Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
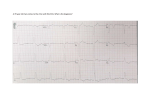
Clinical research European Heart Journal (2007) 28, 2472–2478 doi:10.1093/eurheartj/ehm339 Arrhythmia/electrophysiology Pacemaker current (If) in the human sinoatrial node Arie O. Verkerk1, Ronald Wilders1, Marcel M.G.J. van Borren1, Ron J.G. Peters2, Eli Broekhuis3, Kayan Lam3, Ruben Coronel1, Jacques M.T. de Bakker1,4, and Hanno L. Tan1,2* 1 Heart Failure Research Center, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands; Department of Cardiology, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands; 3Department of Cardiothoracic Surgery, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands; and 4 Interuniversity Cardiology Institute the Netherlands, Utrecht, The Netherlands 2 Received 15 February 2007; revised 1 June 2007; accepted 17 July 2007; online publish-ahead-of-print 6 September 2007 KEYWORDS Electrophysiology; Ion channels; Pacing; Sinoatrial node; Action potentials Aims Animal studies revealed that the hyperpolarization-activated pacemaker current, If, contributes to action potential (AP) generation in sinoatrial node (SAN) and significantly determines heart rate. If is becoming a novel therapy target to modulate heart rate. Yet, no studies have demonstrated that If is functionally present and contributes to pacemaking in human SAN. We aimed to study If properties in human SAN. Methods and results In a patient undergoing SAN excision, we identified SAN using epicardial activation mapping. From here, we isolated myocytes and recorded APs and If using patch-clamp techniques. Pacemaker cells generated spontaneous APs (cycle length 828 + 15 ms) following slow diastolic depolarization, maximal diastolic potential 261.7 + 4.3 mV, and maximal AP upstroke velocity 4.6 + 1.2 V/s. They exhibited an hyperpolarization-activated inward current, blocked by external Csþ (2 mmol/L), characterizing it as If. Fully-activated conductance was 75.2 + 3.8 pS/pF, reversal potential 222.1 + 2.4 mV, and half-maximal activation voltage and slope factor of steady-state activation 296.9 + 2.7 and 28.8 + 0.5 mV. Activation time constant ranged from 350 ms (2130 mV) to 1 s (2100 mV), deactivation time constant 156 + 45 ms (240 mV). The role of If in pacemaker activity was demonstrated by slowing of pacemaker cell diastolic depolarization and beating rate by Csþ. Conclusion If is functionally expressed in human SAN and probably contributes to pacemaking in human SAN. Introduction Cardiac pacemaking is a basic physiological function required to match cardiac performance to metabolic demand. In the mammalian heart, it is accomplished by pacemaker cells in the sinoatrial node (SAN) region. The SAN is a complex tissue with regional differences in morphological and electrical properties.1 Animal studies have revealed that pacemaking in SAN cells follows from diastolic depolarization driven by a net inward current, which results from an interaction of multiple ion currents.1,2 Inward currents are activated during diastole: hyperpolarizationactivated pacemaker current (If),3,4 background Naþ current (Ib,Na),5 sustained inward current (Ist),6,7 T- and L-type Ca2þ currents (ICa,T and ICa,L, respectively),8 and Ca2þ-release activated Naþ–Ca2þ exchange current (INCX).9 Conversely, outward currents are deactivated: rapid delayed rectifier Kþ current (IKr) and slow delayed rectifier Kþ current (IKs).10–13 The relative contributions of these * Corresponding author: Heart Failure Research Center, Department of Clinical and Experimental Cardiology, Academic Medical Center, Meibergdreef 9, 1105 AZ Amsterdam, The Netherlands. Tel: þ31 20 5663264; fax: þ31 20 6975458. E-mail address: [email protected] currents to diastolic depolarization are a matter of debate.14–16 Still, If consistently exhibits a key role in animal studies, mediating the heart-rate modulating actions of autonomic neurotransmitters,17 and underlying heart-rate slowing by aging18 and heart failure.19,20 Human studies also show, albeit indirectly, that If is clinically relevant. Recent studies linked familial SAN disease to mutations in HCN4,21–23 a molecular component of If.24 Accordingly, If is evolving into a novel therapy target: If blocking drugs are being developed to lower heart rates and be beneficial in ischaemic heart disease,25 while If gene transfer may relieve disease-causing bradycardias.26,27 Yet, direct evidence that If is functionally present in the human SAN is still lacking, as the cellular electrophysiological properties of human SAN are virtually unexplored, given the extreme difficulty of obtaining human SAN preparations for in vitro electrophysiological studies. To date, the only study on adult human SAN utilized explanted hearts of patients in end-stage heart failure who underwent cardiac transplantation.28 However, that study was limited because, in addition to possible confounding effects of endstage heart failure, ion currents underlying SAN action potentials (APs) were not studied. In the present study, we explored the electrophysiological properties of isolated Published on behalf of the European Society of Cardiology. All rights reserved. & The Author 2007. For permissions please email: [email protected]. Pacemaker current in human sinoatrial node SAN cells of a patient without structural heart disease who underwent SAN excision, in particular, AP properties and the presence and functional role of If. Methods This study complied with institutional guidelines. The patient provided written informed consent. Epicardial mapping We used a flexible plaque electrode harbouring 64 terminals arranged in an 8-by-8 matrix at inter-electrode distances of 6.5 mm. Recordings were made with a 256-channel mapping system (BioSemi ActiveTwo, 24-bit resolution) in unipolar mode. An electrode attached to the mediastinum served as reference. Cell isolation The excised tissue was immediately submerged in Tyrode’s solution (48C) and cell isolation was started. The region including and surrounding the origin of the spontaneous rhythm during surgery was cut into cubic chunks (,1 mm3), and single cells were enzymatically isolated as described previously.29 Cellular electrophysiology Cell suspensions were placed in a recording chamber on the stage of an inverted microscope, and superfused with Tyrode’s solution, which contained 140 NaCl, 5.4 KCl, 1.8 CaCl2, 1.0 MgCl2, 5.5 glucose, 5.0 HEPES (pH 7.4 with NaOH). Membrane potentials and currents were recorded at 36 + 0.28C in the whole-cell configuration of the patch-clamp technique. The pipette solution contained (mmol/L): 125 K-gluconate, 20 KCl, 5.0 NaCl, 1.0 MgCl2, 5.0 MgATP, 10.0 HEPES (pH 7.2 with KOH). Potentials were corrected for liquid junction potential, and membrane capacitance (Cm) and series resistance (Rs) were compensated for .85%. Signals were low-pass filtered (cut-off frequency 5 kHz) and digitized at 10 kHz. If was assessed as the Csþ-sensitive, time-dependent current that activated during 2 s hyperpolarizing voltage-clamp steps from a holding potential of 240 mV (step interval: 4 s). The If activation curve was obtained by plotting normalized Csþ-sensitive tail current amplitude against potential, and characterized by fitting it to the Boltzmann equation I/Imax ¼ A/{1.0þexp[(V2V1/2)/k]} to determine the membrane potential for half-maximal activation (V1/2) and the slope factor (k). The time courses of Csþ-sensitive If activation and deactivation were fitted by the monoexponential equations I/Imax ¼ A[12exp(2t/t)] and I/Imax ¼ Aexp(2t/t), respectively, ignoring the variable initial delay in If activation.4 Statistics Results are expressed as mean + SEM. AP parameters from 10 consecutive APs were averaged. Two sets of data were considered significantly different if the probability value of the unpaired Student’s t-test was ,0.05. Results Patient A 49-year-old woman had paroxysmal tachycardias from the age of 28 years, which were usually regular and developed within minutes. There was no family history of cardiac arrhythmias and no signs of structural heart disease, as repeated echocardiography, coronary angiography, and invasive hemodynamic studies were normal. Holter recordings and ECGs revealed various tachyarrhythmias, notably atrial 2473 fibrillation, and different regular narrow-complex tachycardias with antegrade P waves (RP.PR) whose morphology was either similar or dissimilar to sinus rhythm (120–150/ min). There was an almost permanently enhanced sinus rate (average daytime rate 100–120/min, average nighttime rate 90/min), suggestive of inappropriate sinus tachycardia (IST) (Figure 1A). Invasive electrophysiological studies showed no evidence of accessory pathways or dual AV nodal physiology. Atrial tachycardias were noninducible upon programmed stimulation, burst pacing, and isoproterenol or acetylcholine infusion, precluding any opportunity for RF catheter ablation. Thus, various antiarrhythmic drugs were used, including pindolol, metoprolol, verapamil, flecainide, propafenone, and amiodarone. As these drugs were either ineffective or, in the case of b-blockers, not tolerated because of side effects, a DDD pacemaker with supraventricular antitachycardia pacing algorithms (Medtronic AT500) was implanted. However, antitachycardia pacing did not terminate the supraventricular arrhythmias, and the patient did not tolerate the frequent antitachycardia pacing episodes. After repeated burst pacing attempts to induce permanent atrial fibrillation had failed, His bundle ablation was performed. Yet, the supraventricular tachyarrhythmias caused frequent mode switches to VVI pacing that resulted in pacemaker syndrome, which the patient tolerated poorly. It became increasingly clear that the patient suffered severely from the IST episodes (palpitations both at rest and already at minimal levels of exercise). We found no evidence for extrinsic dysregulation in the vagosympathetic balance, as autonomic function tests were normal (responses upon respiratory, Valsalva, and orthostatic manoeuvres). Thus, we directed our efforts at relieving these IST episodes by the use of the putative If blocker ivabradine (10 mg bid).25 This drug initially slowed baseline heart rates and partly suppressed tachycardia episodes. However, during prolonged use, these effects waned. Subsequently, we performed two procedures to extinguish the SAN using endocardial RF catheter ablation, but were unsuccessful, presumably because of the subepicardial localization of the SAN.30 Finally, we conducted surgical SAN excision. SAN excision and postoperative follow-up The patient had discontinued all antiarrhythmic drugs for at least three half-lives before SAN excision (including amiodarone and ivabradine, which were discontinued 11 months and 8 weeks before surgery, respectively). At the onset of extracorporeal circulation, she had sinus rhythm (cycle length 655 ms) as assessed by ECG. Intraoperative epicardial mapping revealed earliest activation at the anterior aspect of the superior caval vein, 1 cm cranial from the right atrium, followed by centrifugal spread of activation (Figure 2A and B). After marking the point of earliest activation with a felt-tip pen, we resected this region and closed the defect with an autologous pericardial patch. During a 30 months’ postoperative follow-up, there was no more documentation of sinus rhythm. The patient had atrial rhythm (Figure 1B) or AV sequentially paced rhythm (not shown). She had no more sinus tachycardia episodes. However, she did remain symptomatic of atrial fibrillation/ flutter. 2474 A.O. Verkerk et al. Figure 1 (A) Inappropriate (at rest) sinus tachycardia episode before sinoatrial node excision. (B) Atrial rhythm with ventricular pacing at postoperative follow-up. Note that the P-wave shape and axis are clearly different from the sinus tachycardia episode in (A) Morphological and electrophysiological properties of single cells of the SAN region We performed enzymatic isolation of cells from the resected tissue (region of earliest activation) for single-cell studies. Consistent with previous reports, the living-cell yield was low (,5%). Pacemaker cells and atrial myocytes exhibited distinct morphologies and AP configurations (Figure 2C and D; Table 1).4 Pacemaker cells had an irregular, bent, spindle shape and a paucity of myofilaments, along with spontaneous contractions and rhythmic pacemaker activity Pacemaker current in human sinoatrial node 2475 diastolic threshold. Most AP parameters differed significantly between both cell types. Atrial myocytes had a stable resting membrane potential, whereas pacemaker cells had a less negative maximum diastolic potential (MDP) and diastolic depolarization, which resulted in pacemaker activity. The maximal AP upstroke velocity (dV/ dtmax) was typically slower in pacemaker cells than in atrial myocytes. In both cell types, APs overshot the zero potential value, but the overshoot was higher in atrial myocytes. APs of atrial myocytes repolarized earlier and faster, resulting in shorter AP duration at 20% (APD20) and 50% (APD50), but not 90% (APD90) of repolarization. The If blocker Csþ (2 mmol/L) slowed pacemaker cell activity (26% cycle length increase) by slowing diastolic depolarization (Figure 2E), leaving AP overshoot and duration unaltered. If properties in pacemaker cells Figure 2 (A) Mapping electrode array over right atrium (RA) containing 64 electrodes (dots) in an 8 8 matrix, positioned 6.5 mm apart. RV, right ventricle; RAA, right atrial auricle; LA, left atrium; LAD, left anterior descending artery; SVC, superior caval vein; IVC, inferior caval vein; PA, pulmonary artery. (B) Activation map obtained with mapping electrode. Colours indicate moment of local activation (inset); lines are 10 ms isochrones. (C, D) Photographs (top) and action potentials (bottom) of an isolated pacemaker cell (spontaneous electrical activity, C) and atrial myocyte (stimulated at 2 Hz, D). (E) Effect of Csþ on the action potential of a single pacemaker cell. (Figure 2C). Atrial myocytes were rod-shaped with a clear striation pattern (Figure 2D) and were significantly longer and wider than pacemaker cells; consequently, membrane capacitance (Cm) was larger (Table 1). Atrial myocytes were quiescent or exhibited irregular slow spontaneous contractions due to spontaneous Ca2þ release (intracellular Ca2þ was measured using Indo-1, data not shown). Spontaneously contracting atrial myocytes are a consistent finding in human atrial-cell isolations.31,32 In the present study, the irregularly beating atrial myocytes had resting membrane potentials between 210 and 240 mV. Typically, these myocytes went into contracture within 15 min after the start of superfusion and were excluded from further analysis. Figure 2C and D shows typical APs of a pacemaker cell and an atrial myocyte (note different time scales); average AP properties are summarized in Table 1. APs of atrial myocytes were elicited at 2 Hz by 3 ms current pulses 50% above Subsequent voltage-clamp experiments revealed that pacemaker cells had an inward current that activated following 2 s hyperpolarizing steps from 240 mV (Figure 3A). This hyperpolarization-activated current became larger and activated more rapidly at increasingly negative potentials. Moreover, it was strongly reduced by 2 mmol/L Csþ (Figure 3A, middle), characterizing it as If.4,33 Its properties were analysed from the Csþ-sensitive current (Figure 3A, bottom). Average maximal If conductance was 75.2 + 3.8 pS/pF (Figure 3B) and the reversal potential was 222.1 + 2.4 mV, as determined from comparison of the fully-activated current (Istep) at 2130 mV and the associated tail current (Itail) at 240 mV. Voltage-dependence of activation was analysed by plotting normalized Itail amplitude against the preceding voltage steps. Activation threshold was between 250 and 260 mV (Figure 3C), and the average half-maximal activation voltage (V1/2) and slope factor of the Boltzmann fit to the data were 296.9 + 2.7 and 28.8 + 0.5 mV, respectively (Figure 3C). Activation and deactivation time constants were obtained from monoexponential fits of the step and tail currents, respectively. Activation time constants ranged from 350 ms at 2130 mV to 1 s at 2100 mV, and deactivation time constant was 156 + 45 ms at 240 mV (Figure 3B). Discussion We provide the first demonstration that human SAN pacemaker cells functionally express If. If probably contributes to diastolic depolarization and pacemaking in human SAN, as evidenced by the effects of Csþ to block If and reduce pacemaker cell beating rates by slowing diastolic depolarization. Yet, in accordance with animal studies,4 Csþ did not abolish pacemaker activity completely, indicating that other ion currents are also involved in cardiac pacemaking. Compensatory mechanisms may attenuate the effects of If blockade. For instance, removal of depolarizing current carried by If hyperpolarizes maximum diastolic potential4 and slows diastolic depolarization rate, thereby increasing other inward currents (e.g. the background Naþ current34) and decreasing outward currents (potassium current is reduced because potassium driving force is reduced and deactivation rate is increased). 2476 A.O. Verkerk et al. Table 1 General properties of single human pacemaker and atrial cells Pacemaker cells Morphology Cell length (mm) Cell width (mm) Electrophysiology Cm (pF) Cycle length (ms) MDP (mV) DDR100 (mV/s) dV/dtmax (V/s) APA (mV) Overshoot (mV) APD20 (ms) APD50 (ms) APD90 (ms) 66.7 + 6.3 (n ¼ 4) 7.8 + 0.4 (n ¼ 4) 56.6 + 8.7 (n ¼ 4) 828 + 15 (n ¼ 3) 261.7 + 4.3 (n ¼ 3) 48.9 + 18 (n ¼ 3) 4.6 + 1.2 (n ¼ 3) 78.0 + 4.5 (n ¼ 3) 16.4 + 0.7 (n ¼ 3) 64.9 + 16.9 (n ¼ 3) 101.5 + 27.0 (n ¼ 3) 143.5 + 34.9 (n ¼ 3) Atrial cells 87.8 + 4.9 (n ¼ 4)* 16.7 + 0.9 (n ¼ 4)* 88.3 + 10.7 (n ¼ 5)* — 279.7 + 4.1 mV (n ¼ 3)* — 243 + 96 (n ¼ 3)* 124.2 + 8.0 (n ¼ 3)* 44.5 + 4.3 (n ¼ 3)* 3.1 + 1.1 (n ¼ 3)* 11.6 + 4.7 (n ¼ 3)* 92.3 + 28.2 (n ¼ 3) Data are mean + SEM; n, number of cells; Cm, membrane capacitance; MDP, maximal diastolic potential; DDR100, diastolic depolarization rate over the 100 ms time interval starting at MDP; dV/dtmax, maximal upstroke velocity; APA, action potential amplitude; APD20, APD50, and APD90, action potential duration at 20, 50, and 90% repolarization. *P , 0.05. Figure 3 Voltage- and time-dependence of the hyperpolarization-activated inward current, If, in single pacemaker cells. (A) Top: superimposed original current recordings of If elicited by various voltage steps (voltage-clamp protocol in inset). Middle: effects of external Csþ on If. Bottom: Csþ-sensitive current obtained by digital subtraction of current traces recorded in the absence and presence of external Csþ. Arrows indicate If step (Istep) and tail (Itail) current. (B) Average current–voltage relationship of the If step and tail currents (n ¼ 3). (C ) Steady-state activation curve (dashed line is Boltzmann fit to the experimental data) and time constants of (de)activation (n ¼ 3). The If activation threshold (between 260 and 250 mV) and V1/2 (296.9 + 2.7 mV) were at more hyperpolarized potentials than in a recent study in which wild-type HCN4 was heterologously expressed.35 These differences may be partly explained by the different experimental models. Moreover, our hyperpolarizing shift of If activation may be due to (1) absence of cyclic AMP from the pipette solution,36 (2) cell dialysis,35 and (3) lack of full activation of If during the 2 s voltage-clamp steps at less negative potentials. Of note, the AP properties of the single pacemaker cells studied here differed markedly from those reported previously in a study of intact human SAN preparations.28 Most notably, the single cells in our study had faster spontaneous beating rates (cycle lengths 828 vs. 2300 ms). It must be noted, however, that the reported intact SAN preparations were obtained from explanted hearts of patients Pacemaker current in human sinoatrial node in end-stage heart failure,28 although heart failure was not present in our study. It is likely that these differences in AP properties are better explained by the effects of heart failure than by the type of preparation (intact node vs. isolated cells), as intrinsic SAN rate in humans is reduced in heart failure.37 Moreover, in a rabbit model of heart failure, the intrinsic rates of single SAN cells19 and intact SAN preparations38 were reduced to the same extent. Molecular characterization of If has pointed to the hyperpolarization-activated cyclic nucleotide (HCN) gated family, which comprises four members, HCN1–4.24 All except HCN3 are present in the heart, with their expression varying somewhat among species, cardiac tissue, and age.39 In SAN of various species, HCN4 is the dominant HCN isoform, constituting 80% of the total HCN message.40,41 Our demonstration of If in human SAN pacemaker cells agrees with previously reported links between familial SAN disease and HCN4 mutations.21–23 We did not attempt molecular characterization of SAN pacemaker cells, because we allocated the entire SAN region for cell isolation and chose to focus on obtaining as many electrophysiological data as possible. At the same time, we expected that the yield of enzymatic dissociation would be low and sufficient only for cellular electrophysiological studies. Study limitations This study is based on one patient, whose SAN was excised because of tachycardias which were resistant to drug therapy and RF catheter ablation. We cannot rule out that the SAN pacemaker cell If properties found here are not fully representative of the general population. Yet, obtaining human SAN cells for patch-clamp studies is extremely difficult, as SAN excision should be conducted only in patients who are refractory to all conventional therapies, as demonstrated in the present case. Similarly, it is unlikely that SAN excision of healthy subjects will ever be conducted, and large-scale studies of fully normal human SAN cells are inconceivable. Conclusion Human SAN cells have pacemaker activity qualitatively similar to that of various species. If probably contributes to pacemaking in the human SAN. These findings provide support for novel If targeting therapies, using new drug classes or gene transfer, aimed at common diseases, such as ischaemic heart disease and sick sinus syndrome, to reduce or increase heart rate. Acknowledgements The authors thank Jan Zegers and Jan Bourier for excellent technical assistance. Conflict of interest: none declared. Funding H.L.T. was supported by the Royal Netherlands Academy of Arts and Sciences (KNAW), the Netherlands Heart Foundation (NHS2002B191, NHS2005B180), and the Bekales Foundation. 2477 References 1. Boyett MR, Honjo H, Kodama I. The sinoatrial node, a heterogeneous pacemaker structure. Cardiovasc Res 2000;47:658–687. 2. Irisawa H, Brown HF, Giles W. Cardiac pacemaking in the sinoatrial node. Physiol Rev 1993;73:197–227. 3. DiFrancesco D. Pacemaker mechanisms in cardiac tissue. Annu Rev Physiol 1993;55:455–472. 4. van Ginneken ACG, Giles W. Voltage clamp measurements of the hyperpolarization-activated inward current If in single cells from rabbit sino-atrial node. J Physiol 1991;434:57–83. 5. Hagiwara N, Irisawa H, Kasanuki H, Hosoda S. Background current in sino-atrial node cells of the rabbit heart. J Physiol 1992;448:53–72. 6. Shinagawa Y, Satoh H, Noma A. The sustained inward current and inward rectifier Kþ current in pacemaker cells dissociated from rat sinoatrial node. J Physiol 2000;523:593–605. 7. Guo J, Ono K, Noma A. A sustained inward current activated at the diastolic potential range in rabbit sino-atrial node cells. J Physiol 1995;483: 1–13. 8. Hagiwara N, Irisawa H, Kameyama M. Contribution of two types of calcium currents to the pacemaker potentials of rabbit sino-atrial node cells. J Physiol 1988;395:233–253. 9. Bogdanov KY, Vinogradova TM, Lakatta EG. Sinoatrial nodal cell ryanodine receptor and Naþ–Ca2þ exchanger: molecular partners in pacemaker regulation. Circ Res 2001;88:1254–1258. 10. Verheijck EE, van Ginneken ACG, Bourier J, Bouman LN. Effects of delayed rectifier current blockade by E-4031 on impulse generation in single sinoatrial nodal myocytes of the rabbit. Circ Res 1995;76:607–615. 11. Lei M, Cooper PJ, Camelliti P, Kohl P. Role of the 293b-sensitive, slowly activating delayed rectifier potassium current, iKs, in pacemaker activity of rabbit isolated sino-atrial node cells. Cardiovasc Res 2002;53:68–79. 12. Lei M, Brown HF. Two components of the delayed rectifier potassium current, IK, in rabbit sino-atrial node cells. Exp Physiol 1996;81:725–741. 13. Ono K, Ito H. Role of rapidly activating delayed rectifier Kþ current in sinoatrial node pacemaker activity. Am J Physiol 1995;269:H453–H462. 14. Honjo H, Inada S, Lancaster MK, Yamamoto M, Niwa R, Jones SA, Shibata N, Mitsui K, Horiuchi T, Kamiya K, Kodama I, Boyett MR. Sarcoplasmic reticulum Ca2þ release is not a dominating factor in sinoatrial node pacemaker activity. Circ Res 2003;92:e41–e44. 15. Lakatta EG, Maltsev VA, Bogdanov KY, Stern MD, Vinogradova TM. Cyclic variation of intracellular calcium: a critical factor for cardiac pacemaker cell dominance. Circ Res 2003;92:e45–e50. 16. Bers DM. The beat goes on: diastolic noise that just won’t quit. Circ Res 2006;99:921–923. 17. Brown HF, DiFrancesco D, Noble SJ. How does adrenaline accelerate the heart? Nature 1979;280:235–236. 18. Accili EA, Robinson RB, DiFrancesco D. Properties and modulation of If in newborn versus adult cardiac SA node. Am J Physiol 1997;272: H1549–H1552. 19. Verkerk AO, Wilders R, Coronel R, Ravesloot JH, Verheijck EE. Ionic remodeling of sinoatrial node cells by heart failure. Circulation 2003;108: 760–766. 20. Zicha S, Fernandez-Velasco M, Lonardo G, L’Heureux N, Nattel S. Sinus node dysfunction and hyperpolarization-activated (HCN) channel subunit remodeling in a canine heart failure model. Cardiovasc Res 2005;66:472–481. 21. Milanesi R, Baruscotti M, Gnecchi-Ruscone T, DiFrancesco D. Familial sinus bradycardia associated with a mutation in the cardiac pacemaker channel. N Engl J Med 2006;354:151–157. 22. Schulze-Bahr E, Neu A, Friederich P, Kaupp UP, Breithardt G, Pongs O, Isbrandt D. Pacemaker channel dysfunction in a patient with sinus node disease. J Clin Invest 2003;111:1537–1545. 23. Ueda K, Nakamura K, Hayashi T, Inagaki N, Takahashi M, Arimura T, Morita H, Higashiuesato Y, Hirano Y, Yasunami M, Takishita S, Yamashina A, Ohe T, Sunamori M, Hiraoka M, Kimura A. Functional characterization of a trafficking-defective HCN4 mutation, D553 N, associated with cardiac arrhythmia. J Biol Chem 2004;279:27194–27198. 24. Stieber J, Hofmann F, Ludwig A. Pacemaker channels and sinus node arrhythmia. Trends Cardiovasc Med 2004;14:23–28. 25. Borer JS. Drug insight: If inhibitors as specific heart-rate-reducing agents. Nature Clinical Practice Cardiovascular Medicine 2004;1:103–119. 26. Qu J, Plotnikov AN, Danilo P Jr, Shlapakova I, Cohen IS, Robinson RB, Rosen R. Expression and function of a biological pacemaker in canine heart. Circulation 2003;107:1106–1109. 27. Boink GJ, Verkerk AO, van Amersfoorth SC, Tasseron S, van der Meulen J, de Bakker JM, Seppen J, Tan HL. Increased spontaneous activity and 2478 28. 29. 30. 31. 32. 33. 34. autonomic modulation in lentiviral transduced rat cardiac myocytes overexpressing HCN4. Heart Rhythm 2007;4:S147. Drouin E. Electrophysiologic properties of the adult human sinus node. J Cardiovasc Electrophysiol 1997;8:254–258. Verheijck EE, Wessels A, van Ginneken ACG, Bourier J, Markman MWM, Vermeulen JLM, de Bakker JMT, Lamers WH, Opthof T, Bouman LN. Distribution of atrial and nodal cells within the rabbit sinoatrial node: models of sinoatrial transition. Circulation 1998;97:1623–1631. Koplan BA, Parkash R, Couper G, Stevenson WG. Combined epicardial-endocardial approach to ablation of inappropriate sinus tachycardia. J Cardiovasc Electrophysiol 2004;15:237–240. Hoppe UC, Beuckelmann DJ. Characterization of the hyperpolarizationactivated inward current in isolated human atrial myocytes. Cardiovasc Res 1998;38:788–801. Köster OF, Szigeti GP, Beuckelmann DJ. Characterization of a [Ca2þ]idependent current in human atrial and ventricular cardiomyocytes in the absence of Naþ and Kþ. Cardiovasc Res 1999;41:175–187. DiFrancesco D. Block and activation of the pace-maker channel in calf Purkinje fibres: effects of potassium, caesium and rubidium. J Physiol 1982;329:485–507. Noble D, Denyer JC, Brown HF, DiFrancesco D. Reciprocal role of the inward currents Ib,Na and If in controlling and stabilizing pacemaker frequency of rabbit sino-atrial node cells. Proc R Soc Lond B Biol Sci 1992; 250:199–207. A.O. Verkerk et al. 35. Michels G, Er F, Khan I, Südkamp M, Herzig S, Hoppe UC. Single-channel properties support a potential contribution of hyperpolarizationactivated cyclic nucleotide-gated channels and If to cardiac arrhythmias. Circulation 2005;111:399–404. 36. DiFrancesco D, Tortora P. Direct activation of cardiac pacemaker channels by intracellular AMP. Nature 1991;351:145–147. 37. Sanders P, Kistler PM, Morton JB, Spence SJ, Kalman JM. Remodeling of sinus node function in patients with congestive heart failure: reduction in sinus node reserve. Circulation 2004;110:897–903. 38. Opthof T, Coronel R, Rademaker HM, Vermeulen JT, Wilms-Schopman FJ, Janse MJ. Changes in sinus node function in a rabbit model of heart failure with ventricular arrhythmias and sudden death. Circulation 2000;101:2975–2980. 39. Shi W, Wymore R, Yu H, Wu J, Wymore RT, Pan Z, Robinson RB, Dixon JE, McKinnon D, Cohen IS. Distribution and prevalence of hyperpolarization-activated cation channel (HCN) mRNA expression in cardiac tissues. Circ Res 1999;85:e1–e6. 40. Ishii TM, Takano M, Xie L-H, Noma A, Ohmori H. Molecular characterization of the hyperpolarization-activated cation channel in rabbit heart sinoatrial node. J Biol Chem 1999;274:12835–12839. 41. Tellez JO, Dobrzynski H, Greener ID, Graham GM, Laing E, Honjo H, Hubbard SJ, Boyett MR, Billeter R. Differential expression of ion channel transcripts in atrial muscle and sinoatrial node in rabbit. Circ Res 2006;99:1384–1393.