Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
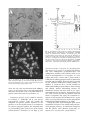
J Mol Evol (2005) 60:257–267 DOI: 10.1007/s00239-004-0185-6 Evolution of Plant-Like Crystalline Storage Polysaccharide in the Protozoan Parasite Toxoplasma gondii Argues for a Red Alga Ancestry Alexandra Coppin,1 Jean-Stéphane Varré,2 Luc Lienard,1 David Dauvillée,1 Yann Guérardel,1 Marie-Odile Soyer-Gobillard,3 Alain Buléon,4 Steven Ball,1 Stanislas Tomavo1 1 Laboratoire de Chimie Biologique, CNRS UMR 8576, Université des Sciences et Technologies de Lille, 59655 Villeneuve dÕAscq cedex, France 2 Laboratoire dÕInformatique Fondamentale de Lille, CNRS UMR 8022, Université des Sciences et Technologies de Lille, 59655 Villeneuve dÕAscq cedex, France 3 Laboratoire Arago, Observatoire océanologique, CNRS UMR 7628, Université Paris VI, 66651 Banyuls-sur-mer cedex, France 4 Institut National de la Recherche Agronomique, Rue de la Géraudiére, 44316, Nantes cedex 03, France Received: 17 June 2004 / Accepted: 9 September 2004 [Reviewing Editor: Dr. Patrick Keeling] Abstract. Single-celled apicomplexan parasites are known to cause major diseases in humans and animals including malaria, toxoplasmosis, and coccidiosis. The presence of apicoplasts with the remnant of a plastid-like DNA argues that these parasites evolved from photosynthetic ancestors possibly related to the dinoflagellates. Toxoplasma gondii displays amylopectin-like polymers within the cytoplasm of the dormant brain cysts. Here we report a detailed structural and comparative analysis of the Toxoplasma gondii, green alga Chlamydomonas reinhardtii, and dinoflagellate Crypthecodinium cohnii storage polysaccharides. We show Toxoplasma gondii amylopectin to be similar to the semicrystalline floridean starch accumulated by red algae. Unlike green plants or algae, the nuclear DNA sequences as well as biochemical and phylogenetic analysis argue that the Toxoplasma gondii amylopectin pathway has evolved from a totally different UDP-glucose-based metabolism similar to that of the floridean starch accumulating red alga Cyanidioschyzon merolae and, to a lesser extent, to those of glycogen storing animals or fungi. In both red algae and apicomplexan parasites, isoamylase and glucan–water dikinase sequences are proposed to explain the appearance of semicrystalline Correspondence to: Stanislas Tomavo; email: Stan.Tomavo@ univlille1.fr starch-like polymers. Our results have built a case for the separate evolution of semicrystalline storage polysaccharides upon acquisition of photosynthesis in eukaryotes. Key words: T. gondii — Plant-like metabolism — Amylopectin — Floridean starch — Evolutionary origin — Glucan water dikinase — Isoamylase — Rhodophyte Introduction The vast majority of living cells store glucose in the form of glycogen. This hydrosoluble polysaccharide consists of a-1, 4-linked glucans that are branched in the a-1, 6 position. Bacteria are known to use ADPglucose as a substrate for glycogen synthesis, while fungi and animal cells use UDP-glucose. The ADPglucose and UDP-glucose pathways differ not only by the nature of the precursor substrate but also by other characteristics. These include distinct regulatory mechanisms and different sets of particular enzymes (Ballicora et al. 2003; Ball and Morell 2003). Green algae, and land plants accumulate starch as an insoluble structurally distinct form of a-1, 4- 258 linked, a-1, 6-branched glucose polymer that is localized within the plastids. It is generally agreed that starch synthesis has evolved from the pathway that existed in the cyanobacterial ancestor of plastids. Unlike glycogen, starch is semicrystalline and organized in a complex granule as a mix of moderately branched amylopectin and mostly linear amylose molecules (Ball and Morell 2003). A wealth of molecular and genetic data conclusively show that starch in land plants and green algae is synthesized through the prokaryote ADP-glucose-based pathway (Recondo and Leloir 1961; Greenberg and Preiss 1964). Green algae and plants both contain numerous and redundant isoenzymes corresponding to each step of the pathway (Ball and Morell 2003; Ral et al. 2005). The significance of this high number of isoenzymes still escapes us and could be related to the formation of crystalline polysaccharides. Other photosynthetic eukaryotes such as the red algae (Viola et al. 2001), the cyanelle-containing glaucophytes, and organisms which are thought to be derived by secondary endosymbiosis of a red algal ancestor (dinoflagellates and cryptophytes) synthesize crystalline polysaccharides outside their plastids. We refer to these as ‘‘floridean starch’’ a name that was coined to describe the form of storage material accumulated in the cytoplasm of red algae. The nature of the pathway used by red algae for storage polysaccharide synthesis is still under debate. Interestingly, dinoflagellates and cryptophytes both harbor plastids which are thought to be derived from the rhodoplasts of red algae (Zhang et al. 1999; Douglas and Penny 1999). Recent molecular evidence has demonstrated that the single-cell apicomplexan parasites harbor a plastid-like nonphotosynthetic apicoplast containing DNA that is somewhat related to the dinoflagellate rhodoplast DNA (McFadden et al. 1996; Wilson et al. 1996; Cavalier-Smith 1999; Marechal and Cesbron-Delauw 2001; Fast et al. 2001). However, it is still controversial whether the apicoplast originated from a green or a red alga (McFadden et al. 1996; Wilson et al. 1996; Cavalier-Smith 1999; Marechal and Cesbron-Delauw 2001; Fast et al. 2001; Köhler et al. 1997; Funes et al. 2002; Waller et al. 2003; Cai et al. 2003). Apicomplexan parasites define important pathogens responsible for widespread diseases such as toxoplasmosis and malaria in humans or coccidiosis in animal species. Some apicomplexan parasites such as Eimeria, the agent of coccidiosis in animals store glucose in the form of amylopectin granules related to floridean starch (Ryley et al. 1969). Others such as Plasmodium, the agent of malaria, lack storage polysaccharides. In humans, Toxoplasma gondii can exist in two distinct forms. At the first stages of infection, the intracellular parasite divides and invades new cells actively. Upon reaction of the host and under pressure of the immune system, this rapidly replicating tachyzoite stage switches into a slowly dividing bradyzoite form. Bradyzoites can be found within dormant cysts in the brain and muscle tissues. These encysted bradyzoites can be reactivated and then differentiated into tachyzoites if the host is immunodepressed. This will lead to fatal toxoplasmic encephalitis, a common death cause in patients with AIDS (Luft and Remington 1988; Luft and Remington 1992). Here we present evidence that the storage polysaccharide accumulated in the cytoplasm by Toxoplasma gondii and that of the dinoflagellate Crypthecodinium cohnii can be defined as a genuine starch. We show that it is synthesized through a UDP-glucose-based pathway much simpler than that described for plants. This pathway is very similar to that we have found to be encoded in the recently sequenced genome of red alga Cyanidioschyzon merolae (Matsuzaki et al. 2004). We propose that two particular functions found in all starch synthesizing organisms (red algae, green algae, plants, apicomplexans) are responsible for the appearance of a semicrystalline insoluble starch granule instead of glycogen. Our data shed light not only on the evolution of apicomplexan parasites but also on those of the green and red lineages of photosynthetic eukaryotes and that of the starch metabolism pathway. Materials and Methods Strains and Growth Conditions. The RH strain of Toxoplasma gondii was grown in HepG2 cells and purified as described (Dzierszinski et al. 1999). However, the culture medium (DulbeccoÕs minimal essential medium containing 10% fetal calf serum) was left until it became acidic (pH 6.2–6.5), conditions known to induce bradyzoite and cyst formation. Media and culture conditions for growing the wild type of Chlamydomonas reinhardtii (137C strain) and the dinoflagellate Crypthecodinium cohnii (ATCC 50050 strain) were previously described (Libessart et al. 1995; Perret et al. 1993). Starch Purification. T. gondii, C. reinhardtii, and C. cohnii cultures were centrifuged and resuspended in 15 ml of phosphatebuffered saline (PBS). Cell suspensions were disrupted three or four times by a French press (10,000 to 15,000 p.s.i.). Crude starch pellets were obtained by spinning down the lysate at 10,000 g for 20 min. The pellets were washed twice with water and passed through a self-formed 90% Percoll gradient. The purified starch pellets were rinsed in distilled water, centrifuged twice at 10,000 g, and kept dry at 4C. Starch amounts were measured by the amyloglucosidase assay (Libessart et al. 1995). Separation of Starch Polysaccharide by Gel Permeation Chromatograghy. Starches (1.5 mg) dissolved in 500 ll of 10 mM NaOH were applied to a Sepharose CL-2B column (0.5 cm · 65 cm) equilibrated in 10 mM NaOH. Fractions of 300 ll were collected at a rate of one fraction each second min. Glucans in each fraction were detected through the iodine–polysaccharide interaction. 259 Chain-Length Distribution Analysis. Purified starches (500 lg) were suspended in 55 mM sodium acetate buffer, pH 3.5, and debranched by 2 units of Pseudomonas amylodermosa isoamylase (Hayashibara Biochemical Laboratory, Japan) at 45C overnight. The samples were further analyzed by high-performance anion-exchange chromatography–pulse amperometric detection (HPAEC-PAD) and capillary electrophoresis. X-ray Diffraction and Nuclear Magnetic Resonance (NMR) Analysis. Powder X-ray diffractograms were collected from purified starches as described previously (Buléon et al. 1997). 1H-NMR spectroscopy of purified starches was performed on a Bruker ASK 400 WB spectrometer. Chemical shifts (expressed as ppm) were measured by reference to internal dimethylsulfoxided6 (d = 2.52 ppm at 70C). The two-dimensional homonuclear correlation spectroscopy (COSY) experiments were performed using the standard Bruker pulse library. Transmission Electron Microscopy. In vitro-induced T. gondii bradyzoites were collected by centrifugation, fixed in 2.5% glutaraldehyde prepared in 0.1 M cacodylate buffer, and post fixed in 1% OsO4 in the same buffer. After ethanol dehydration, the pellet was embedded in Epon. Serial thin sections were cut and collected on longitudinal barred grids. After staining with 2% uranyl acetate prepared in 50% ethanol and incubation with a lead citrate solution, sections were observed on a Hitachi H-600 electron microscope. Transmission electron microscopy (TEM) of Chlamydomonas reinhardtii (137C strain) and Crypthecodinium cohnii were previously described (Libessart et al. 1995; Perret et al. 1993). Gene Identification and Reverse Transcriptase– Polymerase Chain Reaction. The Toxoplasma gondii genes encoding enzymes involved in amylopectin metabolism were identified at the genome Website, http://www.toxodb.org, using key word searches or protein ortholog sequences. All genes were reconstituted in silico using Genewise (Wise 2), Genscan, Genemark. Both exons and introns were determined. The predicted proteins were aligned using ClustalW and the KozakÕs rule was used to determine the translational initiation sites according to the method previously described by Seeber (1997). The following genes were identified: a-amylase (TGG_994355; gene can be found at nucleotide positions 55000 to 60000); isoamylase (TGG_994676; gene at positions 240000 to 255000), D-enzyme (TGG_994353; gene at positions 170000 to 185000), R1 protein (TGG_994467; gene at positions 150000 to 185000), a-glucan phosphorylase (TGG_994281; gene at positions 740000 to 750000), glycogenin (TGG_994281; gene at positions 540000 to 560000), a-glucosidase (TGG_994637; gene at positions,15000–40000), indirect debranching enzyme (TGG_994267; gene at positions 150000 to 175000), UDP-glucose pyrophosphorylase (TGG_994289; gene at positions 535000 to 545000), branching enzyme 1 (TGG_994676; gene at positions 240000 to 250000), branching enzyme 2 (TGG_994574: gene at positions 378000 to 389000), and starch (glycogen) synthase (TGG_994276; gene at positions 60000 to 80000). The transcript level or expression of these genes was demonstrated by RT-PCR. To do this, total RNA from T. gondii tachyzoites and bradyzoites obtained from murine brain cysts was extracted and used for RTPCR as previously described (Dzierszinski et al. 2001). The cDNAs amplified were cloned and sequenced using an Alfexpress DNA automatic sequencer (Amersham Pharmacia Biotech). Enzyme Assays. In vitro-induced bradyzoites of T. gondii, C. reinhardtii, and C. cohnii extracts were prepared using French press disruption of cells suspended in 10 ml of cold 50 mM Hepes buffer, pH 7.0, containing 1 mM MgCl2, 2 mM EDTA, 2 mM bmercaptoethanol, 1 mM PMSF, and a cocktail of protease inhib- itors. After centrifugation at 2000g for 10 min, the supernatants were ultracentrifuged at 100,000 g at 4C for 2 h. The supernatants were recovered and protamine sulfate (20%, w/w) was added. After 10 min on ice, the solutions were centrifuged at 27,000 g at 4C for 30 min. The supernatants were dialyzed against 50 mM Hepes buffer containing 2 mM EDTA at 4C for 24 h and concentrated using Centricon-30 (Amicon). Starch or glycogen synthase activity was determined according to the method described by Karkhanis et al. (1993) using [14C]ADP-glucose (303 mCi/mmol) or [14C]UDPglucose (304 mCi/mmol) purchased from Amersham. Phylogenetic Reconstruction. The phylogenetic analysis was performed using the Phylip package (3.6 version; http:// www.evolution.genetics.washington.edu/phylip.html). Phylogenetic trees were constructed using the neighbor-joining method. Distances were calculated using the Jones–Taylor–Thorton method. Positions corresponding to insertions–deletions were excluded from the analysis. Numbers represent bootstrap replicate values out of 100 (500 bootstrap replicates were computed). The scale (down, left) indicates the branch length corresponding to the number of substitutions per site. Results and Discussion Comparative Analysis of the Structure and Morphology of Crystalline Storage Polysaccharide from T. gondii, C. reinhardtii, and C. cohnii. The presence of abundant floridean starch granules defines a cytological marker of the bradyzoite stage (Fig. 1E) and tachyzoites seem devoid of storage polysaccharides (Tomavo 2001). Like the dinoflagellate Crypthecodinium cohnii (Alveolata) (Fig. 1F), the bradyzoite stage of Toxoplasma gondii (Apicomplexa) accumulates a considerable amount of amylopectin in the cytoplasm .(Fig. 1E), while the unicellular green alga Chlamydomonas reinhardtii (Chlorophyta) contains amylopectin within the chloroplast (Fig. 1D). Toxoplasma gondii, together with Cryptosporidium parvum and the red alga Cyanidionschyzon merolae, defines one of the first floridean starch containing organisms whose genome sequence is presently available (http://www.toxodb.org). We therefore initiated a biochemical characterization of the polysaccharide structure and investigated the pathway of floridean starch biosynthesis and degradation. The polysaccharide was purified from tachyzoites that were induced into bradyzoites in vitro. The purified starch was dispersed in 10 mM NaOH and subjected to size exclusion chromatography in CL2B columns. Results were compared to those obtained with the unicellular green algae Chlamydomonas reinhardtii, which synthesizes starch through the use of the ADPglucose pathway (Fig. 1A). We also compared the results to those gathered from the nonphotosynthetic floridean starch accumulating dinoflagellate Crypthecodinium cohnii (Fig. 1C), a distant cousin of apicomplexan parasites. It is evident from the chromatograms displayed in Fig. 1B that only the high mass amylopectin-like fraction could be recov- 260 Fig. 1. Comparative analysis of the structure and morphology of starches from Toxoplasma gondii, Chlamydomonas reinhardtii, and Crypthecodinium cohnii. CL2B gel permeation chromatography of amylopectin from Toxoplasma gondii (1 mg) dispersed in 10 mM NaOH (B) in comparison with Chlamydomonas reinhardtii (1 mg) (A) and Crypthecodinium cohnii (1 mg) (C). The kmax (maximal absorbance wavelength of the iodine–polysaccharide complex in nanometers) is scaled on the left axis and displayed as a thin solid line in the polysaccharide containing fractions. The optical density (d) of the complex at kmax measured for each fraction is indicated on the right axis. The low kmax fraction excluded from the column defines amylopectin, while the high kmax amylose is separated throughout the column (see arrows). Transmission electron micrographs of nitrogenstarved Chlamydomonas reinhardtii (D), Toxoplasma gondii bradyzoite (E), and a vegetative cell of Crypthecodinium cohnii (F) showing storage polysaccharide or starch (s). ered from Toxoplasma polysaccharide granules, while the low mass amylose fraction recovered from Crypthecodinium cohnii (Fig. 1C) and Chlamydomonas reinhardtii (Fig. 1A) displayed a distribution in size and an iodine interaction identical to those of plant starches. Crystallinity of the Toxoplasma native starch granules was ascertained through the use of wide-angle X-ray diffraction and microscopic observations under nonpolarized and polarized light (Fig. 2). X-Ray powder diffractograms showed a characteristic B-type of diffraction pattern (data not shown), while the granules displayed the classical maltese cross (Fig. 2B) witnessed in many land plant starches. The absence of amylose was confirmed by NMR analysis of the purified polysaccharide (Fig. 3). Indeed, the spectra are consistent with the presence of a moderately branched (1% of a-1, 6 linkages) amylopectin-like component. The purified starches or amylopectin were then subjected to enzymatic debranching and separation of the debranched chains by capillary electrophoresis (Fig. 4). The chain-length histogram distribution displayed in Fig. 4B shows significant differences from those of both Chlamydomonas (Fig. 4A) and Crypthecodinium (Fig. 4C) starches. They remained much more similar to those of amylopectins than to those of glycogen. This feature is shared by many other red algae floridean starches. These patterns of chain-length histogram distribution obtained by capillary electrophoresis were also confirmed by HPAEC-PAD analysis (data not shown). Crude Extract Analysis of the Starch Pathway. Next, we purified as much crude extract as possible from Toxoplasma tachyzoites that were induced to differentiate into bradyzoites in vitro. We monitored both UDP-glucose and ADP-glucose incorporation into rabbit liver glycogen. Results displayed in Fig. 5 show that ADP-glucose is preferred by the Chlamydomonas enzymes. However, significantly more incorporation was found by using UDP-glucose in Toxoplasma gondii and in Crypthecodinium cohnii. The fact that the activity measured in the presence of UDP-glucose with the T. gondii ex- 261 Fig. 3. Deciphering the fine structure of storage polysaccharide by NMR analysis. Superimposed one-dimensional 1H-NMR spectra of Chlamydomonas reinhardtii, Toxoplasma gondii, and Crypthecodinium cohnii starches. A Note that the abundance of peaks seen on right side of the diagram prompted us to represent only the C. cohnii spectrum. The other two samples display exactly the same profile in this area. B The two-dimensional 1H-NMR spectra of C. cohnii starch is shown below. Both T. gondii amylopectin and C. reinhardtii starches display identical NMR spectra. Fig. 2. Crystallinity of the T. gondii amylopectin ascertained using nonpolarized light (A) and polarized light (B) microscopy. The arrows indicate the size (1–3 lm) of one amylopectin granule and the circles display typical maltese crosses. tracts was only twice that measured with ADP-glucose is, in our opinion, due to the high background measured. This in turn is due to the small amount of parasite crude extract that was available to us. Comparative Analysis of the T. gondii, C. merolae, S. cerevisiae, C. reinhardtii and E. coli Storage Polysaccharide Pathway Genes. We probed the completed Toxoplasma genome sequence for the presence of all the enzymes known to be important for starch or glycogen synthesis. We compared these to corresponding genes found in the green lineage (Chlamydomonas reinhardtii), in red algae (Cyani- dioschyzon merolae), in bacteria (E. coli), and in yeast (Saccharomyces cerevisiae). It is evident from Table 1 that both ADP-glucose pyrophosphorylase and ADP-glucose utilizing starch synthase could not be found in apicomplexans or in red algae. It is also evident that both C. merolae and T. gondii contain a UDP-glucose utilizing glycogen (starch) synthase-like sequence and glycogenins. These enzymes are specific for the eukaryote UDP-glucose based pathway. In addition, T. gondii but not C. merolae contains an indirect debranching enzyme similar to those of fungi and animals. Indirect debranching enzymes are bifunctional enzymes that carry both an a-1, 4-glucanotransferase and an amylo-1, 6-glucosidase active site. Table 2 shows that the four distinct conserved blocks of sequences that typify the N-terminal sequences of such enzymes were found both in T. gondii and in Cryptosporidium parvum, another amylopectin producing apicomplexan (Harris et al. 2004). Among the genes which distinguish plant starch metabolism from that of the animal fungal and bacterial glycogen pathways, we found both isoamylase and R1 (glucan water dikinase activity)-like sequences in C. merolae and T. gondii (Table 1). 262 Fig. 4. Comparative polysaccharide chain-length CL distributions. Histogram of CL distributions of Toxoplasma gondii amylopectin (B) in comparison with Chlamydomonas reinhardtii (A) and Crypthecodinium cohnii (C) starches. The CL distributions were obtained after isoamylase-mediated enzymatic debranching through capillary electrophoresis of 8-aminopyrene-1, 3, 6-trisulfonate (APTS)-labeled fluorescent glucans. The X axis displays the degree of polymerization (DPS 7 to 36) and the Y axis represents the relative frequency of chains expressed as percentages. Differential Expression of Starch Biosynthetic and Degradation Genes. The expression pattern of the genes listed in Table 1 was investigated by RT- PCR in tachyzoites and brayzoites isolated from mouse brain cysts. Results displayed in Fig. 6 show that enzymes of amylopectin metabolism are transcribed in tachyzoites and/or bradyzoites. It is noteworthy that transcripts coding for enzymes known to be involved in the catabolic functions such as the R1 protein (GWD), a-glucan phosphorylase, a-glucosidase, and a-amylase seem to be preferentially expressed in bradyzoites. The indirect debranching enzyme transcript, despite being found in tachyzoites, seems to be preferentially expressed in bradyzoites, suggesting a catabolic function. Transcripts coding for enzymes known to be involved in glycogen or starch synthesis are preferentially expressed in tachyzoites (glycogenin, starch (glycogen) synthase, one branching enzyme isoform) but can also be detected in lesser amounts in bradyzoites (Fig. 6). This pattern is consistent with the production of amylopectin during differentiation of tachyzoites into bradyzoites and with the mobilization of the glucose stores during Fig. 5. Substrate requirement (ADP-glucose and/or UDP-glucose) for transglucosylase reaction catalyzed by Chlamydomonas reinhardtii, Toxoplasma gondii and Crypthecodinium cohnii starch (glycogen) synthases. Error bars represent the mean and standard deviations of three reproducible experiments. bradyzoite- to- tachyzoite interconversion (Tomavo 2001). Isoamylase, which has been proposed to be responsible for the crystallization of amylopectin, is found to be transcribed as a biosynthetic gene, consistent with its hypothetical function (Ball and Morell 2003). We believe that the transcription of D-enzyme (a-1, 4-glucanotransferase) reflects a requirement of this function during starch biosynthesis as demonstrated by an analysis of Chlamydomonas mutants (Colleoni et al. 1999). Surprisingly, the preferential transcription of one particular branching enzyme sequence in bradyzoites is also suggestive of some catabolic function for this particular enzyme. A catabolic function remains to be demonstrated for a branching enzyme in plants. However, plant mutants defective for the well-conserved BEI have failed to display a phenotype consistent with a function in starch biosynthesis (Ball and Morell 2003). 263 Table 1. Number of genes corresponding to enzymes of glycogen and starch metabolism UDP-glucose pyrophosphorylase ADP-glucose pyrophosphorylase Starch (glycogen) synthase Branching enzyme Isoamylase Indirect debranching enzyme a-1, 4-Glucanotransferase Phosphorylase a-Amylase a-Glucosidase R1 protein Glycogenin Apicomplexa (T. gondii) Red algae (C. merolae) Green algae (C. reinhardtii) Yeast (S. cerevisiae) Bacteria (E. coli) 1 0 1 2 1 1 1 1 1 1 1 1 1 0 1 1 2 0 2 1 1 1 1 1 1 3 6 3 2 0 2 2 3 0 2 0 3 0 2 1 0 1 0 1 0 3 0 2 1 1 1 1 1 0 1 2 2 1 0 0 Note. The protein-deduced sequences were recovered from Escherichia coli (http://www.genome.wisc.edu/), Saccharomyces cerevisiae (http:// www.yeastgenome.org/) Cyanidiumschyzon merolae (http://merolae.biol.s.u-tokyo.ac.jp/) and Chlamydomonas reinhardtii (http://genome.igipsf.org/chlrel/chlrel.home.html). The Toxoplasma gondii enzymes were identified at the genome Web site (http://www.toxodb.org) by keyword searches or using protein orthologue sequences. Expression of all T. gondii sequences coding for enzymes involved in amylopectin metabolism was checked by RT-PCR (Fig. 6) and nucleotide sequencing. The sequences of Cryptosporidium parvuum, another apicomplexan parasite whose genome has been sequenced, is presently not sufficiently assembled and annotated to allow a systematic identification of full length genes (http://www.cryptodb.org) Table 2. Consensus sequences of indirect debranching enzymes in humans, rabbit, yeast, C. parvum, and T. gondii Consensus sequence Human Rabbit Yeast C. parvum T. gondii Conserved amino acid in the catalytic site N-Terminal domain I N-Terminal domain II N-terminal domain III N-Terminal domain IV C-Terminal peptide C-Terminal peptide 209-NVICITDVVYNH 232-VLCITDVVYNH 218-MLSLTDIVFNH 338-GILSACDVVLNH 371-GLLSTIDLVLNH 521-GVRLDNC 544-GVRLDNC 530-DGFRIDNC 643-HAIRLDNC 671-GVRLDNC * 551-VVAELFT 574-VVAELFT 560-YVVAELFS 673-VYAELF 701-WIFAELFT * 619-ALFMDITHD 642-ALFMDITHD 662-LFMDCTHD 769-AIFFDCTHD 853-LFYDCTHD * 1099-WGRDTFJ 1122-WGRDTFI 1083-WGRDVFI 1426-WGRDTFI 1505-WGRDTEI * 1161-IQDYC 1184-IQDYC 1145-VQDYV 1488-ALDYC 1567-IQDYS * Asp-535 Glu-564 Asp-670 Asp-1086 Asp-1147 Note. The GenBank and other database accession numbers of indirect debranching enzymes used are as follows: human (P35573), rabbit (P35574), yeast (Q06625), C. parvum (Q7YY51) and T. gondii (contig TGG_994267 from http://www.toxodb.org). The amino acids were numbered from the N-terminal end. Conserved residues are in bold face. Asterisks indicate the amino acid residues that are essential for the catalytic sites. Phylogenetic Analyses. Phylogenetic analyses performed with two key enzymes (amylopectin synthase and the R1 protein) demonstrate that the Toxoplasma gondii amylopectin synthesis pathway has evolved from the red algal starch synthesizing machinery through a secondary endosymbiotic event (Figs. 7 and 8). We believe that these phylogenetic data, together with our enzyme assays (Fig. 5), establish that apicomplexans (Toxoplasma gondii and Cryptosporidium parvum) and red algae, such as C. merolae, use a UDP-glucose pathway to build insoluble crystalline amylopectin. That this is the pathway used by all floridean starch accumulating organisms is suggested by the recent characterization of a UDP-glucose utilizing amylopectin synthase in red algae (Nyvall et al. 1999). However, it is equally apparent that Toxoplasma and C. merolae also contain plant-like genes that are not found in yeast and mammals (Table 1). These consist of genes that are required in plants and green algae to breakdown and synthesize semicrystalline polysaccharides. For instance, one of the early steps of starch mobilization in plants requires the recently identified R1 protein. This enzyme has also been named a-glucan water dikinase (GWD). It has been shown to phosphorylate amylopectin at specific positions and is thought to facilitate subsequent hydrolytic attack through amylases (Ritte et al. 2002). In addition, both bacteria and plants polysaccharide catabolism leads to the production of malto-oligosaccharides which are not generated from glycogen in yeasts or mammalian cells. Toxoplasma gondii and C. merolae also contain, 264 Fig. 6. Results of semiquantitative RT-PCR of expression of genes coding for enzymes involved in T. gondii amylopectin metabolism. Note the bradyzoite stage- specific expression of aamylase, a-glucosidase, R1 protein, and a-glucan phosphorylase, while the other genes are expressed in both tachyzoite and bradyzoite. Except for glycogenin, the other genes are overexpresed either in tachyzoites (isoamylase, D-enzyme, branching enzyme 2, and starch [glycogen] synthase) or in bradyzoites (branching enzyme 1, UDP-glucose pyrophosphorylase, and debranching [indirect] enzyme). respectively, one or two genes encoding enzymes of malto-oligosaccharide metabolism such as an a-1, 4glucanotransferase (D-enzyme). This hints that the evolution of semicrystalline polysaccharides has required the maintenance or acquisition of genes similar to those that are necessary in green algae and plants to mobilize glucose from such polymers (Table 1). The presence of an isoamylase-related expressed sequence in the genome also hints that maintenance of this activity is required to ensure the synthesis of an insoluble semicrystalline form of storage polysaccharide. Indeed, this enzyme has been proposed to ensure the processing of the hydrosoluble precursor of amylopectin into mature crystalline material (Ball and Morell 2003). Apicomplexans and dinoflagellates are suspected to have evolved following a secondary endosymbiotic event. It is likely that the ancestral host cell engulfed a Fig. 7. Phylogenetic tree of starch, amylopectin, and glycogen synthases. Unrooted phylogenetic tree from starch (glycogen) synthases using neighbor joining (NJ). Sequences of starch (glycogen) synthases were aligned with ClustalW (Thompson et al. 1994). Boostrap values are indicated. The scale indicates the branch length corresponding to the number of substitutions per site. The GenBank and other database accession numbers of the starch (glycogen) synthases used for the tree analysis are as follows: H. sapiens (P13807), M. musculus (Q9Z1E4), O. cuniculus (P13834), S. cerevisiae 1 (P23337), S. cerevisiae 2 (P27471), N. crassa (O93869), A. thumefaciens (P39670), S. melilati (P58593), B. halodurans (Q9KDX6), B. subtilis (P39125), C. muridarum (Q9PLC3), C. pneumoniae (Q9Z6V8), C. tepidum (Q8KAY6), C. trachomatis (084804), C. perfringens (Q8XPA1), D. radiodurans (Q9RWS1), E. coli (P08323), F. nucleatum (Q8RF65), H. influenzae (P45176), L. lactis (Q9CHM9), R. loti (Q985P2), R. tropici (Q9EUT5), S. penumoniae (Q97QS5), T. caldophilus (P58395), T. maritima (Q9WZZ7), V. cholerae (Q9KRB6), Y. pestis (Q8ZA78), C. merolae (locus CMM317C from http://merolae.biol.s.u-tokyo.ac.jp/), T. gondii (contig TGG_994276 from http://toxodb.org), C. reinhardtii (AAL28128), S. acidocaldarius (Q9hh97), S. solfataricus (Q97ZD3), P. abyssi (Q9V2J8), P. furisus (Q8TZF1), P. aerophilum (Q8ZT56), A. majus (O82627), A. thaliana (Q9MAQ0), Z. mays GB (P04713), O. sativa (P19395), P. sativum (Q43092), S. tuberosum (Q00775), S. vulgare (Q43134), T. aestivum (P27736), A. thaliana (Q9FNF2), O. sativa (Q40739), and S. tuberosum (P93568). red alga that contained the UDP-glucose based pathway of floridean starch metabolism. This host cell, like most nonphotosynthetic eukaryotes, probably contained its own UDP-glucose-based pathway of glycogen synthesis. There is a possibility that additional genes of the ancestral host were maintained and thus added to the pathway of floridean starch metabolism after secondary endosymbiosis. The absence and presence of indirect debranching enzyme sequences, respectively, in red algae and apicomplexans could be understood in this light. However, we cannot presently exclude other mechanisms to explain these differences, such as accidental lateral transfers during evolution. Evidence for such transfers can be found in the Arabidopsis genome, where one of the two branching enzymes is clearly more related to fungal enzymes than to other plant or bacterial sequences. A similar fate seems to explain the presence of one of the two branching enzymes in the phylogeny displayed in Fig. 9. Indeed, the T. gondii BE1 sequence is more similar to those of bacteria than to those of green or red algae. However, the second BE isoform (T. gondii BE2) seems to have 265 Fig. 8. Phylogenetic tree of dikinases including GWD (a-glucan, water dikinase [also known as R1 protein], PPDK (pyruvate, phosphate dikinase), and PPS (pyruvate, water dikinase) family. Unrooted phylogenetic tree from dikinases using neighbor joining (NJ). Sequences were aligned with ClustalW (Thompson et al. 1994). Boostrap values are indicated. The scale indicates the branch length corresponding to the number of substitutions per site. The GenBank and other database accession numbers of the dikinases used for the tree analysis are as follows: T. gondii (contig TGG_994467 from http://toxodb.org), C. merolae (locus CMT547C from http://merolae.biol.s.u-tokyo.ac.jp/), C. parvum (EAK88766), A. thaliana 1 (AAG47821), A. thaliana 2 (AAO42141), C. reticulata (AAM18228), S. tuberosum (T07050), T. aestivum (CAC22583), C. symbosium (P22983), F. bidentis (S56650), O. sativa (BAA22420), E. histolytica (AAA 18944), Z. mays (P1 1155), E. coli (S20554), M. maripaludis (AAD28736), N. meningitidis (NP_273662), P. aeruginosa (AAG05159), and S. marinus (S51006). evolved separately from other BE of fungi, animals, plants, and bacteria. Once again, this could be related to genes that have been retained from the host cell that engulfed a red alga. Despite the presence of these additional plant-like genes, both Toxoplasma and C. merolae define remarkably simple systems leading to the biosynthesis of semicrystalline amylopectin polymers. Indeed, a recent study establishes that multiple forms of enzymes catalysing each synthetic or degradation step is an ancient characteristic of the green lineage (Ral et al. 2005). It is indeed remarkable that Ostreoccus tauri, a unicellular green alga, requires, for example, six distinct elongation enzymes to build a similar structure and that these isoenzymes have been conserved throughout the evolution of plants. The results reported in this paper, together with our knowledge of storage polysaccharide in green algae, suggest that there has been a strong drive to maintain storage polysaccharides into an insoluble semicrystalline physical state. This has occurred in several independent lineages using very different enzymatic toolkits. In agreement with an apicoplast Fig. 9. Phylogenetic tree of starch (glycogen) branching enzymes (BE). Unrooted phylogenetic tree from starch (glycogen) branching enzymes using neighbor joining (NJ). Sequences were aligned with ClustalW (Thompson et al. 1994). Boostrap values are indicated. The scale indicates the branch length corresponding to the number of substitutions per site. The GenBank and other database accession numbers of the starch (glycogen) branching enzymes used for the tree analysis are as follows: B. subtilis (P39118), E. coli (P07762), H. sapiens (Q04446), S. tuberosum (P30924), S. cerevisiae (P32775), T. gondii BE1 (contig TGG_994676 from http:// www.toxodb.org), G. gracilis (AAB97471), S. pneumoniae (Q97QS8), C. parvum (CAD98370), C. merolae (locus CMH144C from http://merolae.biol.s.u-tokyo.ac.jp/), E. chrysanthemi (Q8GQC5), M. tuberculosis (Q10625), O. sativa (Q01401), M. musculus (NP_083079), F. catus (AAR13899), T. gondii BE2 (contig TGG_994574 from http://www.toxodb.org), A. thaliana (NP_1 95985), Z. mays (Q08047), X. axonopodis (Q8PR13), B. japonicum (Q89FD3), R. solanasearum (Q8XT76), and Y. pestis (Q8ZA75). of red algal evolutionary origin (Wilson et al. 1996; Fast et al. 2001), apicomplexan parasites have inherited the pathway of floridean starch metabolism. This pathway involves a mosaic of genes of prokaryotic and eukaryotic origin. The green algae and land plants display a pathway relying entirely on genes related to the bacterial glycogen metabolism. Because plastids are presently thought to be monophyletic, it is reasonable to assume that the common ancestors of the green and red lineages displayed both eukaryotic and prokaryotic pathways. Subsequently, when the ancestors of green and red algae diverged they maintained either the endosymbiont or the host pathways of glycogen accumulation. This occurred with a concomitant loss of most of the host or endosymbiont redundant functions. It is striking that the cellular localization of storage polysaccharides in either the endosymbiont or the host cytoplasm correlates with the pathway chosen. Despite the loss of photosynthesis in apicomplexans, the presence of this inherited pathway remained suitable and probably required for those parasites that harbor classical 266 dormant cysts in their life cycles. However, for those apicomplexans such as Plasmodium species, which propagate without dormant encysted forms, storage polysaccharide metabolism was lost. Indeed, screening the full genome database of Plasmodium falciparum (http://www.plasmoDB.org) failed to yield any sequence involved in glycogen or starch metabolism. Interestingly in bacteria, the loss of glycogen metabolism has been correlated with the more parasitic and dependent life cycles (Henrissat et al. 2002). Concluding Remarks. Our findings indicate that the storage polysaccharide accumulated in the cytoplasm of apicomplexans such as Toxoplasma gondii and Cryptosporidium parvum and that of the dinoflagellate Crypthecodinium or of the red alga C. merolae are synthesized using a UDP-glucosebased metabolic pathway. This metabolic pathway is very similar to the fungal and animal glycogen pathway and distinct from the plant starch pathway. Thus, the complexity that has been conserved throughout the evolution of green plants is not per se required to build semicrystalline polymers in apicomplexan parasites. It is also striking that the only genes that distinguish the parasiteÕs pathway from that of fungi and animal cells are precisely those that are thought to have been recruited from the bacterial ancestor of plastids by green plants to achieve the biosynthesis of semicrystalline polysaccharides. These data shed light not only on the evolution of the green and red lineages of photosynthetic eukaryotes but also on the basic distinction between hydrosoluble glycogen and semicrystalline starch metabolism. Considerable progress has been made in understanding the biogenesis of glycogen in mammals and that of starch in plants. The different sets of enzymes involved have been identified and characterized. Developing a better understanding of amylopectin biogenesis in Toxoplasma gondii is worthy of further exploration since compounds which inhibit this process could define potential drugs for combating apicomplexan-mediated diseases. Acknowledgments. For comments, logistical support, and invaluable technical assistance in the field and lab, we thank Michael Kibe, Marléne Mortuaire, Hervé Moreau, Christian Slomianny, Emmanuel Maes, Frédéric Chirat, Yves Leroy, Florence Dzierszinski, Brigitte Bouchet, and Bruno Pontoire. We acknowledge the Toxoplasma Genome Sequencing Consortium for making available the genome database: Preliminary genomic and/or cDNA sequence data were accessed via http://ToxoDB.org and/or http:// www.tigr.org/tdb/t_gondii/. Genomic data were provided by The Institute for Genomic Research (supported by NIH Grant AI05093), and by the Sanger Center (Wellcome Trust). EST sequences were generated by Washington University (NIH Grant 1R01AI045806-01A1). This research was funded by the Centre National de la Recherche Scientifique (CNRS) through the Action Thématique Incitative sur Programme et Equipe (ATIPE), the Programme Inter-organisme de Microbiologie Fondamentale, and the Agence Nationale de la Recherche sur le Sida (ANRS). References Ball SG, Morell MK (2003) From bacterial glycogen to starch: understanding the biogenesis of the plant starch granule. Annu Rev Plant Biol 54:207–233 Ballicora MA, Iglesias AA, Preiss J (2003) ADP-glucose pyrophosphorylase, a regulatory enzyme for bacterial glycogen synthesis. Microbiol Mol Biol Rev 67:213–225 Buléon A, Gallant D-J, Bouchet B, Mouille G, DÕHulst C, Kossman J, Ball SG (1997) Starches from A to C. Chlamydomonas reinhardtii as a model microbial system to investigate the biosynthesis of the plant amylopectin crystal. Plant Physiol 115:949–957 Cai X, Fuller AL, McDougald LR, Zhu G (2003) Apicoplast genome of the coccidian Eimeria tenella. Gene 321:39–46 Cavalier-Smith T (1999) Principles of protein targeting in secondary symbiogenesis: euglenoid, dinoflagellate and sporozoan plastid origins and the eukaryote family tree. J Euk Microbiol 46:347–366 Colleoni C, Dauvillee D, Mouille G, Buleon A, Gallant D, Bouchet B, Morell M, Samuel M, Delrue B, dÕHulst C, Bliard C, Nuzillard JM, Ball S (1999) Genetic and biochemical evidence for the involvement of alpha-1, 4-glucanotransferases in amylopectin synthesis. Plant Physiol 120:993–1004 Douglas SE, Penny SL (1999) The plastid genome of the cryptophyte alga, Guillardia theta: complete sequence and conserved synteny groups confirm its common ancestry with red algae. J Mol Evol 48:236–244 Dzierszinski F, Popescu O, Toursel C, Slomianny C, Yahiaoui B, Tomavo S (1999) The protozoan parasite Toxoplasma gondii expresses two functional plant-like glycolytic enzymes. Implications for evolutionary origin of apicomplexans. J Biol Chem 274:24888–24895 Dzierszinski F, Mortuaire M, Dendouga N, Popescu O, Tomavo S (2001) Differential expression of two plant-like enolases with distinct enzymatic and antigenic properties during stage conversion of the protozoan parasite Toxoplasma gondii. J Mol Biol 309:1017–1027 Fast NM, Kissinger JC, Roos DS, Keeling PJ (2001) Nuclear-encoded, plastid-targeted genes suggest a single common origin for apicomplexan and dinoflagellate plastids. Mol Biol Evol 18:418–426 Funes S, Davidson E, Reyes-Prieto A, Magallon S, Herion P, King MP, Gonzalez-Halphen D (2002) A green algal apicoplast ancestor. Science 298:2155 Greenberg E, Preiss J (1964) The occurrence of adenosine diphosphate glucose: glycogen transglucosylase in bacteria. J Biol Chem 239:4314–4315 Harris JR, Adrian M, Petry F (2004) Amylopectin: a major component of the residual body in Cryptosporidium parvum oocysts. Parasitology 128:269–282 Henrissat B, Deleury E, Coutinho PM (2002) Glycogen metabolism loss: a common marker of parasitic behaviour in bacteria?. Trends Genet 18:437–440 Karkhanis YD, Allocco JJ, Schmatz DM (1993) Amylopectin synthase of Eimeria tenella: identification and kinetic characterization. J Euk Microbiol 40:594–598 Köhler S, Delwiche CF, Denny PW, Tilney LG, Webster P, Wilson RJ, Palmer JD, Roos DS (1997) A plastid of probable green algal origin in apicomplexan parasites. Science 275:1485–1489 Libessart N, Maddelein ML, Van den Koornhuyse N , Decq A, Delrue B, Mouille G, DÕHulst C, Ball S (1995) Storage, photosynthesis, and growth: the conditional nature of mutations affecting starch synthesis and structure in Chlamydomonas. Plant Cell 7:1117–1127 Luft BJ, Remington JS (1988) AIDS commentary. Toxoplasmic encephalitis. J Infect Dis 157:1–6 267 Luft BJ, Remington JS (1992) Toxoplasmic encephalitis in AIDS. Clin Infect Dis 15:211–222 Marechal E, Cesbron-Delauw MF (2001) The apicoplast: a new member of the plastid family. Trends Plant Sci 6:200–205 Matsuzaki M, Misumi O, Shin IT, Maruyama S et al. (2004) Genome sequence of the ultrasmall unicellular red alga Cyanidioschyzon merolae 10D. Nature 428:653–657 McFadden GI, Reith ME, Munholland J, Lang-Unnasch N (1996) Plastid in human parasites. Nature 381:482 Nyvall P, Pelloux J, Davies HV, Pedersen M, Viola R (1999) Purification and characterisation of a novel starch synthase selective for uridine 5¢-diphosphate glucose from the red alga Gracilaria tenuistipitata. Planta 209:143–152 Perret E, Davoust J, Albert M, Besseau L, Soyer-Gobillard M-O (1993) Microtubule organization during the cell cycle of the primitive eukaryote dinoflagellate Crypthecodinium cohnii. J Cell Sci 104:639–651 Ral JP, Derelle E, Ferraz C, Wattebled F, Farinas B, Corellou F, Buléon A, Slomianny MC, Delvalle D, dÕHulst C, Rombauts S, Moreau H, Ball S (2004) Starch division and partitioning a mechanism for granule propagation and maintenance in the picophytoplanktonic green alga Ostreococcus tauri. Plant Physiol 36:3333–3340 Recondo E, Leloir L (1961) Adenosine diphosphate glucose and starch biosynthesis. Biochem Biophys Res Commun 6:85– 88 Ritte G, Lloyd JR, Eckermann N, Rottmann A, Kossmann J, Steup M (2002) The starch-related R1 protein is an alpha-glucan, water dikinase. Proc Natl Acad Sci USA 99:7166–7171 Ryley JF, Bentley M, Manners DJ, Stark JR (1969) Amylopectin, the storage polysaccharide of the coccidian Eimeria brunetti and E. tenella. J Parasitol 55:839–845 Seeber F (1997) Consensus sequence of translational initiation sites from Toxoplasma gondii genes. Parasitol Res 83:309–311 Thompson JD, Higgins DG, Gibson TJ (1994) ClustalW: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22:4673–4680 Tomavo S (2001) The differential expression of multiple isoenzyme forms during stage conversion of Toxoplasma gondii: an adaptive developmental strategy. Int J Parasitol 31:1023–1031 Viola R, Nyvall P, Pedersen M (2001) The unique features of starch metabolism in red algae. Proc R Soc Lond B Biol Sci 268:1417– 1422 Waller RF, Keeling PJ, Van Dooren GG , McFadden GI (2003) Comment on a green algal apicoplast ancestor. Science 301:49 Wilson RJM, Denny PW, Preiser PR, Rangachari K, Roberts K, Roy A, Whyte A, Strath M, Moore DJ, Moore PW, Williamson DH (1996) Complete gene map of the plastid-like DNA of the malaria parasite Plasmodium falciparum. J Mol Biol 261:155–172 Zhang Z, Green BR, Cavalier-Smith T (1999) Single gene circles in dinoflagellate chloroplast genomes. Nature 400:155–159