Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
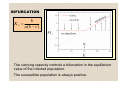
Diffusion and home ranges in mice movement Guillermo Abramson Statistical Physics Group, Centro Atómico Bariloche and CONICET Bariloche, Argentina. with L. Giuggioli and V.M. Kenkre Oh, my God, Kenkre has told everything! OUTLINE The basic model Implications of the bifurcation Lack of vertical transmission Temporal behavior Traveling waves The diffusion paradigm Analysis of actual mice transport Model of mice transport THREE FIELD OBSERVATIONS AND A SIMPLE MODEL • Strong influence by environmental conditions. • Sporadical dissapearance of the infection from a population. • Spatial segregation of infected populations (refugia). Population dynamics + Contagion + (Mice movement) Mathematical model Single control parameter in the model simulate environmental effects. The other two appear as consequences of a bifurcation of the solutions. BASIC MODEL (no mice movement yet!) dM S MSM = bM − c M S − − aM S M I , dt K dM I MIM = −c M I − + aM S M I , dt K MS (t) : Susceptible mice MI (t) : Infected mice M(t)= MS (t)+MI (t): Total mouse population carrying capacity Rationale behind each term Births: bM → only of susceptibles, all mice contribute to it Deaths: -cMS,I → infection does not affect death rate Competition: -MS,I M/K → population limited by environmental parameter Contagion: ± aMS MI → simple contact between pairs BIFURCATION b Kc = a (b − c ) The carrying capacity controls a bifurcation in the equilibrium value of the infected population. The susceptible population is always positive. The same model, with vertical transmission MS and MI dM S M M = b S M − cM S − S − aM S M I , dt K M M dM I = b I M − cM I − I + aM S M I , dt K 14 a = 0.1 12 c = 0.1 bS = 1.0 10 8 bI = 0.0 6 4 2 0 0 2 4 6 8 10 12 14 K Kc 16 18 20 The same model, with vertical transmission MS and MI dM S M M = b S M − cM S − S − aM S M I , dt K M M dM I = b I M − cM I − I + aM S M I , dt K 14 a = 0.1 12 c = 0.1 bS = 0.99 10 8 bI = 0.01 6 4 2 0 0 Kc = 0 2 4 6 8 10 K 12 14 16 18 20 The same model, with vertical transmission MS and MI dM S M M = b S M − cM S − S − aM S M I , dt K M M dM I = b I M − cM I − I + aM S M I , dt K 14 a = 0.1 12 c = 0.1 bS = 0.9 10 8 bI = 0.1 6 4 2 0 0 Kc = 0 2 4 6 8 10 K 12 14 16 18 20 The same model, with vertical transmission MS and MI dM S M M = b S M − cM S − S − aM S M I , dt K M M dM I = b I M − cM I − I + aM S M I , dt K 14 a = 0.1 12 c = 0.1 bS = 0.8 10 8 bI = 0.2 6 4 2 0 0 Kc = 0 2 4 6 8 10 K 12 14 16 18 20 The same model, with vertical transmission MS and MI dM S M M = b S M − cM S − S − aM S M I , dt K M M dM I = b I M − cM I − I + aM S M I , dt K 14 a = 0.1 12 c = 0.1 bS = 0.5 10 8 bI = 0.5 6 4 2 0 0 Kc = 0 2 4 6 8 10 K 12 14 16 18 20 Temporal behavior K=K(t) A “realistic” time dependent carrying capacity induces the occurrence of extinctions and outbreaks as controlled by the environment. Temporal behavior of real mice critical population Nc=Kc(b-c) Real populations of susceptible and infected deer mice at Zuni site, NM. Nc=2 is the “critical population” derived from approximate fits. THE DIFFUSION PARADIGM ∂u ( x, t ) = r u (1 − u ) + D ∇ 2u ∂t (Fisher, 1937) diffusion nonlinear “reaction” (logistic growth) Epidemics of Hantavirus in P. maniculatus Abramson, Kenkre, Parmenter, Yates (2001-2002) ∂M S ( x, t ) M M = b M − c M S − S − a M S M I + DS ∇ 2 M S , ∂t K ( x) ∂M I ( x, t ) M M = − c M I − I + a M S M I + DI ∇ 2 M I , ∂t K ( x) Three categories of wrongfulness Okubo & Levin, Diffusion and Ecological Problems Wrong but useful: the simplest diffusion models cannot possibly be exactly right for any organism in the real world (because of behavior, environment, etc). But they provide a standardized framework for estimating one of ecology most neglected parameters: the diffusion coefficient. Not necessarily so wrong: diffusion models are approximations of much more complicated mechanisms, the net displacements being often described by Gaussians. Woefully wrong: for animals interacting socially, or navigating according to some external cue, or moving towards a particular place. THE SOURCE OF THE DATA Gerardo Suzán & Erika Marcé, UNM Six months of field work in Panamá (2003) 17 P 27 PA 37 PA 47 B 57 B 67 B 77 B 16 P 26 PA 36 PA 46 B 56 B 66 B 76 B 15 P 25 PA 35 PA 45 B 55 B 65 B 75 B 14 P 24 PA 34 PA 44 B 54 B 64 B 74 B 13 P 23 PA 33 PA 43 B 53 B 63 B 73 B 12 P 22 P 32 P 42 B 52 B 62 B 72 B 11 P 21 P 31 P 41 B 51 B 61 B 71 B Zygodontomys brevicauda Host of Hantavirus Calabazo 60 m THE SOURCE OF THE DATA Terry Yates, Bob Parmenter, Jerry Dragoo and many others, UNM Ten years of field work in New Mexico (1994-) N 100 50 0 -50 -100 Peromyscus maniculatus -100 -50 0 200 m 50 100 Host of Hantavirus Sin Nombre Recapture and age Zygodontomys brevicauda, 846 captures: 411 total mice, 188 captured more than once (2-10 times) P. maniculatus: 3826 captures: 1589 total mice, 849 captured more than once (2-20 times) Recapture probability: J SA A Z. Brevicauda 0.13 0.37* 0.49 P. maniculatus 0.32 0.48 0.58 J: juvenile SA: sub-adult A: adult *One mouse (SA, F) recaptured off-site, 200 m away Different types of movement Adult mice Ö diffusion within a home range Sub-adult mice Ö run away to establish a home range Juvenile mice Ö excursions from nest Males and females… The recaptures 60 Z. brevicauda 40 200 0 -20 P. maniculatus all sites, all mice 100 -40 -60 0 20 40 60 80 Δt (days) 100 120 140 Δx (m) Δx (m) 20 0 -100 -200 0 30 60 90 120 150 180 210 240 270 300 330 time (days) x(t) (m) MOUSE WALKS 60 50 40 30 20 10 0 -10 -20 -30 -40 -50 -60 Z. brevicauda captured ~10 times 926 30 799 835 801 0 P.m. tag 3460 897 898 899 953 927 A117008 A117039 A117075 A117104 A117281 0 30 60 t (days) 90 925 -30 744 745 954 771 -60 834 120 -40 0 40 80 Julian date 2450xxx (Sept. 97 to May 98) An ensemble of displacements An ensemble of displacements An ensemble of displacements …representing the walk of an “ideal mouse” PDF of individual displacements As three ensembles, at three time scales: 0.35 Z. brevicauda 0.30 247 steps 170 steps 17 steps 0.25 dt ~ 1day dt ~ 1 month dt ~ 2 months 0.15 0.10 0.05 0.016 q(x) p(x) (renormalized) Gaussian fit 0.03 0.02 0.00 -0.05 0.012 -80 -60 -40 -20 0 dx 20 40 60 80 p(x) P(dx) 0.20 1 day intervals 0.01 P. maniculatus 0.00 -100 -50 0 50 100 x (m) 0.008 0.004 0.000 -150 -100 -50 0 x (m) 50 100 150 Mean square displacement 3000 2 <Δx >, <Δy > (m ) 600 2500 2 <Δx >, <Δy > (m ) 0 2000 2 200 2 <Δx > 2 <Δy > 2 2 2 400 0 30 60 90 1500 1000 East-West direction North-South direction 500 t (days) 0 Z. brevicauda (Panama) 0 30 60 90 120 150 180 t (days) P. maniculatus (New Mexico) Confinements to diffusive motion • Home ranges • Capture grid L • Combination of bothG G L A harmonic model for home ranges U2 U1 P1(x) -xc/2 U3 P3(x) P2(x) L/2 L/2 L/2 xc1 xc2 xc3 -G/2 G/2 xc/2 ∂P( x, t ) ∂ ⎡ dU ( x ) ⎤ 2 = + ∇ P ( x , t ) D P ( x, t ) ⎢ ⎥ ∂t ∂x ⎣ dx ⎦ PDF of an animal Time dependent MSD 11 L=G 0 0 L=∞ 3000 2500 2 saturation 2000 2 200 2 <Δx > 2 <Δy > 2 2 2 22>/(G22/12) <x <x >/(G /12) 400 <Δx >, <Δy > (m ) L>G 2 <Δx >, <Δy > (m ) 600 30 60 90 1500 1000 L<G 500 t (days) initially diffusive ~t0 Z. brevicauda (Panama) 00 00 0 30 East-West direction box potential, box potential, North-South direction 60 P. maniculatus 2 2 Dt/G Dt/G concentric concentricwith with the 90 window 120 150 180 the window t (days) 0.3 (New0.3 Mexico) Saturation of the MSD 1.0 2 0.8 2 2 <<x >> /(G /6) L /6 0.6 harmonic numerical harmonic analytical box numerical box analytical asymptotics 0.4 from measurements 0.2 0.0 0.0 0.5 1.0 1.5 L/G resulting value 2.0 2.5 Application of the use of the saturation curves to calculate the home range size of P. maniculatus (NM average) Periodic arrangement of home ranges … … 2.0 a a/G 1.5 1.0 2 1.0 0.9 0.8 0.7 0.6 0.5 0.4 0.3 0.2 0.1 0 0.5 0.0 0.0 2 Δx /(G /6) 0.5 1.0 1.5 L/G 2.0 2.5 Periodic arrangement of home ranges 1.0 Measurement 1 (G1 = 1) Measurement 2 (G2 = 0.5) 0.8 Measurement 3 (G3 = 0.75) Δx12 = f (L / G1 , a / G1 ) 0.6 a Δx22 = f (L / G2 , a / G2 ) intersection Δx32 = f (L / G3 , a / G3 ) 0.4 0.2 0.0 0.0 0.2 0.4 0.6 L 0.8 1.0 SUMMARY ¾Simple model of infection in the mouse population ¾Important effects controlled by the environment ¾Extinction and spatial segregation of the infected population ¾Propagation of infection fronts ¾Delay of the infection with respect to the suceptibles ¾Mouse “transport” is more complex than diffusion ¾Different subpopulations with different mechanisms •Existence of home ranges •Existence of “transient” mice ¾Limited data sets can be used to derive some statistically sensible parameters: D, L, a ¾Possibility of analytical models Thank you! TRAVELING WAVES How does infection spread from the refugia? The sum of the equations for MS and MI is Fisher’s equation for the total population: ⎛ M ⎞ ∂M ( x, t ) ⎟⎟ + D∇ 2 M = (b − c) M ⎜⎜1 − ∂t ⎝ (b − c) K ⎠ (Fisher, 1937) There exist solutions of this equations in the form of a front wave traveling at a constant speed. Traveling waves of the complete system Allowed speeds: vS ≥ 2 D(b − c) vI ≥ 2 D[− b + aK (b − c)] Depends on K and a Two regimes of propagation: v I < vS if K < K 0 v I = vS if K > K 0 2b − c K0 = a (b − c ) The delay Δ is also controlled by the carrying capacity 1. Spatio-temporal patterns in the Hantavirus infection, by G. Abramson and V. M. Kenkre, Phys. Rev. E 66, 011912 (2002). 2. Simulations in the mathematical modeling of the spread of the Hantavirus, by M. A. Aguirre, G. Abramson, A. R. Bishop and V. M. Kenkre, Phys. Rev. E 66, 041908 (2002). 3. Traveling waves of infection in the Hantavirus epidemics, by G. Abramson, V. M. Kenkre, T. Yates and B. Parmenter, Bulletin of Mathematical Biology 65, 519 (2003). 4. The criticality of the Hantavirus infected phase at Zuni, G. Abramson (preprint, 2004). 5. The effect of biodiversity on the Hantavirus epizootic, I. D. Peixoto and G. Abramson (preprint, 2004). 6. Diffusion and home range parameters from rodent population measurements in Panama, L. Giuggioli, G. Abramson, V.M. Kenkre, G. Suzán, E. Marcé and T. L. Yates, Bull. of Math. Biol (accepted, 2005). 7. Diffusion and home range parameters for rodents II. Peromyscus maniculatus in New Mexico, G. Abramson, L. Giuggioli, V.M. Kenkre, J.W. Dragoo, R.R. Parmenter, C.A. Parmenter and T.L. Yates (preprint, 2005). 8. Theory of home range estimation from mark-recapture measurements of animal populations, L. Giuggioli, G. Abramson and V.M. Kenkre (preprint, 2005).