Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
033rd MEETING. LONDON
Two distinct populations of labelled peptides were
ohtained. one with molecular masses in the range 28-45 kDa
and the other in the range 3- 13 kDa. T h e former population
appears to include N-terminal regions o f the mAChRs
sequences which contain the N-glycosylation sites. T h e
higher molecular mass population corresponds to that
ohtained when mAChRs are labelled with the highly specific alkylating antagonist propylhenzilylcholine mustard
(PrBCM)and cleaved with Lys-C under the same conditions
151.
T h e [ 'HINEM-labelled peptides were further separated hy
f.p.1.c. gel-filtration and fractions enriched in the higher
molecular mass species were subcleaved with cyanogen
bromide (CNBr; 4 0 mg/ml) as descrihcd previously (51.
Analysis by SDS/PAGE on 18-22'L gradient gels detected
two major labelled peptides with molecular masses ahout 3.9
k D n and 2.7 kDa.
These peptides seem identical in molecular mass t o the
major peptides formed hy CNBr cleavngc o f 13HIPrBCMlahellcd mAChRs. suggesting that the lahellcd cystcine
residue is close to the ('HIPrBCM attachment site. Peptide
sequencing experiments suggest that the 3.1) kI>a peptide is
formed hy cleavage at the conserved methionine residues in
T M 2 and T M 3 (corresponding t o Met-79/1 14 in the ml
sequence) and the 2.7 kDa peptide hy cleavage at methionine
residues in the first extracellular loop and in T M 3 (corresponding t o Met-XX/I 14 in the m l sequence), and that the
I 'ti IPrBCM estcrifies a conserved aspartic acid residue in
T M 3 (corresponding to Asp-105 in the ml sequence) 161.
fidman degradation of the two I 'HINEM-labelled
peptides gave release o f label after I 0 and 1 Y sequenator
443
cycles. This is consistent with lahclling o f conserved cysteine
residue in the first extracellular loop (corresponding t o Cys98 in the m l sequence). This result strongly supports the
participation of this residue in an intramolecular disulphide
hridge.
Preliminary results suggest that ii conserved cystcine
residue in the second extracellular loop of muscarinic reccptors (corresponding t o Cys-178 in the m l sequence) could
possihly be the other component o f the disulphide bond.
Further experiments must be undertaken t o confirm these
results and t o search for other possihle disulphide bonds in
the muscarinic receptors.
I , Honner. 7'.I.. Hucklcy. N. J., Young. A. <'. YC I3rann. M. K. . S c i c w w
231.527532
2. Bcrstein, (3.. Haga. K.. Haga, I'.di Ichiyama. A. ( I W X ) J. iVc,irroc.licvn. 50. I hX7- 1604
3. Nishiyama. '1. .. I3crstein. G , . lkcgayii. 'I' .. Hngn. '1. .. Ichiyom;i. A..
Kohayashi. A. YC Yamazaki. N . ( I UXO) /3ioc./ic,nr. Re,.\. 10.
25 1-260
4, Florio. V. A. & Stcrnwei\. f'. ( ICMO)i J. / h i / . C ' / i e , ~ i . 264.
3909-3Y I5
5. Curtis. C. A. M.. Whcatley. M.. 13ansol. S., Hirdsall. N. J. M..
Eveleigh. P.. Pcdder. ti. K. YC Hulmc. E. <'. ( I (NU)
J. Iliol. <'/icm.
264.4XV-495
0 . Hulme, E. C . . Curtis, C . A. M.,Whcatley, M.. Harris. A. (.. M. di
Aitken, A. ( 10x9)) /rcvitl.\ I'/irrrt?iccc~ol.. S u . 10 (.Siippl. .Sirh/y/ic\ o/
h~rr.sc~crrirri1~
/<c~c~c~/l/or.s
IV ), 2 2 - 2 5
Kcceived 23 Novcmhcr I 9 X 9
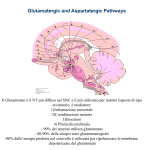
Glutamic acid concentration in brains of patients with Alzheimer's Disease
S'l'til'tlt" L. LOWE and DAVID M. BOWEN
,tliri(irtI h1rrX.s Ikpcirirticvri (fN~~iiroc~/ier7ristt~~,
Itistinrtc of
Nc,rrrolop ( C ) i w c t i Scpuirc). Uriiivr.si!\' o/'Loticloti.
I \\'irkc~/icltl Strwr, Loticlori WC'I N II'J. U.K.
T h e glutamic acid content o f post-mortem Alzheimer's
tlisensc ( A D ) brain tissue has heen found reduced I 1-5 I, hut
not consistently 16-I II. However. shortly after death. cxtensive alterations in amino acid content occur with increasing
post-mortem delay I I 2 I and changes pcvi-rrioricrii may affect
the rate o f autolysis 151. To invehtigate glutamate status
further. the concentrations of glutamate and a number o f
other amino acids have been determined in samples ohtained
hy various neurosurgical procedures.
('ortical grey matter from frontal o r temporal lohe was
olitaincd. and maintained at - 196°C. from I 0 patients with
AD. 5 with other dcmentias. 3 Y controls and 1 8 frontal lobe
samples from patients with intractable depression 15, I 3 1.
Tissue containing all cortical layers. free o f meninges and
underlying white matter. was extracted for putative cortical
tr;insniitter amino acids (aspartate. glutamate and taurinc)
and non-transmitter amino acids (alanine. asparagine, glutamine. glycine, isolcucine, Ieucine, mcthioninc. ornithinc,
phcnylalanine. serinc and tryptophan) which were determined by h.p.1.c. with fluorometric detection 15 ]. Nucleolated
pyramidal nerve cell packing density in two cortical layers
and the frequency o f senile plaques and neurofibrillary
tangles were determined in sections from the A D samples
I 14I. Results are expressed as mean f s.11Single comparisons
were made by either the Student's I-test o r Mann Whitney Utest. ;IS appropriate (Fisher I.-test). Multiple comparisons
A1ilircvi;ition i r x d A I). Alzhcimer'h disease.
Vol. I8
were made hy either one-way analysis of variance (ANOVA ).
followed by the least significant difference test (probabilities significant when I'< 0.05) o r Krusknll-Wallis ANOVA
and the Mann Whitncy U-test (null hypothesis rejected at
I'< 0.0 I), as appropriate (Fisher I.-test). Intercorrelation of
variables was tested by Spearman's rank ( l i , )correlation.
T h e effect o f various factors on amino acid values were
assessed by dividing suhjects into appropriate subgroups 15 1.
None o f the values in any subject group were influenced hy
age. gender. drug treiitnient ( A D group) o r associated lesion
types (control group). There w a s no difference (/'> 0.05)
hetween frontal and temporal cortex (control group) in the
content o f any non-transmitter amino acid s o values have
hcen pooled t o increase the statistical power o f the study.
T h e mean values o f most amino acids are higher in A D
compared with control. T h e individual increases ;ire smnll.
hut the sum ( I5 nmol) o f the seven significantly affected (five
in Fig. I.plus methioninc and ornithine. data not shown) is
very similar t o the reduction in glutamate content ( I 0 nmol)
which is the only significant reduction in A D (Fig. I ) . A
correlation was found hetween glutamate content and only
neuron packing density in layer 111 (Table I ). No significant
differences occur in the group o f other dementias. but the
mean glutamate content is low. Glutamate is not reduced in
the depressives. Titurine content (nmol/nig o f protein) o f A D
temporal cortex (23.5f 8.1, t i = 7 ) is unchanged from control
(22.6k 6.7. I1 = 15).
In this series. some pyramidal neurons have disappeared
from the neocortex. whereas interneurons seem spared [ 14.
15 I. T h e resulting atrophy o f A D hrain tissue may lead to iiii
increase in constituents o f unaffected structures. Such selective degeneration o f structures enriched in glutamate I 10. I 7 1
probably explains the increased content o f amino acids. T h e
444
BIOCHEMICAL SOCIETY TRANSACTIONS
Glutamate
U
*U
T
T
Aspartate
I
I
*
Alanine
Serine
*
most straightforward explanation for the one relationship
(Table 1 ) is that degeneration o f corticocortical association
tracks (see IS])has had a major influence on the glutamate
content of the samples. Estimates of the glutamate transmitter pool, based on experimental paradigms, vary from 1 0
to 40% o f total glutamate content [ 16, 171. The latter is
reduced by 14% in A D temporal cortex (Fig. 1). so at least
35% of the transmitter glutamate may have been lost, though
metabolic changes IS] and neuronal loss may complicate this
interpretation. The A D samples were obtained shortly after
emergence of symptoms 15. I S ] . Thus glutamate deficiency
seems to be quite an early change that is selective, by comparison with other amino acids (including y-aminobutyrate
and other transmitters, somatostatin, dopamine and nordrenaline [ 181). not a feature of depression and probably is
due to degeneration of glutamatergic corticocortical association tracks. Is this due to excitotoxicity and will either
glutamate antagonists [ 191 o r manipulation of oxidative metabolism [ 201 retard disease progression?
Dr D. M. A. Mann kindly provided the morphological measurements.
$2
Z E
-c
*
*-
41
Leucine
*
2 t
Isoleucine
1
2
0
0
Fig. 1. Amitio acids altered in A D
.,
Non-transmitter amino acid values are for 10 AD, 5 other
dementia, I8 depressives and 39 controls. 0,Control; H,
AD; 0,other dcmentias;
depressives. Glutamate values
(nmol/mg o f protein), aspartate in parentheses, are
1OS.Xf32.2 ( 2 4 . 7 f 6 . 1 ) , 82.6f20.1 ( 2 0 . 1 f 3 . 0 ) and
98.6 f 15.9 ( 19.6 k 3.7) for 18 depressives, 5 other
dementia and 24 controls (frontal cortex), respectively;
values for 7 A D and 15 controls (temporal cortex) are as
described [ 51. *Difference from control, P < 0.05 (single
comparison); **Significantly different from control (multiple
comparison).
Table 1 . (‘oric,enrrcirion of’ glutumic acid and morphologit,ul
mcw.sitri’.s in [hi)remporul cortex
Results are for individual A D subjects. Glutamic acid content
and morphological values a r e nmol/mg of protein and number/
rnm?, respectively. Tissue glutamate correlates with only neuron
density of layer 111 (H,= 0.75; /’= 0.05).
Subject
I
-1
3
4
5
Ti\sue
glutamate
I20
I I8
6
105
96
88
84
7
76
Pyramidal neurone
density
Tangle
frequency
Plaque
frequency
Layer I l l
Layer V
20 1
287
27
18
171
1x1
151
29
19
242
I94
205
259
13
29
I97
31
7
13
8
16
20
I17
I60
I37
I24
16
15
I . Arai. H.. Kobayashi, K.. Ichimiya. Y.. Kosaka, K. & lizutca. K.
( 1984) Niwrohiol. Agitig 5 . 3 I 9-32 I
2. Ellison, D. W., Beal, M. F., Mazurek, M. F., Bird. E. D. &
Martin. J. H. ( 1986) Anti. Neitrol. 2 0 . 6 16-62 I
3. Sasaki, H., Murarnoto. 0..Kanazawa, I., Arai. H., Kosaka. K. &
lizaka. K. ( 1986) Atin. Ncitrol. 19. 263-260
4. Hyman. €3. T.. Van Hoescn. G. W. & Damansio. A. K. (1987)
h i t i . Neitrol. 2 2 . 37-40
5. Procter, A. W.. Palmer. A. M., Francis. P. I..,Lowe. S. L., Neary.
E., Murphy. E.. I>oshi. K. & Howen. I>. M. ( I 9 8 8) J. N w r o them. SO, 790-802
6. larbit. I.. Perry. E. K.. Perry. K. H.. Blessed. G. & ‘l‘omlinson.
H. E. ( I98 I ) J. Neirrochcm. 35. 1246- I249
7. Antuono. P. G.. Lchmann. J.. Koller. K. J.. Struhle. R. G,. Price.
D. L.. Coyle, J. I,.,Whitehouse. P. J. & Clark. A. W. (1984)
Neitrology 34 (Suppl. I ), I I 9
8. Perry. E. K., Atack, J. K.. Perry, K. H., Hardy, J. A,. Ilodd. P. K.,
Edwardson, J. A,, Blessed, G.. Tomlinson. B. E. & Fairbairn.
A. F. ( 1984) J. N w r o c h e m . 42. 1402- I4 I0
9. Perry, T. L.. Yong. V. W.. Hergeron. C.. Hansen. S. di Jones. K.
(I987)Anti.N ( w r o l . 2 1 , 3 3 1 - 3 3 6
1 0 . Procter, A. W., Lowe, S. L., Palmer, A. M., Francis, P. I..,Esiri,
M. M.. Stratmann, G. C.. Najlerahim. A,. Patel, A,. Hunt, A. &
Howen. I>. M. ( I 9 8 8 ) J. N w r o l . S c i . 84, 125- 140
I I . Reynolds, G. P., Warner, P. & Mercer. K. ( 1989) J. Noiirul
7rutism. 1 , I I8
12. Perry. 7.. L.. Hanscn. S. di Gandham, S. S. ( I98 I ) J. Ncwrockni.
36,406-4 I 2
13. Francis, P. ‘I.., Poynton. A,. Lowe. S. L.. Pearson, K. C. A,.
Bridges. P. K.. Hartlctt, J. K.. Procter. A. W. & Bowen. I>. M.
( 1989) Hruiti K(,.s. 494. 3 15-324
13. Neary. I>.. Snowden. J. S.. Mann. I>. M. A,. Howen, I>. M.. Sims.
N. K.. Northen. 13.. Yates. P. 0. & Ihvison. A. N. (1986) J.
Niwrol. Noitrosittg. I’.sydiiut. 49, 220- 237
15. Lowe, S. L.. Francis. P. T., Procter, A. W., Palmer. A. M.,
Davison. A. N. & Bowen, D. M. (1988) Bruin 11 1,785-799
16. Fonnum, F. ( 1984) J. Neirrochem. 42, 1 - 1 1
17. Nicholas, D. G., Sihra. T. S. & Sanchez-Prieto, J. (1987) J.
Neurochem 49.50-57
18. Francis. P. T. & Bowen. D. M. (1989) C’uti. J. Nritrol. Sci. 16.
504-5 10
19. Greenamyre. J. T. & Young. A. B. ( 1989) Neitrohiol. Aging 10,
593-602
20. Sims, N. R., Finegan, J. H., Blass. J. P.. Bowen, D. M. & Neary,
D. ( 1987) Bruin Rev. 436, 30-58
Received 20 November I989
1990