Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
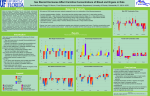
Indian Journal of Experimental Biology Vol. 48, May 2010, pp. 503-510 Effect of carnitine supplementation on mitochondrial enzymes in liver and skeletal muscle of rat after dietary lipid manipulation and physical activity Jyothsna Karanth & K Jeevaratnam1* Biochemistry and Nutrition Discipline, Defence Food Research Laboratory, Mysore 570 011, India Received 25 September 2009; revised 14 December 2009 Effect of carnitine supplementation in enhancing fat utilization was investigated by looking into its effects on mitochondrial respiratory enzymes activity in liver and muscle as well as on membrane fatty acid profile in rats fed with hydrogenated fat (HF) and MUFA-rich peanut oil (PO) with or without exercise. Male Wistar rats were fed HF-diet (4 groups, 8 rats in each group) or PO-diet (4 groups, 8 rats in each group), with or without carnitine for 24 weeks. One group for each diet acted as sedentary control while the other groups were allowed swimming for 1 hr a day, 6 days/week, for 24 weeks. The PO diet as well as exercise increased the activities of mitochondrial enzymes, NADH dehydrogenase, NADH oxidase, cytochrome C reductase, cytochrome oxidase, while carnitine supplementation further augmented the oxidative capacity of both liver and muscle significantly by enhancing the activity of carnitine palmitoyl transferase and the respiratory chain enzymes. These effects can be attributed to the enhanced unsaturated fatty acids in phospholipids of mitochondria and may be due to increased fluidity of the membrane in these rats. Results of this study show a significant health promoting effects of carnitine supplementation which could be further augmented by regular exercise. Keywords: Carnitine supplementation, Dietary fat, Liver, Membrane phospholipids composition, Mitochondrial enzymes, Physical activity, Skeletal muscle Dietary fat and exercise are known determinants of good health. Many a studies have shown that the different type of dietary fat influenced the fatty acid composition of plasma and cellular membranes1,2 including mitochondrial membrane and their function3,4,5. Fatty acid compositional changes due to dietary fat (monounsaturated or polyunsaturated fatty acids) were associated with extensive changes in mitochondrial enzyme activities3. Carnitine palmitoyl transferase (CPT) activity is also influenced by dietary fat4. It has been reported that in addition to dietary fat (virgin olive oil or sunflower oil); physical exercise also altered the fatty acid profile of mitochondrial membrane5 and increased the activity of cytochrome oxidase in liver and skeletal muscle6. However, there have been contradicting reports about the influence of physical training on hepatic mitochondrial function, i.e., either a decrease in oxidative capacity7 or no change in the oxidative capacity or an increased cytochrome oxidase activity6. ________________ * Correspondent author 1 Present address : Department of Biochemistry and Molecular Biology, Pondicherry University, Kalapet, Puducherry 605 014, India Telephone: 0413-2654421 (O); 0413-2256813 (R) E-mail: [email protected] A higher level of CPT activity was observed in muscle of endurance athletes8. Supplementation of L-Carnitine, an obligatory cofactor for mitochondrial fatty acid oxidation, has been shown to increase pyruvate dehydrogenase complex, NADH–cytochrome C reductase, succinate–cytochrome C reductase and cytochrome oxidase in muscle of endurance athletes9, while the CPT activity remained unchanged8. L-carnitine supplementation could enhance fat utilization in rats fed either hydrogenated fat (HF) or MUFA-rich peanut oil (PO) diet with or without exercise10. Dietary fat type and regular exercise are known to modulate fatty acid profile of mitochondrial membranes and associated with extensive changes in the activity of mitochondrial enzymes. Carnitine supplementation has been hypothesized to improve exercise performance in humans by increasing respiratory chain enzymes activity. Previously, we have reported that L-carnitine supplementation could enhance fat utilization in rats fed either hydrogenated fat (HF) or MUFA-rich peanut oil (PO) diet with or without exercise10. Hence, the present study was undertaken to assess the influence of the type of dietary fat and regular exercise of moderate intensity (swimming for 1 hr) for 6 months duration on the liver mitochondrial membrane lipid profile and the activities 504 INDIAN J EXP BIOL, MAY 2010 of enzymes such as NADH dehydrogenase, NADH oxidase, cytochrome C reductase, cytochrome oxidase and carnitine palmitoyl transferase in liver and muscle mitochondria, and also to assess the effect of dietary supplementation of carnitine on these parameters. The dietary fats chosen were hydrogenated fat and peanut oil. The former is used in the preparation of food for Indian armed forces as it gives long shelf life, while the peanut oil is predominantly used in southern part of India. Materials and Methods The Meals-Ready to-Eat (MRE) ration developed at Defence Food Research Laboratory, Mysore for Indian armed forces was used as diet for the study. Each ration pack consisted of sooji halwa, vegetable pulav, chapati, potato peas curry, soft bar, tealeaves, skimmed milk powder and sugar. The ration was prepared either with hydrogenated vegetable fat, vanaspati (HF) as source of SFA or with refined peanut oil (PO) as a source of MUFA-rich fat. Carbohydrates, fat and proteins contributed 58, 27 and 15 % respectively, of the total energy in this diet. The fatty acid profile of the Indian vanaspati (Dalda) and peanut oil analyzed by gas chromatography has been reported earlier11. Chemicals used were of AR grade obtained from either E.Merck or SRL (India). The fatty acid methyl esters (FAME) standards were procured from Sigma (USA). L-Carnitine as LCarnitine tartarate was kindly gifted by Lonza, Basel (Switzerland). L- Carnitine was given at the level of 0.5 % on dry weight basis of diet. Experimental protocol⎯Male Wistar rats (64), bred in animal house of Defence Food Research Laboratory, Mysore, and weighing 140 ± 10 g were used. The Institutional Animal Ethics Committee had approved this animal experimentation. Rats were housed individually in stainless steel cage with a 12hr L/D cycle, relative humidity and temperature under control. After being on the MRE ration diet for 2 weeks rats were grouped randomly into 8 groups of 8 rats in each group. Groupings and treatment protocol are summarized in Table 1. Rats were given preweighed diet and water ad libitum. Left out diet in each cage was collected over a week to quantify the food consumption of rats weekly for 6 months of experimental period. The composition of the diet has been reported earlier10. Tissue preparation and analytical methods⎯At the end of the experimental period, the animals were allowed to rest for about 20 hr after the exercise; fasting blood was drawn into heparinised tube and later killed by cervical dislocation between 9-10 am. Liver and muscle from left hind limb were excised, rinsed in ice-cold physiological saline and kept on ice until processed. Blood was centrifuged to obtain plasma, while the sediment after removing the buffy coat was washed twice with isotonic phosphate buffer, pH 7.5 to obtain pure erythrocytes. The plasma, a part of liver and muscle were immediately frozen and stored at 20oC until the analysis of free carnitine according to the method of Wieland et al12. A part of erythrocytes was used to determine the osmotic fragility according to Kaplay13. Results were expressed as % haemolysis in 0.4% NaCl, taking the haemolysis in 0.1% NaCl as 100%. The remaining erythrocytes were used for the preparation of ghosts according to the procedure of Dodge et al14. The erythrocytes were lysed with 20 milliosmolar (mOsm) phosphate buffer, pH 7.5 and centrifuged at 20000 g for 40 min. The pellet (ghosts) was washed thrice subsequent to haemolysis. The ghosts were then incubated in 320 mOsm (isotonic) phosphate buffered saline solution, pH7.5 at 37oC for 1 hr during which the membrane resealed15,16. The membrane protein was estimated by Lowry et al17. Pyrene was used as the fluorescent probe to monitor the fluidity of erythrocyte membrane because of its lateral diffusing capability in the hydrophobic region of the membranes18. Incorporation of pyrene into the ghost membrane was accomplished by adding a concentrated solution of pyrene in alcohol to the aqueous suspension of membranes (0.2 mg of erythrocyte membrane protein) and incubating at 25oC with gentle stirring for 30 min. Steady state fluorescence measurements were performed at 90o to the exciting beam, 342 nm using Elico SL 174 spectrofluorometer. Dimer: monomer (D/M) ratios were estimated by comparing the fluorescence intensity Table 1⎯Treatment groups Group 1 2 3 4 5 6 7 8 Treatment HFS – fed with hydrogenated fat diet and sedentary POS – fed with peanut oil (MUFA-rich) diet and sedentary HFE – Exercising HF POE – Exercising PO HFCRS – Sedentary HF supplemented with carnitine POCRS – Sedentary PO supplemented with carnitine HFCRE – Exercising HF supplemented with carnitine POCRE – Exercising PO supplemented with carnitine KARANTH & JEEVARATNAM: CARNITINE, DIETARY LIPID, EXERCISE & MITOCHONDRIAL ENZYMES at 480 nm to that at 386 nm, using 342 nm as the excitation wavelength in the erythrocyte ghosts incorporated with various concentrations (10-30 µM) of pyrene16. Liver was homogenized in ice-cold buffered 0.25 M sucrose containing 1 mM ethylene diamine tetra acetic acid (EDTA) and used for preparation of mitochondria as described elsewhere19. Muscle mitochondria was prepared from finely minced muscle tissue and gently homogenized with a manual glass homogenizer in icecold Tris buffer cocktail as described by Jackman and Willis20. Protein was determined by the modified biuret method21. The frozen and thawed mitochondria from liver and muscle were used for various mitochondrial enzyme assays. Standard spectrophotometric procedures were used for the assay of NADH oxidase, NADH dehydrogenase, NADH cytochrome C reductase and cytochrome oxidase19. The carnitine palmitoyl transferase (CPT) was assayed according to the method of Bieber et al22. Lipid peroxidation in these mitochondria was also assessed based on malondialdehyde (MDA) formation and assayed by the thiobarbituric acid method after washing mitochondrial preparation with phosphate buffer to remove sucrose in the samples23. The remaining hepatic mitochondria were used for the extraction of fat and analysis of fatty acid profile of phospholipids. The lipids were extracted according to Folch procedure24 using chloroform and methanol at 2:1 containing 0.01 % butylated hydroxy toluene (BHT) as antioxidant. Thin layer chromatographic separation of phospholipids was carried out on alkaline silica gel. The plates (20 cm and 0.25 mm layer thickness) were prepared with alkaline silica gel G (E. Merck) using 0.01 M Na2CO3 solution and were activated for 1 hr at 110oC before use. The plates were first developed with hexane/methanol/acetone (80:20:1). After air drying for 30 min, they were subsequently developed at right angle with chloroform/methanol/glacial acetic acid/water (50:45:3.5:0.15). Iodine vapors were used for detection and identification of the various fractions by comparing with Rf of authentic standards. The fractions corresponding to phosphotidylcholine, phosphotidylethanolamine and cardiolipin were scrapped from the TLC plate and eluted with chloroform/methanol (2:1 containing 0.01 % BHT). The individual fractions were esterified in methanolic BF3. The fatty acid methyl esters (FAME) were then 505 analysed by gas liquid chromatography in a Chemito 8610 model GC. The column used was Supelco BPX 70. Column temperature was set at 110 to 220o C, 6oC /min, N2 was used as carrier gas using flame ionization detector. The fatty acids (FAME’s) were identified by comparison with authentic standard mixtures. Peak areas of individual fractions were calculated by an integrator and expressed as relative percents of total fatty acids extracted. Statistical analysis⎯The results represent mean ± standard error of values from 8 animals. A two-way ANOVA was performed to evaluate the effect of dietary fat and physical activity on each variable. Significant (P<0.05) interaction terms were evaluated by Duncan’s test. Results There was no appreciable difference in body weight or weight gain between the groups fed on HF and PO, sedentary as well as exercising groups. Food consumption was not significantly different across the various groups though there was a mild increase in exercising rats (data not shown). Effect on plasma and tissue free carnitine level⎯The plasma free carnitine was higher in POfed rats as compared to HF-fed ones, while tissue level was not altered (Table 2). Regular exercise decreased the free carnitine level in plasma, liver and muscle from all the groups of non-carnitine supplemented rats. There was a significant increase in free carnitine level in all carnitine supplemented ones including exercising rats as compared to corresponding non-supplemented groups. Effects on liver and muscle mitochondrial CPT activity⎯Dietary fat had no influence on the hepatic Table 2⎯Effect of dietary fat, carnitine and exercise on free carnitine in plasma, muscle and liver Groups* HFS POS HFE POE HFCRS POCRS HFCRE POCRE Plasma Liver (nmol/g) Muscle(nmol/g) (µmol/L) [Values are mean + SE of 8 rats] 118 ± 6a 178 ± 14a 33.2 ± 1.7a b b 138 ± 3 211 ± 18b 37.9 ± 1.6 30.0 ± 1.8a 105 ± 3a 146 ± 18c a a 117 ± 4 187 ± 9a 32.9 ± 1.2 c c 53.6 ± 3.1 180 ± 3 328 ± 18d 194 ± 9c 337 ± 12d 64.3 ± 2.0d c d 51.1 ± 2.0 163 ± 6 350 ± 13d 195 ± 7c 365 ± 20d 59.2 ± 1.8d Values in the same column that do not share the same superscript letters are significantly different at P<0.05. * Groups as in Table 1 INDIAN J EXP BIOL, MAY 2010 506 and muscle mitochondrial CPT activity except for an increased in PO fed rats, while regular exercise increased this enzyme activity in both the tissues (Table 3). The carnitine supplementation further increased CPT activity in both the tissues, more so in HFCRE and POCRE rats. Effect on activities of liver and muscle mitochondrial enzymes⎯Activities of hepatic mitochondrial enzymes, NADH dehydrogenase, NADH oxidase, cytochrome C reductase, cytochrome oxidase were significantly higher in PO fed group. Regular exercise further increased these activities. Activities of these enzymes were significantly higher in carnitine fed rats from both the dietary groups as compared to non-carnitine supplemented ones more so in PO fed exercising rats (Table 3). Dietary fat did not alter the activities of muscle mitochondrial enzymes while regular exercise was found to enhance the activity of these enzymes in both the dietary conditions. Carnitine fed rats showed significantly higher activities of these enzymes as compared to non-carnitine supplemented ones more so in HFCRE and POCRE groups (Table 3). Effect on the fatty acid composition of hepatic mitochondrial membrane⎯Fatty acid composition of PC was greatly influenced by the dietary fatty acids (Table 4). Rats fed on HF diet showed higher palmitic acid and lower stearic acid levels as compared to PO fed ones, while carnitine supplementation resulted in decreased palmitic acid in all the groups and a differential response for stearic acid, increase in HF and a decrease in PO fed. Linoleic and arachidonic acids increased in all dietary groups supplemented with carnitine. Regular exercise found to increase unsaturated fatty acids such as linoleic and arachidonic in all the groups. The effect of dietary fatty acids on the fatty acid composition of PE has been given in Table 4. A lower palmitic acid and higher unsaturated fatty acids such as oleic, linoleic and arachidonic acid were observed in PO fed rats as compared to HF, while exercise augmented this response. The magnitude of above response was greater in all the carnitine supplemented rats. The cardiolipin fraction from hepatic mitochondria showed higher oleic acid in HF fed rats, while higher levels of linoleic acid were observed in PO fed ones. Regular exercise induced a mild increase in the unsaturated fatty acids in both the dietary groups, while carnitine supplementation led to further increase in unsaturated fatty acids along with a significant increase in arachidonic acid in both the dietary groups (Table 4). Effect on the liver and muscle mitochondrial lipid peroxidation⎯The level of MDA was higher in Table 3⎯Effects of dietary fat, carnitine and exercise on activity of liver and muscle mitochondrial enzymes [Values are mean ± SE of 8 rats] Parameter/ Groups* Liver HFS POS HFE POE HFCRS POCRS HFCRE POCRE Muscle HFS POS HFE POE HFCRS POCRS HFCRE POCRE CPT (nmole/mg/min) NADH dehydrogense (nmole/mg/min) NADH oxidase (nmole/mg/min) Cytochrome C reductase Cytochrome oxidase (nmole/mg/min) (nmole/mg/min) 5.3 ± 0.22a 6.1 ± 0.34a 5.7 ± 0.13b 7.0 ± 0.55c 5.3 ± 0.22a 6.1 ± 0.34a 5.7 ± 0.13b 7.0 ± 0.55c 1583 ± 44a 2032 ± 117b 1637 ± 53a 2823 ± 118c 3313 ± 133d 3427 ± 127e 3268 ± 151d 3430 ± 474f 268 ± 1.3a 499 ± 2.9b 392 ± 4.9c 521 ± 3.2d 419 ± 2.2e 565 ± 6.9f 432 ± 2.0g 689 ± 3.9h 65 ± 4.5a 75 ± 1.7b 65 ± 4.4a 128 ± 13c 181 ± 28d 199 ± 16e 200 ± 6f 220 ± 18g 782 ± 7a 989 ± 7b 1100 ± 12c 1167 ± 6d 2153 ± 7e 2274 ± 18f 3146 ± 11g 3637 ± 25h 0.5 ± 0.05a 0.6 ± 0.02a 0.8 ± 0.02b 0.9 ± 0.09c 0.6 ± 0.03d 0.7 ± 0.05e 1.0 ± 0.05f 1.2 ± 0.08f 431 ± 23a 446 ± 24a 613 ± 39b 782 ± 25c 438 ± 29a 473 ± 26a 966 ± 39d 1094 ± 63e 90 ± 6a 73 ± 3b 108 ± 5c 110 ± 12c 224 ± 18d 244 ± 12e 319 ± 27f 339 ± 12f 29 ± 1.7a 30 ± 1.3a 31 ± 1.5a 46 ± 2.7b 23 ± 2.4a 27 ± 1.2a 33 ± 2.5ab 64 ± 5.0c 521 ± 30a 560 ± 32a 580 ± 47ab 678 ± 45b 605 ± 29ab 659 ± 54ab 652 ± 5ab 813 ± 44c Values in the same column that do not share the same superscript letters are significantly different at P<0.05 * Groups as in Table 1 KARANTH & JEEVARATNAM: CARNITINE, DIETARY LIPID, EXERCISE & MITOCHONDRIAL ENZYMES 507 Table 4⎯Fatty acid composition (% of total fatty acids) of hepatic mitochondrial membrane phosphotidyl choline, phosphotidyl ethanolamine and cardiolipin Groups* HFS POS HFE POE HFCRS POCRS HFCRE POCRE 16:0 46.7 33.8 35.8 31.9 39.9 30.5 29.1 21.1 Phosphotidyl choline 18:0 18:1 18:2 21.7 4.7 4.4 25.8 10.4 5.4 28.9 4.4 5.9 27.8 7.9 7.9 25.6 5.6 6.3 19.9 4.6 7.9 30.9 5.3 8.3 24.3 6.3 11.6 20:4 4.0 4.7 5.4 6.6 17.0 20.3 19.4 24.4 16:0 31.6 23.9 29.9 23.5 29.9 19.6 19.1 15.5 Phosphotidyl ethanolamine 18:0 18:1 18:2 25.3 3.5 4.9 25.9 4.4 5.5 26.5 6.4 5.6 25.9 6.6 6.6 15.8 3.9 6.9 23.8 10.0 11.4 28.9 4.1 8.9 24.9 15.1 14.5 20:4 5.3 5.4 6.8 7.0 16.7 17.9 17.9 18.9 16:0 5.8 2.6 4.8 2.6 5.7 2.2 4.7 2.1 Cardiolipin 18:0 18:1 18:2 3.5 44.5 12.1 2.3 22.4 51.7 3.9 48.9 15.2 3.6 30.0 54.2 4.6 47.5 13.3 2.7 14.1 52.5 4.9 50.2 17.4 3.4 16.8 53.6 20:4 2.9 7.5 4.2 7.8 21.5 23.5 25.5 26.6 * Groups as in Table 1 mitochondria of liver and muscle from PO-fed rats as compared to HF fed ones. Regular exercise resulted in an increase in MDA formation in liver and muscle mitochondria, more so in PO-fed rats (Table 5). Carnitine supplementation increased MDA level in hepatic mitochondria from both the dietary groups and exercise resulted in a further increase in MDA, while muscle mitochondria showed lower MDA levels except for POCRE rats. Effect on the fluidity and fragility of erythrocytes⎯The dietary fat and exercise did not have appreciable influence on the fluidity of erythrocyte membrane, while carnitine supplementation increased the fluidity significantly as compared to non-supplemented groups (Fig. 1). Fragility of erythrocytes was not affected by the dietary fat, while regular exercise decreased it significantly (Table 5). Carnitine supplementation also significantly decreased the fragility of erythrocytes as compared to respective non-carnitine fed groups. Discussion The present study deals with the effect of dietary fat, carnitine supplementation and exercise on the activities of membrane bound enzymes in mitochondria from liver and muscle of the rats. Regular moderate exercise were shown to ameliorate deleterious effects of hydrogenated fat, while carnitine supplementation further enhanced fat utilization, even in sedentary HF-fed rats10. The results of the present study showed an increased activity of oxidative enzymes in hepatic mitochondria from PO-fed rats, while regular exercise enhanced the activities of these enzymes in both liver and muscle from both the dietary groups. Carnitine supplementation led to a significant increase in activities of these enzymes in both the tissues, more so in PO-fed exercising rats. The most widely established function of carnitine is its obligate role in the transport of long chain fatty acid in to the mitochondria, where they are metabolized through β-oxidation and the Kreb’s cycle. The regular physical exercise reduced free carnitine content especially in muscle due to increased flux of fatty acid by carnitine (transport of fatty acid through inner membrane) for β-oxidation in muscle mitochondria. Earlier observation that regular physical exercise increased free fatty acids in skeletal muscle10, resulting in an increased CPT activity. The intake of PO-diet resulted in higher activity of CPT in liver and skeletal muscle as compared to HF-fed ones. The fatty acid composition of these diets might have played a role in the elevation of CPT in accordance with earlier reports4,25. Considering the effect of carnitine on CPT and other enzymes as described below in muscle confirm increased fatty acid oxidation with carnitine supplementation. Moreover, carnitine supplementation increased the free carnitine level in plasma and muscle in sedentary as well as exercising rats leading to enhanced fatty acid oxidation26. The increased fatty acid oxidation as indicated by high FFA level and low triglyceride10 in liver can also be attributed to the increased free carnitine level in carnitine supplemented rats in the present study. This finding is in agreement with the earlier observation where conditions known to stimulate fatty acid oxidation such as starvation and nephrosis were associated with higher carnitine level27, however, the nature of this association is not clear. The PO-diet containing about 13% linoleic acid has also resulted in higher activities of all the respiratory enzymes as compared to HF-fed rats as reported 508 INDIAN J EXP BIOL, MAY 2010 Table 5⎯Effects of dietary lipid, carnitine supplementation and exercise on liver and muscle mitochondrial lipid peroxidation and osmotic fragility of erythrocytes. [Values are mean ± SE of 8 rats] Groups* Lipid peroxidation (nmole/g) Liver HFS POS HFE POE HFCRS POCRS HFCRE POCRE Osmotic fragility (% hemolysis) of erythrocytes Muscle a 0.22 ± 0.011 0.28 ± 0.018ab 0.24 ± 0.010a 0.31± 0.039b 0.30 ± 0.106b 0.42 ± 0.036c 0.34 ± 0.036b 0.47 ± 0.029c 0.27 ± 0.010ad 0.31 ± 0.010ab 0.33 ± 0.013b 0.47 ± 0.037c 0.24 ± 0.013d 0.28 ± 0.006ad 0.26 ± 0.007ad 0.43 ± 0.008c 86 ± 3.4a 79 ± 3.8a 59 ± 6.6b 57 ± 5.1b 66 ± 3.4b 61 ± 2.8b 51 ± 3.9b 41 ± 5.4c Values in the same column that do not share the same superscript letters are significantly different at P<0.05. Groups as in Table 1 * Fig. 1⎯Effect of dietary fat, carnitine supplementation and exercise on the fluidity of erythrocyte membrane through dimer:monomer (D:M) ratio of pyrene incorporated erythrocyte ghosts. [Values are mean ± SE of 6 experiments. Values having different alphabets (m.n.o.p.q and r) comparing various treatments are significantly different among various groups at P<0.05.] previously in rats fed on linoleic acid compared to beef tallow diet28. Regular physical exercise resulted in a significantly increased activity of all respiratory chain enzymes in liver as well as muscle in accordance with the earlier reports7,29. Exercise has also shown to enhance mitochondria electron transport chain activity in older human skeletal muscle30. Carnitine supplementation with these diets has further enhanced the respiratory enzymes activities along with profound increase in free carnitine levels in plasma, liver and muscle. The effect of carnitine supplementation was even greater in exercising rats compared to other groups. Previously, exercise training and carnitine supplementation in diet were shown to increase carnitine levels, triglyceride utilization and endurance in rats31. Surprisingly the activities of respiratory enzymes were higher in sedentary rats fed on carnitine supplemented diet. Recently, L-carnitine supplementation and physical exercise were shown to restore age-associated decline in mitochondrial functions in the rat32. The mechanism whereby L-carnitine induces this shift in respiratory chain enzymes pattern is not known yet. In present study, the effects of dietary fat and exercise on hepatic mitochondrial fatty acid profiles were in agreement with those found by Quiles et al5, but significant increase in 20:4 level was the most interesting results obtained with respect to the effect of carnitine supplementation (Table 4). The enhanced CPT and oxidative enzymes activity may be attributed to the higher unsaturated fatty acid incorporation into the membrane5 resulting in an increased fluidity. As such we have observed an increased fluidity of the erythrocyte ghost membrane and increased lipid peroxidation in mitochondria further supports the above conclusions. However, the lipid peroxidation was reduced in muscle mitochondria of carnitine fed rats. The earlier study on carnitine as an antioxidant has shown that carnitine protects muscle from oxidative stress due to dietary lipids or exercise33. In summary, the PO diet, exercise and carnitine supplementation increases the activities of mitochondrial enzymes, probably due to the changes in mitochondrial membrane fatty acid composition and fluidity. Carnitine supplementation increases the oxidative capacity of both liver and muscle in sedentary and exercising rats by enhancing the activity of respiratory enzymes and also influences lipid profile of liver mitochondria. KARANTH & JEEVARATNAM: CARNITINE, DIETARY LIPID, EXERCISE & MITOCHONDRIAL ENZYMES Acknowledgement Financial support by Defence Research and Development Organisation in the form of Senior Research Fellowship to one of the authors, JK is acknowledged. References 1 Periago J L, Suarez M D & Pita M L, Effect of dietary olive oil, corn oil and medium chain triglyceride on the lipid composition of rat red blood cell membranes, J Nutr, 120 (1990) 986. 2 Suarez A, Ramrez M C D, Faus M J & Gil A, Dietary longchain polyunsaturated fatty acids influence tissue fatty acid composition in rats at weaning, J Nutr, 126 (1996) 887. 3 Barzantti V, Battino M, Baracca M, Cavazzoni M, Cocchi M, Noble M, Maranesi M, Turchetto E & Lenaz G, The effect of dietary lipid changes on the fatty acid composition and function of liver, heart and brain mitochondria in the rat at different ages, Brit J Nutr, 71 (1994) 193. 4 Power G W, Yaqoob P, Harvey D J, Newsholme E A & Calder P C, The effect of dietary lipid manipulation on hepatic mitochondrial phospholipid fatty acid composition and carnitine palmitoyl transeferase I activity, Biochem Mol Biol Internl, 34 (1994) 671. 5 Quiles J L, Huertas J R, Manas M, Battino M & Mataix J, Physical exercise affects the lipid profile of mitochondrial membranes in rats fed with virgin olive oil or sunflower oil, Brit J Nutr, 81 (1999) 21. 6 Quiles J L, Huertas J R, Mañas M, Ochoa J J, Battino M & Mataix J, Dietary fat type and regular exercise affect mitochondrial composition and function depending on specific tissue in the rat, J Bioenerg Biomembr, 33 (2001) 127. 7 Terblanche S E, Gohil K, Packer L, Henderson S & Brooks G A, The effects of endurance training and exhaustive exercise on mitochondrial enzymes in tissues of the rat (Rattus norvegicus), Comp Biochem Physiol, 128A (2001) 889. 8 Arenas J, Huertas R, Campos Y, Diaz A T, Villalon J M & Vilas E, Effects of L-carnitine on the pyruvate dehydrogenase complex and carnitine palmitoyl transferase activities in muscle of endurance athletes, FEBS Lett, 341 (1994) 91. 9 Huertas R, Campos Y, Diaz E, Esteban J, Vechietti L, Montanari G, D’Iddio S, Corsi M & Arenas J, Respiratory chain enzymes in muscle of endurance athletes: Effect of Lcarnitine, Biochem Biophys Res Commun, 188 (1992) 102. 10 Jyothsna Karanth & Jeevaratnam K, Effect of dietary lipid, carnitine and exercise on lipid profile in rat blood, liver and muscle, Indian J Exp Biol, 47 (2009) 748. 11 Jyothsna Karanth, Kumar R & Jeevaratnam K, Response of antioxidant system against dietary fat and exercise on rat tissues, Indian J Physiol Pharmacol, 48 (2004) 446. 12 Wieland O, Duefall T & Paetzke-Brummer I, Free and esterified carntine, Colorimetric method, in: Meth Enz Anal. Vol III (Metabolites, 3) edited by H U Bergemeyer, J Bergemeyer & M Grab (VCH Publishers) 1985, 481. 13 Kaplay S S, Lack of association between human erythrocyte membrane acetylcholinesterase and osmotic fragility, Biochem Med, 19 (1978) 299. 509 14 Dodge J T, Mitchell C & Hanahan D J, The preparation and characteristics of hemoglobin- free ghosts of human erythrocytes, Arch Biochem Biophys, 100 (1963) 119. 15 Chang T, Sharpless P B, Devenport D A, Radunsky M B & Yu H, Osmotic deformation of red blood cell ghosts induced by carbohydrates, Biochem Biophys Acta, 731 (1983) 346. 16 Jeevaratnam K & Vaidyanathan C S, Acute toxicity of methyl isocyanate in rabbit: In vitro and in vivo effects on rabbit erythrocyte membrane, Arch Environ Contam Toxicol, 22 (1992) 300. 17 Lowry O H, Rosebrough N J, Farr A L & Randal R J, Protein measurement with the folin phenol reagent, J Biol Chem, 193 (1951) 265. 18 Kurup C K R, Aithal H N & Ramasarma T, Increase in hepatic mitochondria on administration of ethyl-p-chlorophenoxyisobutyrate to the rat, Biochem J, 116 (1970) 773. 19 Jeevaratnam K, Vidya S & Vaidyanathan C S, In vitro and in vivo effect of methyl isocyanate on rat liver mitochondrial respiration, Toxicol Appl Pharmacol, 117 (1992) 172. 20 Jackman M R & Willis W T, Characteristics of mitochondria isolated from type I and type IIb skeletal muscle, Am J Physiol, 270 (1996) C673. 21 Gornall A G, Bardwill G J & David M M, Determination of serum proteins by means of biuret reaction, J Biol Chem, 117 (1949) 751. 22 Bieber L L, Abraham T & Helmrath T, A rapid spectrophotometric assay for carnitine palmitoyl transferase, Anal Biochem, 50 (1972) 509. 23 Jeevaratnam K, Vidya S & Vaidyanathan C S, Role of lipid peroxidation in impairment of mitochondrial function at complex I by methyl isocyanate treatment of rats, Biochem Mol Biol Internl, 30 (1993) 411. 24 Folch J, Lees M & Stanley G H S, A simple method for the isolation and purification of total lipids from animal tissues, J Biol Chem, 226 (1957) 497. 25 Power G W & Newsholme E A, Dietary fatty acids influence the activity and metabolic control of mitochondrial carnitine palmitoyl transferase I in rat heart and skeletal muscle, J nutr, 127 (1997) 2142. 26 Bacurau R F P, Navarro F, Bassil R A, Meneguello M O, Santos R V T, Almeida A L R & Rosa F B P C, Does exercise training interfere with the effects of L- carnitine supplementation? Nutr Res, 19 (2003) 337. 27 Shurbaji A, Berglund L, Berge R K, Cederblad G & Humble E, On the interrelationship between hepatic carnitine, fatty acid oxidation and triglyceride biosynthesis in nephrosis, Lipids, 32 (1999) 547. 28 Abuirmeileh N M & Elson C E, The influence of linoleic acid intake on membrane-bound respiratory activities, Lipids, 15 (1980) 918. 29 Davies K J A, Packer A L & Brooks G A, Biochemical adaptation of mitochondria muscle and whole–animal respiration to endurance training, Arch biochem Biophys, 209 (1981) 539. 30 Menshikova E V, Ritov V B, Fairfull L, Ferrell R E, Kelly D E & Goodpaster B H, Effects of exercise on mitochondrial content and function in aging human skeletal muscle, J Gerontol, 61A (2006) 534. 31 Kim E. Park H & Cha Y S, Exercise training and supplementation with carnitine and antioxidants increases carnitine stores, triglyceride utilization, and endurance in exercising rats, J Nutr Sci Vitaminol, 50 (2004) 335. 510 INDIAN J EXP BIOL, MAY 2010 32 Bernard A, Rigault C, Mazue F, Le Borgne F & Demarquoy J, L-Carnitine supplementation and physical exercise restore age-associated decline in some mitochondrial functions in the rat, J Gerontol 63A (2008) 1027. 33 Jyothsna Karanth & Jeevaratnam K, Oxidative stress and antioxidant status in rat blood, liver and muscle: effect of dietary lipid, carnitine and exercise, Int J Vitam Nutr Res, 75 (2005) 333.