracted and estimated by liquid scintillation counting. Immunofluorescence. To visualize LPL and GPIHBP1 localization, rat hearts were perfused with cold PBS and fixed in 10% paraformaldehyde overnight, and 8- to 10-m parafilm-embedded sections were prepared. Slides were incubated in a 56°C oven for 30 min and dehydrated by xylene (twice for 10 min) and ethanol (35–95%, 5 min) followed by PBS. Sections were permeabilized with 0.2% Triton X-100 in PBS for 30 min and incubated with blocking buffer containing 10% donkey serum and 0.2% Triton X-100 in PBS for 30 min at room temperature. Slides were then incubated with primary antibodies at 4°C overnight: 1:100 for anti-CD31, anti-GPIHBP1 (8), and anti-LPL. Secondary antibodies were used at a dilution of 1:200 and incubated at room temperature for 1 h. For immunofluorescence detection of proteins in rat aortic endothelial cells(RAOECs), cells were seeded on slides and grown until they were 80 –90% confluent. Following treatment with or without high glucose (25 mM), cells were washed with cold PBS and fixed with methanol at 4°C for 1 h, and the above procedures (permeabilization, blocking, primary/secondary antibodies) were repeated. Western blot. To measure protein expression, 30 g of protein was loaded. Membranes were incubated with primary antibodies (1:1,000 for anti-GPIHBP1, 1:2,000 for anti--actin, and 1:500 for anti-heparanase) at 4°C overnight and subsequently with secondary antibodies (1:1,000) at room temperature for 1 h. To measure medium hepara- E1275 E1276 HIGH-GLUCOSE REGULATION OF GPIHBP1 (Rn00667869_m1) probe, and TaqMan fast advanced master mix were obtained from Invitrogen. Protein array kit (Rat Cytokine Array C2) was purchased from RayBiotech. Statistical analysis. Values are means ⫾ SE. Wherever appropriate, one-way ANOVA followed by the Bonferroni test was used to determine differences between group mean values. The level of statistical significance was set at P ⬍ 0.05. RESULTS Fig. 1. Streptozotocin (STZ) diabetes increases cardiac lipoprotein lipase (LPL) activity and glycosylphosphatidylinositol-anchored high-density lipoprotein-binding protein 1 (GPIHBP1) expression. Rats were made hyperglycemic by iv injection of 55 mg/kg STZ. After 4 days of hyperglycemia, animals were euthanized; hearts were removed and either perfused with heparin (8 U/ml) to release coronary LPL activity (A, left and top right) or used for immunofluorescent detection of LPL (A, bottom right) and protein measurement of GPIHBP1 (B). Coronary LPL activity was determined at the indicated times by measuring the in vitro hydrolysis of [3H]triolein substrate. LPL immunofluorescence was detected using an anti-rabbit LPL antibody. Scale bar, 20 m. *P ⬍ 0.05 compared with control (CON); n ⫽ 3. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00007.2014 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.1 on August 3, 2017 Cardiac LPL and GPIHBP1 after STZ diabetes. Following injection of STZ and maintenance of animals for 4 days, there was a pronounced increase in blood glucose (Fig. 1A, right). Because utilization of glucose is impaired following diabetes, the heart rapidly adapts to use FA exclusively, an effect determined largely by LPL (18). Indeed, perfusion of hearts with heparin indicated that LPL activity increased following hyperglycemia (Fig. 1A, top right). Interestingly, immunofluorescence detection of LPL revealed that most of the increased enzyme in diabetic hearts was located at the coronary lumen (Fig. 1A, bottom right). In the heart, LPL synthesized in cardiomyocytes is shuttled across ECs with the help of GPIHBP1 (12, 42). To determine whether the increased vascular content of LPL following diabetes is associated with GPIHBP1, its protein (Fig. 1B) and mRNA (Fig. 2B) expression were examined and determined to be augmented. Because the increased GPIHBP1 had an exclusive endothelial location (Fig. 2A), our data suggest that GPIHBP1 is important for myocyte to EC transfer of LPL to increase FA delivery to the diabetic heart. Endothelial GPIHBP1 following cell passage and influence of high glucose. GPIHBP1 is expressed exclusively in endothelial cells (42). We determined the level of this glycoprotein in RAOECs and BCAECs and unexpectedly identified loss of this glycoprotein following passaging of these cells (Fig. 3A). In an attempt to mimic in vivo hyperglycemia, RAOECs were exposed to increasing concentrations of glucose for 12 h. Interestingly, in cells from later passages (in which GPIHBP1 expression had been suppressed), although concentrations of glucose between 10 and 15 mM increased GPIHBP1 mRNA (Fig. 3B, right) and protein (Fig. 3C, right), the increases were more robust with the higher concentration of glucose. Choosing 25 mM glucose and different times, high glucose increased GPIHBP1 mRNA expression, an effect that was pronounced after 12 h (Fig. 3B). Measurement of GPIHBP1 protein under these conditions uncovered an even more rapid increase within 4 h of incubation with high glucose (Fig. 3C). This increased EC GPIHBP1 was predominantly membrane bound (Fig. 4A). It should be noted that RAOECs and BCAECs from the earlier passages (e.g., passages 5–7) also responded to high glucose by increasing GPIHBP1 expression (data not shown). Overall, E1277 HIGH-GLUCOSE REGULATION OF GPIHBP1 these data imply that by sensing the impending loss of glucose transport, ECs respond by increasing GPIHBP1 to shuttle LPL. GPIHBP1-associated transfer of LPL across the endothelial monolayer. To test the functional relevance of the high-glucose-induced augmentation of GPIHBP1, LPL transport from the basolateral to the apical side of RAOEC monolayers was determined. As expected, there was an increase in heparinreleasable LPL activity at the apical side of ECs exposed to high glucose (Fig. 4B), an effect that was blunted by PI-PLC (Fig. 4B, inset). The osmotic control mannitol had no influence on LPL transport (Fig. 4B). The LPL that had been shuttled following high glucose was functionally active, as exposure of this apical heparin-releasable enzyme to lipoprotein triglyceride was capable of increasing its hydrolysis to FA (Fig. 4C). Since prior exposure of cells to PI-PLC [which is expected cleave GPIHBP1 (15) and prevent LPL shuttling] abolished this TG breakdown, our data indicate that in ECs exposed to high glucose, enhancement of GPIHBP1 is an effective stimulus for increasing delivery of FA to cardiomyocytes to support energy requirements. GPIHBP1 expression is linked to endothelial content of HPA. HPA, an EC endoglycosidase, can cleave HS side chains on HSPGs in the extracellular matrix and on the cell surface of cardiomyocytes to release LPL for transfer across the interstitial space to reach ECs (29). As described previously, exposure of RAOECs to high glucose rapidly increased the secretion of both active (Fig. 5A) and latent (data not shown) HPA. Intriguingly, in addition to its ability to rapidly liberate cardiomyocyte HSPG-sequestered LPL, exposure of ECs to active but not latent (data not shown) HPA for 12 h produced a robust increase in GPIHBP1 mRNA and protein (Fig. 5B). Because passaging of RAOECs progressively decreased HPA expression (Fig. 5C) similarly to loss of GPIHBP1, our data have uncovered a novel autocrine role (either direct or indirect) for active HPA to affect GPIHBP1 expression. Cardiomyocytes induce EC GPIHBP1. Although GPIHBP1 is highly expressed in ECs in vivo, ECs in culture lose expression of this glycoprotein (1). Because ECs in the heart are closely appositioned to cardiomyocytes, we reasoned that a paracrine influence from cardiomyocytes may be responsible AJP-Endocrinol Metab • doi:10.1152/ajpendo.00007.2014 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.1 on August 3, 2017 Fig. 2. Increased GPIHBP1 is located at the capillary lumen in diabetic hearts and is associated with augmented gene expression. After 4 days, both CON and STZ hearts were collected for immunofluorescence microscopy and mRNA measurement. Paraffin-embedded sections were used and permeabilized with 0.2% Triton X in PBS. A: primary anti-mouse CD31 and anti-rabbit GPIHBP1 antibodies were used to stain the capillary lumen. Scale bar, 20 m. B: GPIHBP1 mRNA level was determined using TaqMan (ABI) following RNA isolation and reverse transcription. *P ⬍ 0.05 compared with CON; n ⫽ 3. E1278 HIGH-GLUCOSE REGULATION OF GPIHBP1 for EC expression of GPIHBP1. Using a coculture system to mimic the intact heart, we determined that in ECs in which GPIHBP1 had been silenced (passages 8 –10), a simple introduction of cardiomyocytes into the vicinity restored GPIHBP1 expression to levels observed with early cell passages (Fig. 6, A and B). These results point toward paracrine factors from cardiomyocytes influencing EC GPIHBP1 expression. Assuming that the stimulus for the release of signaling mediators from myocytes could originate from ECs (likely HPA), we exposed the coculture to high glucose. Noteworthy, a further expression of EC GPIHBP1 was evident in this condition (Fig. 6C), suggesting that high-glucose-induced HPA released from ECs can affect GPIHBP1 by both autocrine and paracrine signaling. HPA-released platelet-derived growth factor from myocytes and ECs can induce GPIHBP1 expression. ECs and myocyte HSPG-anchored proteins can be liberated by HPA (38). Using a protein array and ECs exposed to high glucose or myocytes treated with active HPA, a number of proteins were increased in the culture medium. These included monocyte chemotactic protein-1, prolactin, receptor for advanced glycation end products, thymidylate kinase TMP-1 (data not shown), and platelet-derived growth factor (PDGF) (Fig. 7A). Given the acknowledged role of PDGF in FA metabolism (35), we treated EC with recombinant PDGF and discovered an induction of GPIHBP1 from passage 7 (which still express GPIHBP1) and passage 10 (in which there is a loss of GPIHBP1) (Fig. 7B). Nevertheless, the increase was more robust in cells from passage 10, as these cells had an extremely low initial expression of GPIHBP1. Our data suggest a mediator role (either autocrine or paracrine) for this growth factor in FA delivery to the diabetic myocyte. DISCUSSION Underutilization of glucose and overreliance on FA are hallmarks of the diabetic heart (33). Although albumin-bound circulating FA is AJP-Endocrinol Metab • doi:10.1152/ajpendo.00007.2014 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.1 on August 3, 2017 Fig. 3. Passaging of endothelial cells causes loss of GPIHBP1, which can be induced on exposure to high glucose. Rat aortic endothelial cells (RAOECs) and bovine coronary artery endothelial cells (BCAECs) were cultured to 80–90% confluence. A: cells were detached following trypsin digestion, and aliquots were used either for detection of GPIHBP1 or seeding of a new culture. B and C: RAOECs from passages 7–10 were treated with increasing concentrations of glucose (B and C, right) for different times, and GPIHBP1 mRNA (B) and protein (C) were determined using TaqMan and Western blotting, respectively. *P ⬍ 0.05 compared with 0 h or 5.5 mM glucose; n ⫽ 3–4. E1279 HIGH-GLUCOSE REGULATION OF GPIHBP1 an important source of this substrate, its molar concentration is ⬃10-fold less than that of lipoprotein TG (26). As such, the hydrolysis of circulating TG is suggested to be the predominant source of FA for cardiac utilization during diabetes (3). Lipoprotein-TG breakdown occurs at the coronary lumen, with the assistance of LPL positioned at apical surface of ECs. Following diabetes, in the absence of changes in LPL synthesis, coronary LPL activity is aug- mented by posttranslational modifications. These include an increase in LPL dimerization (39), vesicle transport (18), and shuttling of the enzyme from the myocyte surface to the basolateral side of ECs (6). Data from the current study present the novel idea that in response to hyperglycemia, transport of LPL to the apical side of ECs to assist in TG breakdown is achievable through induction of GPIHBP1. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00007.2014 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.1 on August 3, 2017 Fig. 4. GPIHBP1 induced by high glucose increases LPL shuttling across endothelial cell monolayers. A: RAOECs seeded on slides were placed in a 6-well plate and treated with 5.5 (CON) or 25 mM glucose (HG) for 12 h. Slides were then washed with ice-cold PBS, fixed with methanol, and permeabilized using 0.2% Triton X in PBS. Anti-rabbit GPIHBP1 (1:100) and anti-mouse CD31 antibodies (1: 100) were used for immunofluorescence staining. Scale bar, 20 m. B: in a separate experiment, RAOECs were seeded on Transwell inserts and grown until they formed a tight monolayer. Cells were then treated with CON or HG for 4 and 12 h, respectively. In some experiments, this was followed by treatment with or without phosphatidylinositol-specific phospholipase C [PI-PLC; 1 U/ml for 1 h (inset)]; 25 mM mannitol (Mnt) for 12 h was used as an osmotic control. Following the indicated times, these Transwell inserts were removed and placed in a different 6-well plate with DMEM on the basolateral side containing 10 g/ml purified LPL. After incubation for 1 h and washing (3 times) with PBS, medium containing heparin (8 U/ml) was used for 3 min to release apical surface-bound LPL. LPL activity was determined by measuring the in vitro hydrolysis of [3H]triolein substrate. C: using the above protocol (with incubation of HG for 12 h), the heparin-releasable LPL medium was incubated with increasing concentrations of very-low density lipoprotein-triglyceride (VLDL-TG; 0 – 0.8 mM) at 37°C for 30 min and the concentration of released free fatty acid (FA) determined. One group of cells was treated with 1 U/ml PI-PLC prior to incubation with high glucose. *P ⬍ 0.05; n ⫽ 3. E1280 HIGH-GLUCOSE REGULATION OF GPIHBP1 Downloaded from http://ajpendo.physiology.org/ by 10.220.33.1 on August 3, 2017 Fig. 5. Endothelial heparanase (HPA) can increase GPIHBP1 gene and protein expression. A: RAOECs from passages 6 –10 were used for determination of latent (L-HPA) and active heparanase (A-HPA). Cells from passages 7–10 were treated with HG (25 mM) for 12 h, and medium was collected. B: the medium was concentrated using an Amicon column at 4°C and subsequently centrifuged for 15 min at 14,000 g. HPA in this concentrated medium was determined by Western blot; n ⫽ 4. C: recombinant HPA was used to treat RAOECs (passages 7–10) for 12 h, cell lysates were collected, and GPIHBP1 mRNA (top) and protein (bottom) were determined using TaqMan and Western blot, respectively; n ⫽ 3– 4. *P ⬍ 0.05. Fig. 6. Loss of endothelial GPIHBP1 expression can be restored on coculture with myocytes (Myo). A and B: RAOECs (A) or BCAECs (B) (passages 8 –10) were seeded on Transwell inserts and cultured until 80 –90% confluence. The Transwell insert was then transfered to a 6-well plate containing attached Myo and cocultured for 12 h. Isolation of Myo was achieved by collagenase digestion of rat hearts, and cells were attached to laminin-coated 6-well plates in M199 containing 0.1% BSA and kept overnight. After coculture with myocytes, endothelial cell protein was collected and used for Western blot of GPIHBP1. C: in some experiments, endothelial cells (with or without coculture with Myo) were exposed to normal (5.5 mM) or high glucose (HG; 25 mM) for 12 h, and GPIHBP1 was determined; n ⫽ 3– 4. *P ⬍ 0.05 compared with control; #P ⬍ 0.05 compared with normal glucose. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00007.2014 • www.ajpendo.org HIGH-GLUCOSE REGULATION OF GPIHBP1 E1281 Following a single injection of a moderate dose (55 mg/kg) of STZ, there is an induction of hypoinsulinemia and hyperglycemia. Increasing the dose to 100 mg/kg also creates an environment of hyperlipidemia (39). In the former situation, coronary LPL activity is augmented, whereas with the latter setting and the presence of higher circulating FA, LPL is turned off (32). With D55 hearts, in the absence of any change in protein synthesis, the increase in LPL activity principally at the vascular lumen could be explained largely by posttranslational modifications that increased transfer of myocyte enzyme to the ECs (30). Not yet determined is whether the concluding step, moving LPL across EC, is also increased following diabetes. GPIHBP1, a glycosylphosphatidylinositol-an- chored protein expressed exclusively in ECs (15), is a recent addition to the mechanism that transfers LPL across ECs (42). GPIHBP1 has a strong binding affinity for LPL, accepting it from the interstitial space and moving it to the coronary lumen (41). Intriguingly, the mRNA for GPIHBP1 changes much more rapidly than most mRNAs in mammalian cells (22). Our results present the novel observation that to guarantee FA supply to the diabetic heart, the LPL transporter GPIHBP1 increases rapidly in ECs. In an attempt to elucidate the mechanism by which diabetes influences GPIHBP1, we exposed two different EC lines to glucose, a substrate whose concentrations increase rapidly following STZ. However, given that under standard cell culture AJP-Endocrinol Metab • doi:10.1152/ajpendo.00007.2014 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.1 on August 3, 2017 Fig. 7. Endothelial cells and myocyte heparan sulfate proteoglycan (HSPG)-bound platelet-derived growth factor (PDGF) can augment GPIHBP1. A: RAOECs were cultured with or without HG (25 mM) for 12 h, whereas myocytes were incubated with or without A-HPA also for 12 h. With both cell types, the culture medium was collected and tested using a cytokine array. Of the proteins detected, PDGF was chosen. B: purified recombinant PDGF (200 ng/ml) was used to treat RAOECs [passage 7 (left) or 10 (bottom right)] for 12 h, and GPIHBP1 was determined by Western blot. *P ⬍ 0.05; n ⫽ 2– 4. C: summarization of how the endothelial cell increases its GPIHBP1 in response to hyperglycemia. Briefly, HG induces secretion of endothelial HPA, which has multiple functions. These include 1) stimulation of myocyte HSPG-bound LPL release, 2) autocrine and paracrine actions to provoke liberation of PDGF, and 3) reuptake into endothelial cells to influence gene transcription. The latter 2 effects contribute toward augmentation of GPIHBP1, the transporter that moves LPL across EC to its apical side. Out here, LPL is responsible for VLDL-TG hydrolysis to FA, the preferred substrate for the diabetic heart. E1282 HIGH-GLUCOSE REGULATION OF GPIHBP1 HSPG are ubiquitously present in every tissue compartment, particularly the extracellular matrix, cell surface, intracellular granules, and nucleus (17). They consist of a core protein to which several linear HS side chains are covalently linked and function not only as structural proteins but also as anchors due to the high content of charged groups in HS (16). The latter property is implicitly used to electrostatically bind a number of different proteins (chemokines, coagulation factors, enzymes like LPL, and growth factors such as vascular endothelial growth factor) (23). Attachment of these bioactive proteins is a clever arrangement, providing the cell with a rapidly accessible reservoir, precluding the need for de novo synthesis when the requirement for a protein is increased (4). Heparanase is an endoglycosidase, exceptional in its ability to degrade HS, thereby instigating ligand release. Using a cytokine array and incubation of cardiomyocytes with active heparanase, a number of cytokines were increased in the myocyte culture medium. We focused on PDGF, as this protein can regulate angiogenesis but also play a significant role in increasing the metabolic reliance of smooth muscle on FA (35). Favorably, PDGF increased EC GPIHBP1. Because exposure of ECs to high glucose also increased PDGF in the medium, likely through an autocrine stimulus of heparanase secretion with the release of PDGF bound to EC HSPG, our data suggest that this protein “ensemble” (heparanase-PDGF-GPIHBP1) cooperates in the diabetic heart to regulate FA delivery and utilization by the cardiomyocytes. Overall, through their release of heparanase, ECs, which are the first responders to hyperglycemia, are responsible for regulating LPL-derived FA delivery to the cardiomyocytes. Although this mechanism serves to guarantee FA supply and consumption when glucose utilization is compromised, it unintentionally provides a surfeit of FA to the diabetic heart, sponsoring a setting where FA uptake exceeds the mitochondrial oxidative capacity. Chronically, the resulting increase in the conversion of FA to potentially toxic FA metabolites, including ceramides, diacylglycerols, and acylcarnitines, paired with increased formation of reactive oxygen species secondary to elevated FA oxidation, can provoke cardiac cell death (lipotoxicity). GRANTS The present study is supported by an operating grant from the Canadian Diabetes Association. Y. Wang and D. Zhang are the recipients of Doctoral Student Research Awards from the Canadian Diabetes Association. DISCLOSURES No conflicts of interest, financial or otherwise, are declared by the authors. AUTHOR CONTRIBUTIONS A.P.-L.C. and Y.W. conception and design of research; A.P.-L.C., N.L., and B.H. performed experiments; A.P.-L.C. analyzed data; A.P.-L.C., F.W., and I.V. interpreted results of experiments; A.P.-L.C. prepared figures; A.P.-L.C. drafted manuscript; A.P.-L.C., D.Z., A.W., I.V., and B.R. edited and revised manuscript; A.P.-L.C. and B.R. approved final version of manuscript. REFERENCES 1. Adeyo O, Goulbourne CN, Bensadoun A, Beigneux AP, Fong LG, Young SG. Glycosylphosphatidylinositol-anchored high-density lipoprotein-binding protein 1 and the intravascular processing of triglyceride-rich lipoproteins. J Intern Med 272: 528 –540, 2012. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00007.2014 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.1 on August 3, 2017 conditions there has been an insinuation that EC GPIHBP1 is silenced following cell passaging (9, 42), we initially attempted to authenticate this speculation. To our surprise, both rat and bovine aortic ECs lose their ability to express GPIHBP1 protein with increasing subculturing of the cells. However, the introduction of increasing concentrations of glucose to the culture medium increased GPIHBP1 gene and protein expression in both the early and late cell passages. Because high glucose also had comparable effects on LPL shuttling across ECs in association with a greater ability to hydrolyze lipoprotein TG, data from the current study strongly implicate glucose as an important stimulus that influences EC GPIHBP1. Interestingly, high glucose has also been observed to turn on the FA transporters CD36 (13) and FA-binding protein 4 (11) in ECs. Thus, with the onset of hyperglycemia, ECs globally are well adapted to promote FA availability to cardiomyocytes. With EC subculturing, we were interested to discover whether proteins other than GPIHBP1 are also switched off. We focused on heparanase, an endoglycosidase that specifically cleaves HS side chains on HSPGs (4). Intriguingly, in addition to this extracellular function to release bound ligands like LPL, heparanase, by entry into the nucleus to regulate histone acetylation/methylation, is also capable of modulating gene transcription. In this regard, tumor cells, by expressing higher levels of heparanase, are more invasive, as secreted heparanase can breakdown extracellular matrix in addition to promoting gene expression in adjacent cells that drives an aggressive tumor phenotype (27, 28). Similarly to the loss of GPIHBP1, EC passaging also caused nonappearance of heparanase, suggesting a potential connection between these two proteins. Indeed, because the addition of active heparanase to ECs augmented gene and protein expression of GPIHBP1, together with the fact that high glucose induced heparanase secretion, our data for the first time link heparanase to the entire process by which LPL progresses forward during diabetes from the apical side of cardiomyocytes to the apical side of ECs. Notwithstanding the demonstrated impact of high glucose on GPIHBP1, we were unable to explain the disappearance of this glycoprotein on passaging of ECs. A recent study found that in the intact heart, there is a robust expression of EC GPIHBP1, and therefore, it is possible that paracrine signaling influences expression of this glycoprotein (42). Major cell types in the heart include fibroblasts, smooth muscles, and myocytes, with the latter making up almost 60% of the adult rat myocardium (5). We were impressed by our observations that ECs that had lost their ability to express GPIHBP1 regained this glycoprotein when cocultured with cardiomyocytes. Because another study (42) has also hinted at this possibility, our data indicate an interaction between cardiomyocytes and EC likely through paracrine signaling mediators. Currently, it is unclear whether the stimulus for the release of signaling mediators from myocytes could originate from ECs. ECs are first responders to hyperglycemia, releasing heparanase for ensuing hydrolysis of myocyte HSPG, which is linked to liberation of bound proteins. Given that exposure of the coculture to high glucose caused an even greater expression of EC GPIHBP1, our data suggest cellular cooperation between ECs (via heparanase) and myocytes (through unidentified proteins) that facilitates energy switching from glucose to FA in the diabetic heart. HIGH-GLUCOSE REGULATION OF GPIHBP1 23. Lee TY, Folkman J, Javaherian K. HSPG-binding peptide corresponding to the exon 6a-encoded domain of VEGF inhibits tumor growth by blocking angiogenesis in murine model. PloS One 5: e9945, 2010. 24. Lopaschuk GD, Belke DD, Gamble J, Itoi T, Schonekess BO. Regulation of fatty acid oxidation in the mammalian heart in health and disease. Biochim Biophys Acta 1213: 263–276, 1994. 25. Lopaschuk GD, Ussher JR, Folmes CD, Jaswal JS, Stanley WC. Myocardial fatty acid metabolism in health and disease. Physiol Rev 90: 207–258, 2010. 26. Merkel M, Eckel RH, Goldberg IJ. Lipoprotein lipase: genetics, lipid uptake, and regulation. J Lipid Res 43: 1997–2006, 2002. 27. Ogishima T, Shiina H, Breault JE, Tabatabai L, Bassett WW, Enokida H, Li LC, Kawakami T, Urakami S, Ribeiro-Filho LA, Terashima M, Fujime M, Igawa M, Dahiya R. Increased heparanase expression is caused by promoter hypomethylation and up-regulation of transcriptional factor early growth response-1 in human prostate cancer. Clin Cancer Res: 1028 –1036, 2005. 28. Ogishima T, Shiina H, Breault JE, Terashima M, Honda S, Enokida H, Urakami S, Tokizane T, Kawakami T, Ribeiro-Filho LA, Fujime M, Kane CJ, Carroll PR, Igawa M, Dahiya R. Promoter CpG hypomethylation and transcription factor EGR1 hyperactivate heparanase expression in bladder cancer. Oncogene 24: 6765–6772, 2005. 29. Pillarisetti S, Paka L, Sasaki A, Vanni-Reyes T, Yin B, Parthasarathy N, Wagner WD, Goldberg IJ. Endothelial cell heparanase modulation of lipoprotein lipase activity. Evidence that heparan sulfate oligosaccharide is an extracellular chaperone. J Biol Chem 272: 15753–15759, 1997. 30. Pulinilkunnil T, Qi D, Ghosh S, Cheung C, Yip P, Varghese J, Abrahani A, Brownsey R, Rodrigues B. Circulating triglyceride lipolysis facilitates lipoprotein lipase translocation from cardiomyocyte to myocardial endothelial lining. Cardiovasc Res 59: 788 –797, 2003. 31. Pulinilkunnil T, Rodrigues B. Cardiac lipoprotein lipase: metabolic basis for diabetic heart disease. Cardiovasc Res 69: 329 –340, 2006. 32. Rodrigues B, Cam MC, Jian K, Lim F, Sambandam N, Shepherd G. Differential effects of streptozotocin-induced diabetes on cardiac lipoprotein lipase activity. Diabetes 46: 1346 –1353, 1997. 33. Rodrigues B, Cam MC, McNeill JH. Myocardial substrate metabolism: implications for diabetic cardiomyopathy. J Mol Cell Cardiol 27: 169 – 179, 1995. 34. Rodrigues B, Spooner M, Severson DL. Free fatty acids do not release lipoprotein lipase from isolated cardiac myocytes or perfused hearts. Am J Physiol Endocrinol Metab 262: E216 –E223, 1992. 35. Salabei JK, Hill BG. Mitochondrial fission induced by platelet-derived growth factor regulates vascular smooth muscle cell bioenergetics and cell proliferation. Redox Biol 1: 542–551, 2013. 36. Sambandam N, Abrahani MA, Craig S, Al-Atar O, Jeon E, Rodrigues B. Metabolism of VLDL is increased in streptozotocin-induced diabetic rat hearts. Am J Physiol Heart Circ Physiol 278: H1874 –H1882, 2000. 37. Sambandam N, Abrahani MA, St Pierre E, Al-Atar O, Cam MC, Rodrigues B. Localization of lipoprotein lipase in the diabetic heart: regulation by acute changes in insulin. Arterioscler Thromb Vasc Biol 19: 1526 –1534, 1999. 38. Wang F, Kim MS, Puthanveetil P, Kewalramani G, Deppe S, Ghosh S, Abrahani A, Rodrigues B. Endothelial heparanase secretion after acute hypoinsulinemia is regulated by glucose and fatty acid. Am J Physiol Heart Circ Physiol 296: H1108 –H1116, 2009. 39. Wang Y, Puthanveetil P, Wang F, Kim MS, Abrahani A, Rodrigues B. Severity of diabetes governs vascular lipoprotein lipase by affecting enzyme dimerization and disassembly. Diabetes 60: 2041–2050, 2011. 40. Wang Y, Zhang D, Chiu AP, Wan A, Neumaier K, Vlodavsky I, Rodrigues B. Endothelial heparanase regulates heart metabolism by stimulating lipoprotein lipase secretion from cardiomyocytes. Arterioscler Thromb Vasc Biol 33: 894 –902, 2013. 41. Williams KJ. Molecular processes that handle — and mishandle — dietary lipids. J Clin Invest 118: 3247–3259, 2008. 42. Young SG, Davies BS, Voss CV, Gin P, Weinstein MM, Tontonoz P, Reue K, Bensadoun A, Fong LG, Beigneux AP. GPIHBP1, an endothelial cell transporter for lipoprotein lipase. J Lipid Res 52: 1869 –1884, 2011. 43. Zechner R, Kienesberger PC, Haemmerle G, Zimmermann R, Lass A. Adipose triglyceride lipase and the lipolytic catabolism of cellular fat stores. J Lipid Res 50: 3–21, 2009. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00007.2014 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.1 on August 3, 2017 2. An D, Rodrigues B. Role of changes in cardiac metabolism in development of diabetic cardiomyopathy. Am J Physiol Heart Circ Physiol 291: H1489 –H1506, 2006. 3. Augustus AS, Kako Y, Yagyu H, Goldberg IJ. Routes of FA delivery to cardiac muscle: modulation of lipoprotein lipolysis alters uptake of TGderived FA. Am J Physiol Endocrinol Metab 284: E331–E339, 2003. 4. Bame KJ. Heparanases: endoglycosidases that degrade heparan sulfate proteoglycans. Glycobiology 11: 91R–98R, 2001. 5. Banerjee I, Fuseler JW, Price RL, Borg TK, Baudino TA. Determination of cell types and numbers during cardiac development in the neonatal and adult rat and mouse. Am J Physiol Heart Circ Physiol 293: H1883– H1891, 2007. 6. Blanchette-Mackie EJ, Masuno H, Dwyer NK, Olivecrona T, Scow RO. Lipoprotein lipase in myocytes and capillary endothelium of heart: immunocytochemical study. Am J Physiol Endocrinol Metab 256: E818 – E828, 1989. 7. Davies BS, Beigneux AP, Barnes RH 2nd, Tu Y, Gin P, Weinstein MM, Nobumori C, Nyrén R, Goldberg I, Olivecrona G, Bensadoun A, Young SG, Fong LG. GPIHBP1 is responsible for the entry of lipoprotein lipase into capillaries. Cell Metab 12: 42–52, 2010. 8. Davies BS, Goulbourne CN, Barnes RH 2nd, Turlo KA, Gin P, Vaughan S, Vaux DJ, Bensadoun A, Beigneux AP, Fong LG, Young SG. Assessing mechanisms of GPIHBP1 and lipoprotein lipase movement across endothelial cells. J Lipid Res 53: 2690 –2697, 2012. 9. Davies BS, Waki H, Beigneux AP, Farber E, Weinstein MM, Wilpitz DC, Tai LJ, Evans RM, Fong LG, Tontonoz P, Young SG. The expression of GPIHBP1, an endothelial cell binding site for lipoprotein lipase and chylomicrons, is induced by peroxisome proliferator-activated receptor-gamma. Mol Endocrinol 22: 2496 –2504, 2008. 10. Eckel RH. Lipoprotein lipase. A multifunctional enzyme relevant to common metabolic diseases. N Engl J Med 320: 1060 –1068, 1989. 11. Elmasri H, Karaaslan C, Teper Y, Ghelfi E, Weng M, Ince TA, Kozakewich H, Bischoff J, Cataltepe S. Fatty acid binding protein 4 is a target of VEGF and a regulator of cell proliferation in endothelial cells. FASEB J 23: 3865–3873, 2009. 12. Enerbäck S, Gimble JM. Lipoprotein lipase gene expression: physiological regulators at the transcriptional and post-transcriptional level. Biochim Biophys Acta 1169: 107–125, 1993. 13. Farhangkhoee H, Khan ZA, Barbin Y, Chakrabarti S. Glucoseinduced up-regulation of CD36 mediates oxidative stress and microvascular endothelial cell dysfunction. Diabetologia 48: 1401–1410, 2005. 14. Haffner SM. Lipoprotein disorders associated with type 2 diabetes mellitus and insulin resistance. Am J Cardiol 90: 55i–61i, 2002. 15. Ioka RX, Kang MJ, Kamiyama S, Kim DH, Magoori K, Kamataki A, Ito Y, Takei YA, Sasaki M, Suzuki T, Sasano H, Takahashi S, Sakai J, Fujino T, Yamamoto TT. Expression cloning and characterization of a novel glycosylphosphatidylinositol-anchored high density lipoproteinbinding protein, GPI-HBP1. J Biol Chem 278: 7344 –7349, 2003. 16. Iozzo RV. Heparan sulfate proteoglycans: intricate molecules with intriguing functions. J Clin Invest 108: 165–167, 2001. 17. Iozzo RV, San Antonio JD. Heparan sulfate proteoglycans: heavy hitters in the angiogenesis arena. J Clin Invest 108: 349 –355, 2001. 18. Kim MS, Kewalramani G, Puthanveetil P, Lee V, Kumar U, An D, Abrahani A, Rodrigues B. Acute diabetes moderates trafficking of cardiac lipoprotein lipase through p38 mitogen-activated protein kinasedependent actin cytoskeleton organization. Diabetes 57: 64 –76, 2008. 19. Kim MS, Wang F, Puthanveetil P, Kewalramani G, Hosseini-Beheshti E, Ng N, Wang Y, Kumar U, Innis S, Proud CG, Abrahani A, Rodrigues B. Protein kinase D is a key regulator of cardiomyocyte lipoprotein lipase secretion after diabetes. Circ Res 103: 252–260, 2008. 20. Kim MS, Wang F, Puthanveetil P, Kewalramani G, Innis S, Marzban L, Steinberg SF, Webber TD, Kieffer TJ, Abrahani A, Rodrigues B. Cleavage of protein kinase D after acute hypoinsulinemia prevents excessive lipoprotein lipase-mediated cardiac triglyceride accumulation. Diabetes 58: 2464 –2475, 2009. 21. Kim MS, Wang Y, Rodrigues B. Lipoprotein lipase mediated fatty acid delivery and its impact in diabetic cardiomyopathy. Biochim Biophys Acta 1821: 800 –808, 2012. 22. Kroupa O, Vorrsjö E, Stienstra R, Mattijssen F, Nilsson SK, Sukonina V, Kersten S, Olivecrona G, Olivecrona T. Linking nutritional regulation of Angptl4, Gpihbp1, and Lmf1 to lipoprotein lipase activity in rodent adipose tissue. BMC Physiol 12: 13, 2012. E1283