Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
J. Embryol. exp. Morph. Vol. 32, 2, pp. 445-459, 1974
Printed in Great Britain
445
Investigation on the origin of the definitive
endoderm in the rat embryo
By B. L E V A K - S V A J G E R 1 AND A. S V A J G E R 2
From the Institute of Biology, University of Zagreb
SUMMARY
Single germ layers (or combinations of two of them) were isolated from the primitive
streak and the head-fold stage rat embryos and grown for 15 days under the kidney capsule
of syngeneic adult animals. The resulting teratomas were examined histologically for the
presence of mature tissues, with special emphasis on derivatives of the primitive gut.
Ectoderm isolated together with the initial mesodermal wings at the primitive streak
stage gave rise to tissue derivatives of all three definitive germ layers. Derivatives of the
primitive gut were regularly present in these grafts. At the head-fold stage, isolated ectoderm
(including the region of the primitive streak) differentiated into ectodermal and mesodermal
derivatives only.
Endoderm isolated at the primitive streak stage did not develop when grafted and was
always completely resorbed. At the head-fold stage, however, definitive endoderm differentiated into derivatives of the primitive gut if grafted together with adjacent mesoderm.
These findings indirectly suggest the migration of prospective endodermal cells from
the primitive ectoderm, and therefore a general analogy with the course of events during
gastrulation in the chick blastoderm.
INTRODUCTION
The origin of endoderm is one of the most intricate problems in the
embryology of vertebrates (see Peter (1941) for review of older literature).
According to the classical view the endoderm of avian and mammalian embryos
arises by delamination from the epiblast ('Deuterendoderm') and therefore
differs in its origin from the 'Protendoderm' of the amphibian embryo, which
arises as a consequence of gastrulation (Peter, 1941).
During the last four decades, however, this view has been subjected to
substantial revision as far as the chick embryo is concerned. Various experimental techniques, including grafting of dissected parts of the blastoderm
on to the chorioallantoic membrane (Rudnick, 1935; Hunt, 1937a) and labelling of selected areas of the epiblast with vital dyes or carbon particles (Hunt,
19376; Fraser, 1954; Vakaet, 1962; Spratt & Haas, 1965) or with tritiated
1
Author's address: Institute of Biology, Faculty of Medicine, 41001 Zagreb, P.O. Box
166, Yugoslavia.
2
Author's address: Institute of Histology and Embryology, Faculty of Medicine, 41001
Zagreb, P.O. Box 166, Yugoslavia.
446
B. L E V A K - S V A J G E R A N D A. S V A J G E R
thymidine (Nicolet, 1965; Modak, 1966; Rosenquist, 1966; Nicolet, 1967,
1970; Gallera, 1972) have shown that prospective endodermal cells are localized
in the epiblast at the primitive streak stage. These results indicate that the
definitive endoderm of the avian embryo arises by morphogenetic movements
of superficial cells, and therefore, contrary to the classical view, does not
differ essentially in its origin from the endoderm of the amphibian embryo.
There are few experimental data on the origin of the definitive endoderm
in mammalian embryos. Grobstein's experiments (1952) indicated that also
in the mammalian embryo the primitive ectoderm contains prospective endodermal cells. He mechanically removed the outer cell layer (primitive endoderm) of mouse embryonic shields prior to the appearance of the head process
(various egg-cylinder lengths and developmental stages), and grafted precultured clusters of endoderm-deprived shields into the anterior chamber of
the eye of adult mice for 30 days. Gut epithelium was present in many of the
recovered grafts. The author concluded that 'the inner cell layer, or primitive
ectoderm, of the mouse embryonic shield cannot be regarded as a germ layer
with capacities sharply limited to ectodermal differentiation'.
Using a more reliable method for the separation of germ layers by pretreatment with proteolytic enzymes (Levak-Svajger, Svajger & Skreb, 1969), we
recently repeated Grobstein's experiment in a more precise form on rats
(Levak-Svajger & Svajger, 1971). In renal homografts of ectoderm isolated
at the pre-primitive streak stage (two-layered embryonic shield), different
mature tissues, derivatives of all three germ layers, developed in most recovered
grafts. Derivatives of the primitive gut were present in 14 out of 15 grafts.
These results have confirmed Grobstein's conclusion.
The present paper deals with the differentiative capacities of rat embryo
germ layers isolated during the next developmental stages : the primitive streak
and the head-fold stage. It was hoped to obtain data on the origin of the
definitive endoderm in the rat embryo.
MATERIAL AND METHODS
The inbred Fischer strain of albino rat was used in the experiments. Gestation
was considered to have begun early in the morning when sperm was found
in the vaginal smear. Twenty-four hours later the eggs were considered to be
1 day old.
Pregnant females were anaesthetized with ether on gestation days 8^ (at
3 p.m. on the 9th day) and 9 (at 8 a.m. on the 10th day). The entire eggcylinders (embryonic shield + extra-embryonic membranes + ectoplacental cone)
were isolated with watchmaker's forceps in sterile Tyrode's saline. Only eggcylinders belonging to stages 13 and 15 (Nicholas, 1962) were selected for
the experiment. These stages correspond to stages 12 and 14 of Witschi (see
New, 1966).
447
Origin of endoderm in rat embryos
Extra-embryonic part of
the egg-cylinder
_Cut through
the endoderm
Ectoderm
Endoderm
The extra-embryonic
part is discarded
Cut through the
entire egg-cylinder
Primitive streak
Graft series I,
ectoderm+mesoderm
Graft series II,
endoderm
FIGURE 1
Manipulation of the primitive streak stage egg-cylinder
after treatment with enzymes.
Treatment of stage 13 egg-cylinders (primitive streak) (Fig. 1)
The egg-cylinders were treated as in our previous experiments with stage 12
egg-cylinders (Levak-Svajger & Svajger, 1971). The ectoplacental cone and
the Reichert's membrane were removed. The embryonic shields with their
extra-embryonic parts were treated with a mixture of 0-5 % trypsin (crystallized,
lyophilized, Worthington) and 2-5% pancreatin (Difco) in calcium- and
magnesium-free Tyrode's saline at 4 °C (Levak-Svajger et al. 1969). After
20-30 min enzymic reactions were stopped with Tyrode's saline to which a
few drops of rat serum were added. The egg-cylinders were then twice rinsed
in pure saline and transferred to saline-rat serum mixture in which they were
further manipulated with electrolytically sharpened and polished tungsten
needles. First, a circular cut was made through the outer cell layer (endoderm)
at the level of the amnion. The embryonic endoderm was then everted over the
29
E M B 32
448
B. L E V A K - S V A J G E R A N D A. S V A J G E R
Extra-embryonic
part of the
egg-cylinder
(discarded)
2nd cut
Primitive
streak
Primitive
streak
Neural
groove
Hensen's node
Ectoderm
Mesoderm
Endoderm
Discarded
Graft series III
ectoderm
Graft series IV,
endoderm+mesoderm
FIGURE
2
Manipulation of the head-fold-stage egg-cylinder. (A-C) External
aspect. (D-F) Cross-section. (See text.)
Origin of endoderm in rat embryos
449
underlying layers of the embryonic shield. The denuded, cup-shaped inner
cell layer (ectoderm) was partially (about one-third of the surface) covered
with mesoderm expanding bilaterally from the primitive streak. The embryonic
ectoderm with the incomplete mesoderm was separated from the extra-embryonic
part of the egg-cylinder by a transverse cut and grafted under the kidney
capsule (series I). The endoderm was grafted as a separate series (II).
Treatment of stage 15 egg-cylinders {head-fold) (Fig. 2)
These embryos were treated in a somewhat different way from the previous
ones. After isolation of the egg-cylinders from the uterus, the ectoplacental
cone and Reichert's membrane were removed. The embryonic shield was
separated from the extra-embryonic part of the egg-cylinder by an oblique cut
immediately below the insertion of the amnion. A second oblique cut was made
through the entire wall of the embryonic shield, running from its free edge
(near the posterior end of the primitive streak) to its top-end (the region of
Hensen's node). The originally cup-shaped embryo was thus transformed into
an almost flat shield. Now the primitive streak and the neural groove were
not continuous in line as they are in the chick blastoderm, but diverged from
Hensen's node forming a blunt angle. The embryonic shields were then treated
with enzymes for 30 min and rinsed in saline-rat serum mixture as described
before. During this treatment the ectoderm had detached spontaneously from
the underlying mesoderm, except for the region of the primitive streak, where
a complete separation was achieved with needles. The endoderm remained
loosely adherent to the overlying mesoderm. Two grafts were prepared from
each shield:
(a) The entire ectoderm including the region of the primitive streak (series III).
The presence of the primitive streak, which contained cells with developmental
capacities other than ectodermal, was presumed to assure the continuity of
cell migration after grafting.
(b) The incomplete endoderm + mesoderm (series IV). The caudal (posterior)
part of the endoderm-mesoderm combination, immediately underlying the
primitive streak in situ, was cut off and discarded. The combined graft was
thus restricted to the region in which there was no continuity with cells
migrating through the primitive streak.
All operations were performed under a dissecting microscope, using magnifications of x 25-100.
Grafting procedure and examination of grafts
Each specimen prepared as described above was transferred by means of
a braking pipette under the capsule of the right kidney of an adult male rat
of the same strain, about 3 months old. Fifteen days following transfer the
recipient animals were killed. The grafts were isolated from the host tissue,
fixed in Zenker's fluid, embedded in paraffin wax, serially sectioned and stained
29-2
450
B. L E V A K - S V A J G E R A N D A. S V A J G E R
Table 1. Incidence of mature tissues in renal homografts
of rat embryo germ layers
Series
I
...
II
Stage of the embryo (Nicholas)
IV
Ill
A.
A
N
r~
13
(primitive streak)
f
\
15
(head-fold)
A
Type of graft
...
No. transferred
No. differentiated
Skin
Neural tissue
Derivatives of the primitive gut
Pharynx oesophagus
Respiratory tube
Glands
Thymus
Thyroid
Intestine
Total
Adipose tissue
White
Brown
Cartilage
Bone
Muscle
Smooth
Skeletal
Heart
Ecto + meso
Endo
17
15
—
15
17
—
—
—
11
11
11
11
11
10
—
—
13
15
14
2
3
8
15
—
—
—
—
—
—
—
—
1
—
—
—
2
2
5
10
8
2
2
—
10
3
3
15
6
—
—
—
—
10
6
11
9
—
10
10
8
15
5
11
—
—
—
3
10
—
4
6
4
Ecto
Endo + meso
with hemalum and eosin. The sections were examined histologically for the
presence of mature tissues, with special emphasis on derivatives of primitive
gut.
RESULTS
The results of histological examination of grafts belonging to all four
experimental series are listed in Table 1. The incidence of survival and
differentiation into mature tissues was very high in series I, III and IV. On the
contrary, all grafts of isolated endoderm (series II) were completely resorbed.
All successful grafts were tumorous masses of different sizes, composed of
mature tissues, with no signs of any immunological reaction. The general
appearance of these grafts did not differ from those of whole embryonic
shields transferred under the kidney capsule (Skreb, Svajger & Levak-Svajger,
1971). With only a few exceptions, the grafts were loosely adherent to both
Origin of endoderm in rat embryos
451
the kidney capsule and the kidney parenchyme. They were well vascularized
from both sides.
With regard to their histological structure, the grafts were typical teratomas.
Neural tissue, skeletal muscle, bone and cartilage were present in chaotic
mixtures. On the other hand, epithelial tissues most frequently appeared in
organotypic association with tissues of mesenchymal origin. Oesophageal and
intestinal epithelia formed tubes or cysts surrounded by characteristic muscular
layers, which sometimes contained small intra-mural ganglia. Ciliated columnar
epithelium of the respiratory tube was closely associated with cartilage, which
often formed a complete or incomplete annulus. Different epithelial appendages
(hairs, sebaceous glands, serous or mucous glands, thyroid, thymus) were
often found in continuity or in close proximity to their original surface epithelium. Cysts of different sizes, filled with an amorphous, probably mucous
material, were a common finding. They were lined by simple, flattened epithelium, usually indefinable as to its origin.
The pattern of histological differentiation was different in each particular
series of grafts. Only tissues of mesodermal origin were present in all three
successful series of grafts (I, III and IV), and cartilage was the most constant
representative.
With respect to the differentiation of ectodermal and endodermal tissues,
the most prominent features were the following :
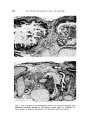
(a) Series I. Neural tissue and derivatives of the primitive gut (including
foregut, midgut and hindgut) developed in all grafts (Figs. 3, 4). Epidermis
and its derivatives were absent.
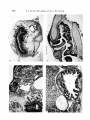
(b) Series III. Skin and neural tissue developed in all grafts (Figs. 5, 6).
A rudimentary gut was present in only 2 out of 11 grafts.
(c) Series IV. The grafts completely lacked ectodermal derivatives. Endodermal epithelia were present in all grafts, but they were restricted to derivatives
of the foregut (Figs. 7, 8).
DISCUSSION
Reliability of methods
The method which we have proposed for the separation of germ layers was
a simple modification of the common enzymic technique for ' splitting off' of
epithelia from the underlying mesenchyme at the level of the basement membrane (Levak-Svajger et al. 1969). A basement membrane exists between
ectoderm and endoderm and between each of these and mesoderm in the
two-layered and the three-layered mouse embryonic shield respectively (Pierce,
1966). Treatment of embryonic shields with enzymes results in a decrease of
mutual adhesiveness of germ layers to such a degree that subsequent mechanical
separation by needles can be accomplished with relative ease.
The treatment with enzymes of head-fold egg-cylinders (stage 15) which
452
B. LEVAK-SVAJGER AND A. SVAJGER
?ÄM';';;
Figs. 3 and 4. Details of the histological structure of teratomas obtained from
ectoderm + mesoderm isolated at the primitive streak stage, ca, cartilage; he,
heart muscle; in, intestine; ph, pharynx; re, respiratory tube; thy, thyroid.
Origin of endoderm in rat embryos
453
have been transformed into flat shields brings about a spontaneous and 'clean'
detachment of ectoderm from the underlying mesoderm. This is most probably
due to the cell-intrinsic deformation tendency of neurectoderm which is involved
in neural groove formation at this stage.
Prior to neurulation (stage 13), however, a spontaneous deformation and
detachment of ectoderm did not occur and therefore no advantage could be
taken of transforming the egg-cylinder into a flat shield before treating it
with enzymes. The embryonic endoderm is very thin and transparent. Any
defect in it which could arise during mechanical manipulation appears as a
large hole. Any eventual 'contamination' of endoderm with adherent mesodermal cells could be easily detected by careful inspection during manipulation.
It has been previously demonstrated that, after, transfer under the kidney
capsule, the rat embryonic shields are in an environment which is suitable
for their further growth and histological differentiation (Skreb et al. 1971).
The great diversity of differentiation and organ-specific association of tissues
in the resulting teratomas is unlikely to have been non-specifically induced
by the host tissue. The results of the present experiment suggest that migration
of cells through the primitive streak and Hensen's node continues after transfer
of the ectoderm under the kidney capsule. However, because these cell movements occur in an environment which is very different spatially, cell groups
establish contacts and inductive interactions by chance rather than by topographically and chronologically ordered displacements.
In our present experiment, different tissue derivatives, representative of
particular definitive germ layers, frequently developed in grafts. Their presence
varied regularly in relation to the original germ-layer composition of the
graft. It is therefore very probable that the final histological composition of
the grafts could be regarded as an expression of the developmental capacities
of the embryonic material at the moment of transfer.
In comparison with the chick blastoderm explanted in vitro, the rat embryonic
shield transplanted under the kidney capsule is subjected to much greater
environmental changes, and therefore greater disturbance of normal development is to be expected. However, when areas of the chick neurectoderm and
mesoderm were cultivated as intra-coelomic grafts (i.e. in environmental conditions similar to those in rat renal homografts), they formed neural structures
showing regional characteristics which were generally in accordance with the
prospective significance of the excised areas (Hara, 1961). Bearing this in
mind, one may postulate that the developmental capacities realized in renal
homografts of isolated rat embryo germ layers correspond at least roughly
to their developmental significance in normal intra-uterine development.
Developmental capacities of the ectoderm at the primitive streak stage
At this stage ectoderm was isolated and grafted together with incipient
mesodermal wings. A complete removal of this 'pioneer' mesoderm is not
454
B. LEVAK-SVAJGER AND A. SVAJGER
Origin of endoderm in rat embryos
455
possible. Moreover, the removed mesoderm would have been replaced very
soon after grafting by newly migrated cells. At this stage migration of cells
through the primitive streak has just begun. The presence of the primitive
streak and the incipient mesodermal wings explains the differentiation of
mesodermal derivatives in these grafts.
It remains to explain the presence of the endodermal epithelial formations
(gut and its derivatives) in these grafts, as well as in those of isolated preprimitive streak ectoderm (Levak-Svajger & Svajger, 1971). The alternatives put
forward by Grobstein (1952) are : (a) the presence of prospective endodermal cells
within the ectoderm, (b) the regenerative capacity of the ectoderm (see also
Rudnick, 1935), and (c) the full plasticity of the ectoderm at this stage. There
is no decisive support for any of these alternatives. If we favour the first one,
i.e. an ectodermal origin of the definitive embryonic endoderm, this is mainly
by analogy with the mechanism of endoderm formation in the chick blastoderm
as demonstrated by Nicolet (1967, 1970). Experimental evidence already
suggests that primary induction involves the same type of embryonic tissue
interaction in birds and in mammals (Waddington, 1936; Törö, 1938). On
the other hand, it is possible that the mechanism of the germ-layer formation
could be modified in some details as an adaptation to the entypic development
of the mouse and rat embryo (Bonnevie, 1950).
The idea of the ectodermal contribution to the definitive endoderm in the
mouse and rat embryo is not new. Sobotta (1903) asserts that the mouse
embryonic ectoderm before gastrulation 'nicht allein Ektoderm ist sondern
dass es Teilen aller drei Keimblätter den Ursprung gibt, dem gesamten
Ektoderm und Mesoderm und Teilen des Entoderms'. It was suggested that
the head process contributed to the lining of the gut as it seemed to be continuous with the endoderm on histological sections through the mouse
embryonic shield (Jolly & Férester-Tadié, 1936; Snell & Stevens, 1966). Consistent with this view is the designation of the ectoderm before gastrulation
as the primary (Widakowich, 1910; Huber, 1915) or primitive ectoderm
(Grobstein, 1952) in contrast to the secondary or definitive ectoderm after
gastrulation.
Some previous results from our laboratory strongly suggest a remarkable
change in the ultrastructural and enzymic pattern of the rat and mouse
embryonic endodermal cells after mesoderm formation. At the two-layered
stage, both embryonic and extra-embryonic endoderm are composed of cuboidal
cells with abundant microvilli, pinocytotic vacuoles and lysosomes, and a high
FIGURES 5-8
Details of the histological structure of teratomas obtained from ectoderm (Figs.
5, 6) and endoderm + mesoderm (Figs. 7, 8) isolated at the head-fold stage, b.a.t.,
Brown adipose tissue; br, brain; ca, cartilage; chp, choroid plexus; oe, oesophagus;
re, respiratory tube; s, skin.
456
B. LEVAK-SVAJGER AND A. SVAJGER
activity of acid phosphatase and nonspecific esterase. After mesoderm formation
these characteristics become restricted to the extra-embryonic endoderm, whose
cells also show the longest phase of DNA synthesis. The cells of the embryonic
endoderm are at this stage flattened, depleted of microvilli, vacuoles and
lysosomes, and devoid of hydrolytic enzyme activity (Rodé, Damjanov &
Skreb, 1968; Solter, Damjanov & Skreb, 1970,1973; Solter, Skreb & Damjanov,
1971). These findings suggest a highly developed absorptive (i.e. nutritive)
function of the entire endoderm before gastrulation. After gastrulation, the
differentiated absorptive cells are replaced by (or have dedifferentiated into?)
the flattened cells destined to differentiate into gut epithelium.
The present results indicate that the histogenetic capacity of ectoderm with
incipient mesoderm at the primitive streak stage does not differ significantly
from the capacity of ectoderm alone, isolated at the pre-primitive streak stage
(Levak-Svajger & Svajger, 1971). The prospective endodermal cells supposedly
either still reside within the ectodermal layer, or are just leaving it through the
primitive streak.
The surprising fact that skin did not develop in grafts of ectoderm taken
from the pre-primitive streak and the primitive streak stages (Levak-Svajger
& Svajger, 1971, and the present experiment) is not relevant to the problem
of endoderm formation and therefore will be discussed elsewhere.
Developmental capacities of the ectoderm
at the head-fold stage
It has been shown that in the chick blastoderm, the migration of the
prospective endodermal cells starts very early in development (at the short
primitive streak stage) and occurs in the anterior part of the primitive streak.
As soon as the head process appears, this region gives rise to somite mesoderm
only (Nicolet, 1967). Similarly, in our experiment ectoderm isolated and
grafted at the head-fold stage gives rise only to mesodermal and ectodermal
tissues. The presence of gut in 2 out of 15 grafts may be ascribed either to the
presence of some residual prospective endodermal cells within the primitive
streak at the moment of grafting, or to the unsuccessful removal of the endoderm in these specimens.
Developmental capacities o f the endoderm
The failure of endoderm isolated at the pre-primitive streak (Levak-Svajger
& Svajger, 1971) and at the primitive streak stage (present experiment) to
differentiate as renal homografts is not surprising. Even at the head-fold stage
isolated endoderm does not differentiate but is completely resorbed after
transfer under the kidney capsule (Levak-Svajger et al. 1969). At the same
developmental stage, endoderm differentiates into typical endodermal epithelia
if grafted together with adherent mesoderm (present experiment). Obviously
Origin of endoderm in rat embryos
457
an appropriate epithelio-mesenchymal interaction is needed for histological
differentiation of endodermal organs.
Unfortunately, any information on the developmental capacities of the
primary (pre-gastrulation) embryonic endoderm is lacking. This problem might
be approached by grafting pre-gastrulation or early gastrulation endoderm
in combination with post-gastrulation mesoderm. But this could not be regarded
as a decisive experiment, for it has been shown that yolk-sac endoderm of the
13- to 14-day-old rat embryo could differentiate into intestine-like structures
and other mature tissues when grafted into the mother's omentum (Payne &
Payne, 1961).
The absence of intestine in series IV grafts could be explained by the removal
of the posterior part of the endoderm (underlying the primitive streak) prior
to transplantation.
Conclusion: The possible mechanism of formation
of the definitive endoderm.
Neither the present investigation nor the previous one (Levak-Svajger &
Svajger, 1971) have provided direct evidence of an ectodermal origin of the
definitive endoderm in the rat embryo. However, the results of our experiments
strongly suggest an analogy with the course of events during gastrulation in
the chick blastoderm. In the pre-primitive streak stage, the inner cell layer
(the primary or primitive ectoderm) seems to contain prospective cells of all
three definitive germ layers. During the primitive streak stage, the prospective
endodermal and mesodermal cells migrate from the primary ectoderm and
form the definitive endoderm (or at least a part of it) and mesoderm. At the
head-fold stage the ectoderm already lacks prospective endodermal cells, but
it still contains some prospective mesodermal cells, which are able to leave
their original position and to differentiate into mesodermal tissues. At this
stage, endoderm is already formed as a definitive germ layer able to differentiate
into gut and its derivatives if combined with mesoderm. The ectoderm as a
whole does not become the definitive germ layer until after the end of
migration of the mesodermal cells through the primitive streak. This is in
agreement with the claim that ' it is only legitimate to speak of the separate
germ layers when their segregation from one another is complete' (De Beer,
1958).
ZUSAMMENFASSUNG
Isolierte Keimblätter (oder Kombinationen zweier von ihnen) von Rattenembryonen auf
den Stadien des Primitivstreifens und des Kopffortsatzes wurden 15 Tage unter der Nierenkapsel erwachsener syngegetischer Tiere gezüchtet. Die resultierenden Teratome wurden
histologisch untersucht, mit besonderer Berücksichtigung der reifen Geweben endodermaler
Herkunft.
Das auf dem Stadium des Primitivstreifens zusammen mit der Anlage des Mesoderms isolierte Ektoderm differenzierte sich in die Derivate aller dreien Keimblätter. Die
458
B. LEVAK-SVAJGER AND A. SVAJGER
verschiedenen Abschnitte des Primitivdarms waren in diesen Transplantaten ein regelmässiger
Befund.
Auf dem Stadium des Kopffortsatzes, dagegen, differenzierte sich das Ektoderm nur in
die ektodermalen und mesodermalen Geweben.
Das auf dem Stadium des Primitivstreifens isolierte Endoderm hat sich unter der Nierenkapsel überhaupt nicht weiterentwickelt. Alle Transplantate dieser Serie wurden spurlos
resorbiert. Wenn das Endoderm auf dem Stadium des Kopffortsatzes entnommen wurde,
differenzierte es sich dagegen in typische Derivate des Primitivdarms, wenn es zusammen
mit dem anliegenden Mesoderm transplantiert wurde.
Diese Befunde sprechen für die Annahme, dass das definitive Endoderm im Rattenembryo
wenigstens teilweise durch die Immigration von Zellen aus dem primitiven (primären)
Ektoderm entsteht. Dementsprechend sollte eine allgemeine Analogie mit der Entstehungsweise des definitiven Endoderm während der Gastrulation des Hühnerembryos bestehen.
We are indebted to Prof. Dr N. Skreb and Dr D. A. T. New for their valuable criticism.
The skilful technical assistance of Miss BurÜica Cesar is also acknowledged. This investigation was supported by the grant iv/3 from the Research Foundation of S.R. Croatia, and
in part by NIH PL 480 Agreement No. 02-038-1.
REFERENCES
BONNEVIE, K. (1950) New facts on mesoderm formation and proamnion derivatives in the
normal mouse embryo. /. Morph. 86, 495-545.
DE BEER, SIR G. (1958). Embryos and Ancestors, 3rd ed. Oxford: Oxford University Press.
FRASER, R. C. (1954). Studies on the hypoblast of the young chick embryo. J. exp. Zool.
126, 349-399.
GALLERA, J. (1972). Alteration of the prospective fate and the inductive power of the
definitive streak node in the chick. Experientia 28, 1217-1218.
GROBSTEIN, C. (1952). Intra-ocular growth and differentiation of clusters of mouse embryonic
shields cultured with and without primitive endoderm and in the presence of possible
inductors. /. exp. Zool. 119, 355-380.
HARA, K. (1961). Regional neural differentiation induced by pre-chordal and presumptive
chorda! mesoderm in the chick embryo. Thesis, University of Utrecht.
HUBER, G. C. (1915). The development of the albino rat, Mus norvegicus albums. I. From
the pronuclear stage to the stage of mesoderm anläge; end of the first to the end of the
ninth day. /. Morph. 26, 247-358.
HUNT, T. E. (1937a). The development of gut and its derivatives from the mesectoderm
and mesentoderm of early chick blastoderms. Anat. Rec. 68, 349-369.
HUNT, T. E. (19376). The origin of entodermal cells from the primitive streak of the chick
embryo. Anat. Rec. 68, 449-459.
JOLLY, J. & FÉRESTER-TADIÉ, M. (1936). Recherches sur l'œuf du rat et de la souris. Archs
Anat. microsc. 32, 323-390.
LEVAK-SVAJGER, B. & SVAJGER, A. (1971). Differentiation of endodermal tissues in homografts of primitive ectoderm from two-layered rat embryonic shields. Experientia 27,
683-684.
LEVAK-SVAJGER, SVAJGER, A. & SKREB, N. (1969). Separation of germ layers in presomite rat
embryos. Experientia 25, 1311-1312.
MODAK, S. P. (1966). Analyse expérimentale de l'origine de l'endoblaste embryonnaire chez
les oiseaux. Revue suisse Zool. 73, 877-908.
NEW, D. A. T. (1966). The Culture of Vertebrate Embryos. London: Logos Press.
NICHOLAS, J. S. (1962). Experimental methods and rat embryos. In The Rat in Laboratory
Investigation (ed. E. J. Parris & J. A. Griffith). New York: Hafner.
NICOLET, G. (1965). Étude autoradiographique de la destination des cellules invaginées au
niveau du nœud de Hensen de la ligne primitive achevée de l'embryon de poulet. Acta
Embryol. Morph, exp. 8, 213-220.
Origin of endoderm in rat embryos
459
NICOLET, G. (1967). La chronologie d'invagination chez le poulet: étude à l'aide de la
thymidine tritiée. Experientia 23, 576-577.
NICOLET, G. (1970). Analyse autoradiographique de la localisation des différentes ébauches
présomptives dans la ligne primitive de l'embryon de poulet. J. Embryol. exp. Morph.
23, 79-108.
PAYNE, J. M. & PAYNE, S. (1961). Placental grafts in rats. J. Embryo!, exp. Morph. 9, 106116.
PETER, K. (1941). Die Genese des Entoderms bei den Wirbeltieren. Ergebn. Anat. EntwGesch.
33, 285-369.
PIERCE, G. B. (1966). The development of basement membranes of the mouse embryo.
Devi Biol. 13,231-249.
RODÉ, B., DAMJANOV, I. & SKREB, N. (1968). Distribution of acid and alkaline phosphatase
activity in early stages of rat embryos. Bull, scient. Cons. Acads RPF Yougosl. A 13, 304.
ROSENQUIST, G. C. (1966). A radioautographic study of labelled grafts in the chick blastoderm. Development from primitive streak stages to stage 12. Contr. Embryol. 38, 71-110.
RUDNICK, D. (1935). Regional restriction of potencies in the chick during embryogenesis.
J. exp. Zool. 71, 83-99.
§KREB, N., SVAJGER, A. & LEVAK-SVAJGER, B. (1971). Growth and differentiation of rat
egg-cylinders under the kidney capsule. / . Embryol. exp. Morph. 25, 47-56.
SNELL, G. D. & STEVENS, L. C. (1966). Early Embryology. In Biology of the Laboratory
Mouse (ed. E. L. Green), 2nd ed. New York: McGraw-Hill.
SOBOTTA, J. (1903). Die Entwicklung des Eies der Maus vom Schlüsse der Furchungsperiode
bis zum Auftreten der Amniosfalten. Arch, mikrosk. Anat. 61, 274-330.
SOLTER, D., DAMJANOV, I. & SKREB, N. (1970). Ultrastructure of mouse egg-cylinder.
Z. Anat. EntwGesch. 132, 291-298.
SOLTER, D., DAMJANOV, I. & §KREB, N. (1973). Distribution of hydrolytic enzymes in early
rat and mouse embryos- a reappraisal. Z. Anat. EntwGesch. 139, 119-126.
SOLTER, D., SKREB, N. & DAMJANOV, 1. (1971). Cell cycle in the mouse egg-cylinder. Expl
Cell Res. 64, 331-334.
SPRATT, N. T. & HAAS, H. (1965). Germ layer formation and the role of the primitive
streak in the chick. I. Basic architecture and morphogenetic tissue movements. J. exp.
Zool. 158, 9-38.
TöRÖ, E. (1938). The homeogenetic induction of neural folds in rat embryos. / . exp. Zool.
79, 213-236.
VAKAET, L. (1962). Some new data concerning the formation of the definitive endoblast in
the chick embryo. / . Embryol. exp. Morph. 10, 38-57.
WADDINGTON, C. H. (1936). Organizers in mammalian development. Nature, Lond. 138,
125.
WIDAKOWICH, V. (1910). Über die erste Bildung der Körperform bei Entypie des Keimes.
Beiträge zur Entwicklungsgeschichte der Ratte. Z. wiss. Zool. 94, 240-298.
{Received
18 December
1973, revised 4 March
1974)